福建黄兔线粒体基因组全序列测定与分析
2020-09-10鲍志远赵博昊胡帅帅吴信生
周 彤,周 娟,梁 爽,鲍志远,赵博昊,胡帅帅,陈 阳,吴信生
(扬州大学 动物科学与技术学院,江苏扬州 225009)
绝大多数动物线粒体DNA(Mitochondrial DNA,mtDNA)为闭合的双链环,长度为16~17.5kb,基因组排列紧凑,每个细胞含有成千上万个线粒体基因组 DNA 拷贝,结构基本一致[1]。线粒体DNA与核基因相比具有结构简单保守、母系遗传、无组织特异性、多拷贝和进化速度快的特征,在研究亲缘关系和物种进化等方面具有重要价值[2]。探究动物线粒体DNA的结构特征,可以为鉴定物种的起源与进化提供非常重要的资料与参考。童晓梅等[3]分析藏鸡mtDNA部分序列,发现藏鸡与家鸡都起源于红原鸡,并归属于红原鸡种;汪琦等[4]通过构建牛亚科代表性品种mtDNAD-loop序列的系统进化树,揭示了三江黄牛与秦川牛、平武牛亲缘关系较近;王洪亮等[5]基于线粒体DNA4个基因序列对矮小梅花鹿群体进行起源进化分析,认为该矮小群体有3个母系起源。福建黄兔为原产于福建的家兔品种,因毛色独特、耐粗饲、适应性强和药用价值高而素有“药膳兔”之称,是目前中国保存与开发利用最好、种群最大的地方品种[6]。福建黄兔肉品质优良、药用价值高,因此研究主要集中在生长发育与肉质特性等方面[7-10],而针对福建黄兔起源和遗传特性的研究相对较少,俞春英等[11]利用15个微卫星位点分析了福建黄兔群体的遗传多态性;赵纪萍[12]基于6个地方家兔品种SNPs信息对福建黄兔的遗传地位进行了初步分析。因此,本研究对福建黄兔的mtDNA进行全序列测定与结构分析,以期为福建黄兔的物种分类、种质鉴定和资源保护提供可靠资料。
1 材料与方法
1.1 试验动物
江苏省金陵种兔场提供6只健康成年福建黄兔,耳中央动脉采血,EDTA试剂抗凝,2~10 ℃条件下带回实验室,于-80 ℃超低温冰箱保存,待用。
1.2 基因组DNA提取
利用TIANamp Genomic DNA Kit试剂盒(TIANGEN,北京)提取基因组DNA,使用Nanodrop 1000测定样品的质量浓度和纯度,选留高纯样品。
1.3 引物设计与PCR扩增
根据GenBank公布的近缘物种穴兔的mtDNA全序列(GenBankNo. AJ001588.1),利用Premier 6.0设计12对覆盖福建黄兔线粒体基因组全部序列的特异性引物,引物信息参表1。由擎科生物技术(南京)有限公司进行引物合成,PCR扩增体系(50 μL):2×Phanta Max Buffer 25 μL,dNTP Mix(10 mmol/L each) 1 μL,上、下游引物(10 μmol/L)各2 μL,模板DNA(50~400 ng/μL)1 μL,Phanta Max Super-Fidelity DNA Ploymerase 1 μL,ddH2O 18 μL。引物P1、P2、P3、P4、P5、P6、P8、P9、P10、P12的PCR扩增程序:95 ℃预变性3 min;95 ℃变性15 s,63 ℃复性30 s,延伸3 min,34个循环;72 ℃延伸5 min。引物P7和P11的PCR扩增程序:95 ℃预变性 3 min;95 ℃变性15 s,59 ℃复性15 s,72 ℃延伸 3 min,34个循环;72 ℃延伸5 min。PCR产物送华大基因公司测序。高保真DNA聚合酶等PCR扩增试剂购于诺唯赞生物科技(南京)有限公司,胶回收试剂盒购于宝日医生物技术(北京)有限 公司。

表1 福建黄兔线粒体基因组序列扩增引物信息Table 1 Mitochondrial genome primer information of Fujian Yellow Rabbit
1.4 序列拼接、注释与分析
利用SeqMan 7.0软件对测序结果进行比对、拼接和校正,得到福建黄兔线粒体基因组全序列。利用Clustal X、tRNA scan-SE和MegAlign等软件对基因组序列的结构特点进行分析,包括各基因的位置与大小、碱基组成与偏倚度、tRNA二级结构、氨基酸偏好性等。使用OGDRAW在线软件进行福建黄兔线粒体基因组图谱的绘制。用MEGA 7.0软件将其他序列与福建黄兔基 因组相应序列进行多序列比对后构建系统进 行树。
2 结果与分析
2.1 线粒体基因组的结构特征
经比对拼接,福建黄兔线粒体基因组全序列长度为17 000 bp,包括tRNA基因22个、蛋白编码基因13个、rRNA基因2个和控制区1个,基因排列顺序同其他兔种基本一致(图1,表2),将序列提交至GenBank(No.MN518689)。蛋白编码基因(长度为11 391 bp)、tRNA基因(长度为 1 505 bp)、rRNA基因(长度为2 536 bp)和控制区(长度为1 555 bp)分别占整个线粒体长度的 67.00%、8.85%、14.92%和9.15%。37个编码基因中的9个基因[tRNA-Glu、tRNA-Gln、tRNA-Cys、tRNA-Asn、tRNA-Tyr、tRNA-Ala、tRNA-Ser(UCN)、tRNA-Pro]由轻链(L链)编码,其余28个编码基因由重链(H链)编码。其碱基含量为:A(31.41%)>T(28.18%)>C (26.70%)>G(13.71%),A+T含量 (59.59%)高于C+G含量(40.41%)。基因间无内含子,基因排列紧密,全序列共有12处间隔区域和5处重叠区域,最大间隔区域(8 bp)位于tRNA-Gln和tRNA-Met之间,最大重叠区域 (43 bp)位于ATPase8和ATPase6之间。

图1 福建黄兔线粒体基因组序列结构图谱Fig.1 Sequence map of mitochondrial genome of Fujian Yellow Rabbit

表2 福建黄兔线粒体基因组结构Table 2 Mitochondrial genome structure of Fujian Yellow Rabbit
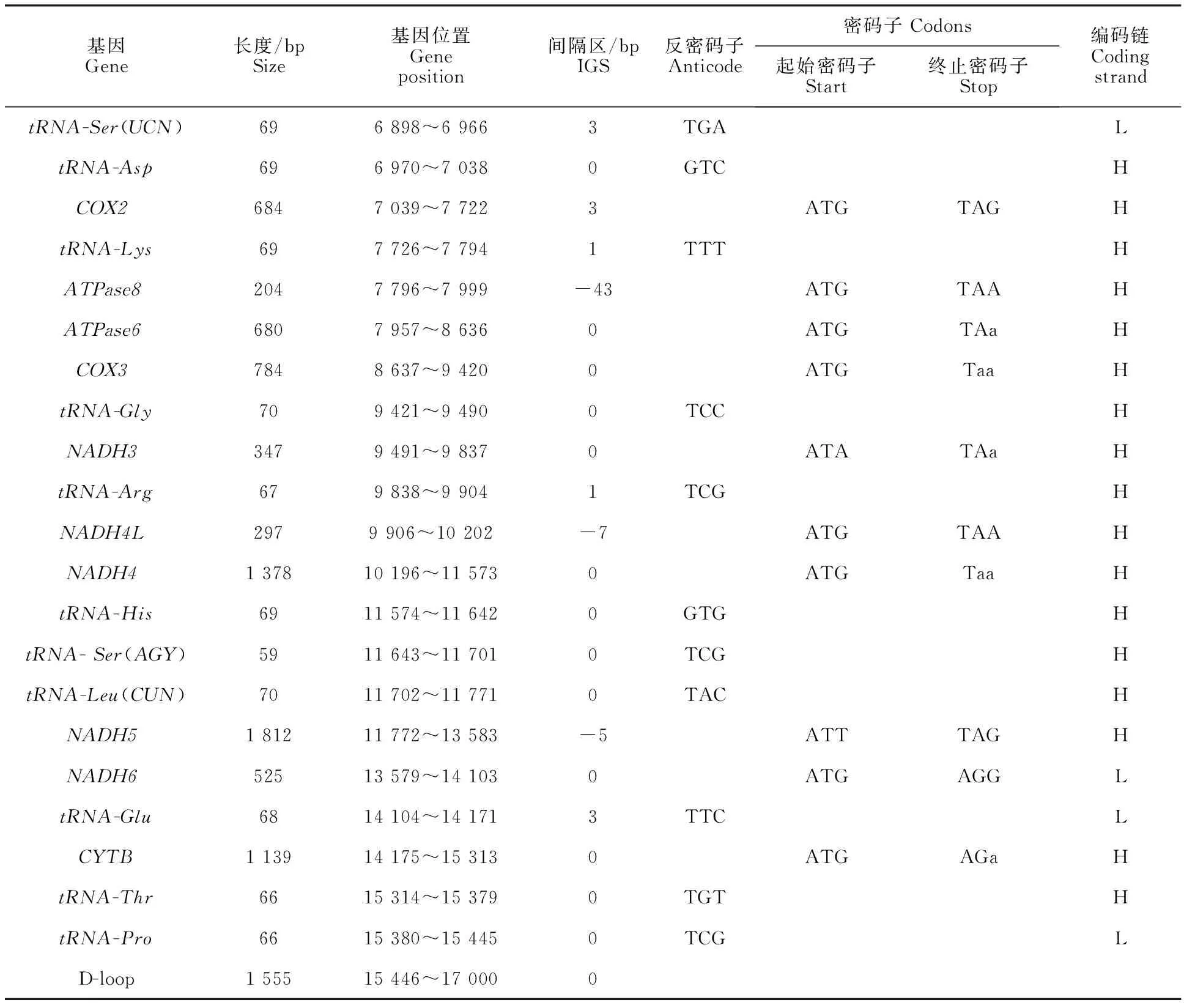
(续表2 Continued table 2)
2.2 蛋白编码基因
福建黄兔线粒体基因组的蛋白编码基因有13个,包括1个CYTB基因、2个ATPase(ATPase8和ATPase6)、3个氧化酶亚基(COX1、COX2和COX3)和7个脱氢酶亚基(NADH1、NADH1-2、NADH1-3、NADH1-4L、NADH1-4、NADH1-5和NADH1-6),使用了3种起始密码子和4 种终止密码子,NADH2和DNAH5使用ATT起始密码子,NADH3使用ATA起始密码子,其余10个蛋白编码基因均使用ATG作为起始密码子,终止密码子以TAA(8个)为主,其次为TAG、AGG和AGA 3种终止密码子。
13个蛋白编码基因除终止密码子外共编码 3 787个密码子,使用频率最高的密码子为UUU(Phe)、CUA(Leu)、AUU(Ile)和AUA(Met)。利用MEGA 7.0软件分析密码子使用情况(表3),发现福建黄兔线粒体编码基因中有CUA、UCA、UGA、CGA等34个偏好密码子(RSCU≥1);密码子第1 位碱基偏倚度最小,T1s、C1s、A1s、G1s( A、T、G、C含量在密码子第3 位碱基总量中所占比)分别为22.40%、24.70%、 31.10%、21.90%;T3s、C3s、A3s、G3s分别为 26.00%、29.90%、37.50%、3.50%,A3s大于T3s、C3s、G3s,表明福建黄兔蛋白编码基因偏好使用以碱基A结尾的密码子。
2.3 基因组tRNA和rRNA
福建黄兔线粒体基因组的14个tRNA位于H链上,8个tRNA位于L链,共22个tRNA。其中tRNA-Leu和tRNA-Ser分别拥有两种反密码子。对tRNA二级结构进行分析发现,21种tRNA均可折叠形成三叶草结构,仅tRNA-Ser(AGY)因为DHU臂缺失而不能正确进行二级结构的折叠,在对应位置上形成一个环状结构,这与已公布的其他兔形目动物tRNA-Ser(AGY)的结构一致。其中,12S rRNA和16S rRNA两基因间相距66 bp,被tRNA-Val隔开,序列长度分别为957 bp、1 579 bp,A、T、A+T含量分别为 35.84%、23.93%、59.77%和36.35%、25.49%、 61.81%,A+T偏度分别为0.199 3和0.176 2。

表3 福建黄兔线粒体基因组的密码子使用频率Table 3 Codon usage frequency of mitochondrial genome in Fujian Yellow Rabbit
2.4 L-链复制起始区和控制区
福建黄兔线粒体基因组中包括L链复制起始区(WANCY区)和控制区(D-loop区)两段非编码区域。L链复制起始区位于WANCY(tRNA-Trp、tRNA-Ala、tRNA-Asn、tRNA-Cys和tRNA-Tyr)基因簇之间,长度为35 bp,该段序列碱基的保守性很强,折叠后形成稳定的茎-环 结构。
福建黄兔控制区长度为1 555 bp,在tRNA-Pro和tRNA-Phe之间,含有较多的重复序列,是基因组中长度变化最大的部分。控制区包含3 个结构域(图2):延长终止序列区(ETAS)、中央保守区(CCSB)和保守序列区(CSB),分别位于1~383 bp、384~701 bp和702~1 555 bp。延长终止序列区中包含两个参与重链复制与终止的序列(ETAS1和ETAS2)。比对多个物种的控制区结构信息,识别出3 段保守序列(CSB1~3)和2 组串联重复序列(短重复序列RS3和长重复序列RS2)。RS3在CSB1和CSB2之间,包含10个拷贝的20 bp长的序列(short repeat, SR);RS2在CSB3 和tRNA-Phe之间,包含2个拷贝的153 bp长的序列(long repeat, LR)。SR表现出重复cbabc基序的内部结构, a=ACCC,b=GTAC, c=GCAC,此外,紧邻RS3序列处存在88 bp SR结构的重排序列。
2.5 系统进化分析
将福建黄兔线粒体基因组D-loop区与韩国野兔(Lepuscoreanus)、雪兔(Lepustimidus)、格拉纳达野兔(Lepusgranatensis)、草兔(Lepuscapensis)、华南兔(Lepussinensis)、美洲兔(Lepusamericanus)、海南兔(Lepushainanus)、高原鼠兔(Ochotonacurzoniae)和柯氏鼠兔(Ochotonakoslowi)9种兔形目动物线粒体D-loop区进行多重序列比对与分析,以柯氏鼠兔和高原鼠兔两种兔形目鼠兔科动物为外群,利用MEGA 7.0软件的邻接法构建系统进化树,分析福建黄兔的进化关系与分类地位(图3)。发现该进化树由两大支构成,鼠兔科的柯氏鼠兔和高原鼠兔聚为一支,韩国野兔、雪兔、海南兔等7种旷兔属物种与福建黄兔(穴兔属)聚为一支,支持将福建黄兔划归为穴兔属。

图2 福建黄兔线粒体基因组控制区结构Fig.2 Control region structure of mitochondrial genome in Fujian Yellow Rabbit

图3 基于线粒体基因组D-loop区采用邻接(NJ)法构建的系统进化树Fig.3 NJ phylogenetic trees based on mitochondrial genome D-loop region
3 讨论与结论
脊椎动物线粒体基因组长度基本稳定在 16~17.5 kb,少数物种序列长度差异较大,如短刀侧殖文昌鱼线粒体基因组长度为14 991 bp,马达加斯加彩蛙和大西洋盲鳗线粒体基因组长度分别为22 874 bp和18 909 bp。蛋白编码基因序列总长稳定在11.3 kb左右,tRNA基因和rRNA基因总长分别在1.5 kb、2.5 kb左右,物种间3 种基因的序列组成也较为接近[13]。本研究发现福建黄兔线粒体基因组序列全长17 000 bp,蛋白编码基因、rRNA、tRNA的长度及基因排列顺序与其他脊椎动物基本一致,基因排列紧密,间隔和重叠序列较少。基因组全序列具有明显的AT偏好性,A+T含量高于G+C含量的特点与已公布的其他兔形目动物相同[14-15]。福建黄兔13个蛋白编码基因起始密码子以ATG(10个)为主,终止密码子以TAA(8个)、TAG(3个)居多,终止密码子存在6个不完全终止密码子(T、TA或AG),通过转录后的多聚腺嘌呤机制补足为TAA或AGA终止密码子[16]。对密码子使用频率分析可知,福建黄兔编码基因中共有34个RSCU值大于1的偏好密码子,密码子第3位碱基中A、C占比明显高于G、T,这与密码子具有使用偏倚性相一致。此外,密码子第2位碱基多为嘧啶,这些特性决定编码基因多编码疏水氨基酸,由于线粒体编码蛋白主要是与内膜结合的呼吸链成分,因此疏水氨基酸的偏好性是与其功能相适应的[17-18]。
rRNA基因在蛋白合成过程中的二级结构决定其功能[19],茎区、环区结构中含有的重要亲缘信息,常用于系统发生的研究[20]。将福建黄兔rRNA序列结构与其他脊椎动物比对分析后发现,其rRNA结构相对保守,12sRNA与16sRNA相比更加保守,这一结果符合rRNA的进化规律。本研究中福建黄兔tRNA-Ser(AGY)因缺失DHU臂不能折叠成经典的三叶草结构,这在脊椎动物中比较常见。对DHU臂缺失现象的研究表明,tRNA在缺失DHU臂的情况下仍然可以形成潜在的L型结构来保证tRNA接受臂与反密码子之间的距离,从而维持其携带、转运特定氨基酸的功能[21-22]。
与其他哺乳动物mtDNA一样,L链复制起始区位于5个tRNA形成的基因簇中,在tRNA-Asn和tRNA-Cys之间,序列同源性较高,该区域可能形成稳固的茎-环结构。在福建黄兔中,利用RNAfold软件计算最小自由能表明L链复制起始区的二级结构非常稳定。D-loop区是线粒体基因编码的控制区,负责mtDNA的复制和转录,也是复制过程中mtDNA与线粒体内膜的结合位点。福建黄兔线粒体D-loop区在tRNA-Glu基因和tRNA-Phe基因之间,同绝大多数脊椎动物一样含有3个结构域,其中中央保守区较为保守,延长终止序列和保守序列区在碱基组成与长度上均表现出异质性[23]。D-loop区中短串联重复序列的拷贝数相对稳定,长串联重复序列因终止子缺乏容易发生拷贝数的变化[24]。D-loop区的重复序列拷贝数改变、序列插入和缺失等都是物种进化的表征[25]。将福建黄兔D-loop区序列与穴兔比对发现,福建黄兔缺少两个153 bp的长串联重复,这表明其属于进化的物种。
D-loop区富含A、T碱基,其碱基替换率较mtDNA其他部分高5~10倍[26],是线粒体基因组的高变区。D-loop区具有进化压力较小、遗传变异大、进化速度快的特点,被广泛用于物种间或物种内的进化分析[27]。本研究将测序所得的福建黄兔D-loop区序列与已公布的多个物种D-loop区序列进行多重比对后构建系统进化树,结果支持将福建黄兔的祖先归为穴兔。
本研究测定并分析福建黄兔线粒体基因组全序列,结果表明,福建黄兔线粒体基因组的组成、结构与其他脊椎动物相似,基因组全长17 000 bp,无内含子,基因排列顺序基本一致。对D-loop区的分析发现,福建黄兔线粒体也可以检测到3 个保守序列模块(CSB1, CSB2和CSB3)、2个终止序列(ETAS1和ETAS2)、重复序列RS3和RS2,不同的是RS2序列与穴兔相比缺少2个拷贝的LR重复片段。系统进化树分析亲缘关系的远近为研究福建黄兔的起源进化和特定遗传特性的形成机制奠定基础。
