向日葵粗脂肪及脂肪酸组分分析
2020-09-10雷中华黄启秀向理军
雷中华,黄启秀,张 黎,吴 伟,杨 辉,孙 逊,向理军
(1.新疆农业科学院经济作物研究所,乌鲁木齐 830091;2.新源县农业技术推广站,新疆新源 835800,3.特克斯县农业局,新疆特克斯 835500,4.昌吉州西亚种子有限责任公司,新疆昌吉 831100)
0 引 言
【研究意义】油用向日葵适应性强,耐旱,耐盐碱,抗贫瘠。向日葵种胚中的脂肪酸主要由棕榈酸(C16∶0)、硬脂酸(C18∶0)、油酸(C18∶1)、亚油酸(C18∶2)及亚麻酸(C18:3)等5种主要脂肪酸组成,其中油酸、亚油酸是向日葵种胚中主要的不饱和脂肪酸,含量占到了脂肪酸总量的50%以上[1]。不饱和脂肪酸具有特殊的营养特点[2,3]。长期食用高不饱和脂肪酸食用油具有保健作用[4]。研究向日葵脂肪酸以及脂肪酸组分所占比例,对油用向日葵品种的定向改良有重要意义。【前人研究进展】向日葵脂肪酸积累的影响因素有很多,既受自身生理发育的调控,又受外界环境因子的影响[5]。对向日葵基因型、播种时期,生育时期的温湿度、水分、光照等有研究表明,油菜花期降水过多,大量降雨使田间湿度增大,就会造成种子含油量的降低,呈现出含油量与降水量是负相关的关系;油菜在受旱的情况下适当灌水,能有效增加脂肪酸积累,提高种子含油量;在相同温度条件下,增加日照的时间能够提高种胚含油量的水平,在相同光照条件下,降低温度也会提高种胚含油量;早播延长了植株干物质的积累时间,对脂肪酸积累有利。而晚播植株因为错过了最好的播种时间,各方面生长受到限制,种子含油量相对就会有所降低[6-9]。【本研究切入点】油料作物向日葵脂肪酸含量以及脂肪酸各组分所占比例,是衡量其油用价值的重要标准,研究向日葵种胚形成时期的粗脂肪和脂肪酸的形成机制。【拟解决的关键问题】对种植在3个不同生态区的5个向日葵材料进行粗脂肪含量以及组分测定,通过粗脂肪总量的比较,以及不同材料在不同地区脂肪酸组分的不同,研究向日葵种胚发育过程中脂肪酸的形成规律,为向日葵调控脂肪酸的关健基因的功能提供依据,为油用向日葵品质的定向改良提供参考。
1 材料与方法
1.1 材 料
参试5个向日葵品种A17、567DW、KWS30、RITOM和TO1244由昌吉州西亚种子有限责任公司提供,生育期均在90~98 d。其中A17和RITOM为高油酸品种,油酸含量分别为82.3%和79.5%。,其余为普通品种。
1.2 方 法
1.2.1 试验设计
将5个向日葵材料分别在新疆3个不同生态气候区种植:乌鲁木齐(E 87.473,N 43.945,592 m)、伊犁新源(E 82.844,N 43.425,829 m)、阿勒泰北屯(E 87.976,N 47.274,550 m),每个点设置3个重复。
取开花后5、10、15、20、25、30和35 d 向日葵种胚,快速烘干后置于0~4℃冰箱保存,用于后续粗脂肪及脂肪酸组分的测定,并记录环境因子的变化情况和生育时期情况。
1.2.2 粗脂肪测定
测定采用索氏抽提的方法,试验仪器及试剂:索氏抽提器、干燥锅、石油醚、分析天平、恒温水浴锅、烘箱和粉碎机等。
索氏抽提:第一步将向日葵不同时期的种胚样品各称取5 g,放入纸袋中,编号排序,置于80℃烘箱中2 h,取出放入干燥器中备用。第二步,将滤纸包编号排序后置于105℃烘箱2 h,取出放入干燥器中备用,记为a。将第一步干燥器中的样品用粉碎机粉碎后均匀分成2份放入步骤2纸包中,封口后置于105℃烘箱中2 h,然后取出放入干燥器中,待其冷却后称重备用,记为b。将装有粉碎样品的纸包放入抽提筒中,加入石油醚至淹没纸包。将抽提器的各部分组装好,在70~80℃恒温中抽提24 h左右。抽提完毕后,取出纸包,置于通风环境下,使石油醚挥发。待乙醚挥发净后,将其放入105℃烘箱中2 h,取出放入干燥器中,至其冷却后称重,记为c。
脂肪含量(%)=(b-c)/(b-a)×100%。
1.2.3 脂肪酸各组分测定
棕榈酸(C16:0)、硬脂酸(C18:0)、油酸(C18:1)、亚油酸(C18:2)、亚麻酸(C18:3)等脂肪酸各组分利用气相色谱来进行测定。所用仪器为安捷伦气相色谱仪(Agilent 6890N 美国)。测定升温法采用程序升温法,测定的柱温210~230℃,升温速率10℃/ min,色谱仪的进样口温度为260℃,同时入口压强为25 Pa,高纯氮气25 mL/min,氢气40 mL/min,空气400 mL/min。
脂肪酸甲酯(FAME)的制备:将向日葵不同时期的种胚样品冷冻干燥至衡重,称重后研磨成粉状,加入石油醚和乙醚(v/v=1∶1)的混合溶剂l mL,再加入0.4 N KOH-甲醇溶液l mL,置于60℃温浴1 h进行甲脂化反应,冷却至室温后加入内标十九碳饱和脂肪酸甲脂,加入2 mL蒸馏水,震荡后离心,取上层清液,备做色谱分析。
以脂肪酸的绝对量(mg/g)和相对量(%)表示。根据各种脂肪酸的量计算脂肪酸总量(mg/g)。绝对量计算公式为Wi=Ai×ms/(As×m),其中Ai为第i种脂肪酸组分的峰面积,As为内标的峰面积,ms为内标的物质量,m为样品干重。相对量(%)=Ai/ ∑Ai×100。每个取样时间点计算3次重复的平均数和标准误,不同脂肪酸累积过程的相关性和主成分分析在脂肪酸绝对含量的基础上进行。
2 结果与分析
2.1 脂肪酸总量
研究表明,随着向日葵种胚的发育,脂肪酸总量稳步上升,但在种胚成熟末期,脂肪酸总量增长趋于缓慢,甚至在个别品种中出现回落趋势。相同品种在不同地区的脂肪酸积累趋势也有差别。
种植于阿勒泰地区的A17脂肪酸总量从15 d开始快速增长,持续至30 d进入积累平台期。而该品种在伊犁地区和乌鲁木齐从5 d就进入快速增长期,前者持续至25 d,后者持续至35 d,且后者的10~15 d出现了缓慢增长期。567DW的脂肪酸总量快速增长期开始时间与A17相同,只是前者的快速增长期的持续时间少于后者,这可能与A17是高油酸品种有关。图1
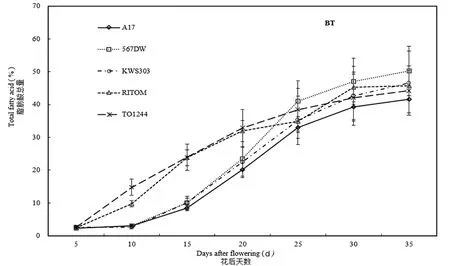
图1 阿勒泰地区种胚发育过程中脂肪酸总量的变化Fig. 1 Total amount of fatty acids during sunflower seed development in Altay region
种植于阿勒泰地区和乌鲁木齐的KWS303,脂肪酸总量均从花后10 d开始快速增长,前者持续至花后30 d,后者持续至花后35 d,且增长速率比较均匀。在伊犁地区该品种从花后5 d开始快速增长持续至花后25 d,增长速率参差不齐,且从花后30 d开始脂肪酸总量有所回落。RITOM在这3个地区的脂肪酸总量均从花后5 d开始快速增长,该品种种植在阿勒泰地区的脂肪酸总量增长趋势出现两个平台期,分别为花后20~25 d和花后30~35 d,伊犁地区快速增长持续至花后20 d,种植于乌鲁木齐的向日葵脂肪酸总量从花后25 d进入缓慢增长期。图2
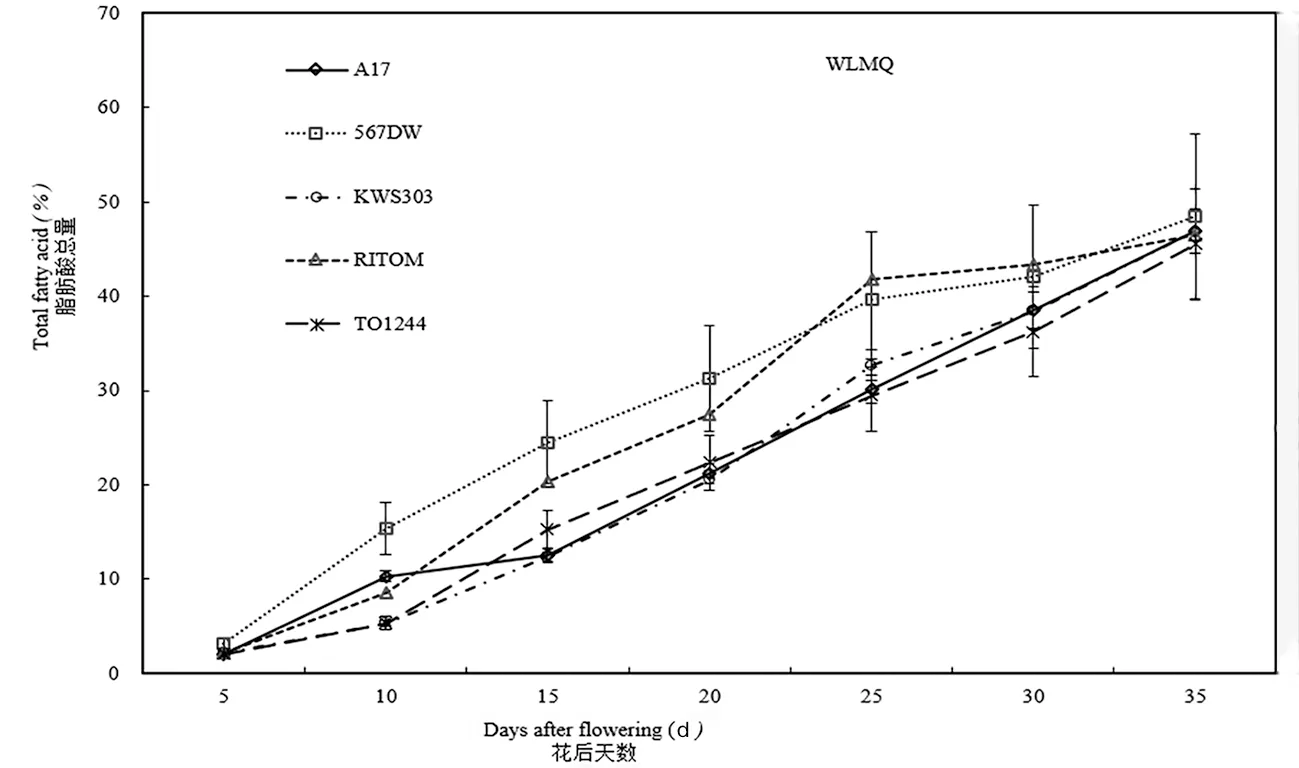
图2 乌鲁木齐种胚发育过程中脂肪酸总量变化Fig. 2 Total amount of fatty acids during sunflower seed development in Urumqi
研究表明,TO1244在阿勒泰地区和伊犁地区的脂肪酸总量均从花后5 d就开始快速增长,分别在花后20和25 d进入积累平台期,而种植在乌鲁木齐的该品种脂肪酸总量的快速增长期从花后10 d开始,且持续至花后35 d。图3
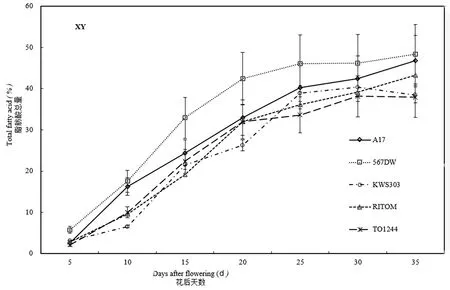
图3 伊犁地区种胚发育过程中脂肪酸总量变化Fig. 3 Total amount of fatty acids during sunflower seed development in Yili region
在乌鲁木齐种植的向日葵,脂肪酸总量快速增长期持续时间均高于其他2个地区,在乌鲁木齐地区种植的向日葵最终(花后35 d)脂肪酸总量均高于另外2个地区,向日葵脂肪酸的总量与种胚发育阶段脂肪酸总量的快速增长持续时间有关。表1
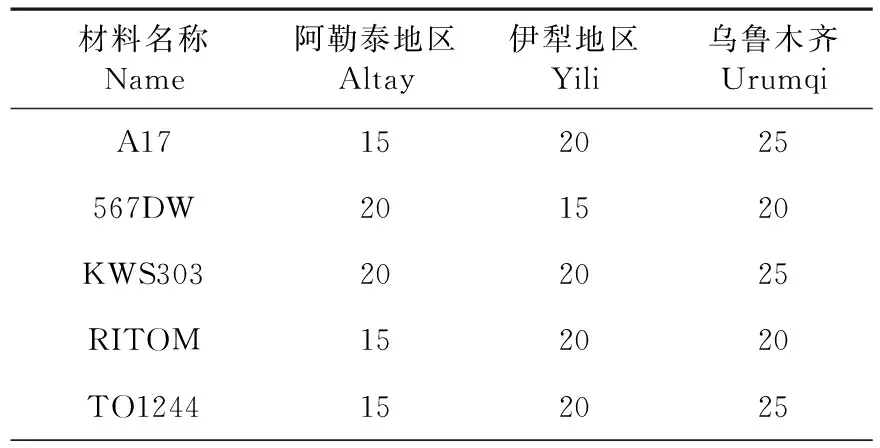
表1 快速增长期持续时间Table 1 Rapid growth period(d)
2.2 脂肪酸组分含量
研究表明,5个品种的棕榈酸、棕榈油酸、硬脂酸和花生酸含量均比较低,不饱和脂肪酸含量均达到90%以上,向日葵种胚属于高不饱和脂肪酸食品。向日葵种胚中含量最高的不饱和脂肪酸是油酸,其次是亚油酸和亚麻酸,在种胚中不饱和脂肪酸合成途径中油酸合成占主要部分。在花后5 d油酸含量极低,有的甚至并未检测到油酸,在花后10 d时检测到大部分品种的油酸含量显著增高,油酸的合成可能从花后5 d才开始。3个点的A17和除新源以外的RITOM,在花后10~30 d的油酸含量呈递增或保持高含量趋势。普通品种的油酸含量均在前期(花后5~10或15~20 d)增长较快,花后20 d开始油酸含量增长放缓,部分品种呈递减趋势,且含量均少于2个高油酸品种,高油酸品种可能在后期保持较高的油酸合成速率。伊犁新源点多个品种的亚油酸含量高于其他地区,而亚麻酸含量则较低。KWS303在北屯和新源点花后20 d显示高亚油酸特性,亚油酸含量均达到54%以上,但在乌鲁木齐该品种开花20 d后亚油酸含量均在45%以下。图4~6

图4 阿勒泰地区种胚发育过程中脂肪酸构成变化Fig. 4 Fatty acid composition during seed development in Aleatai region

图5 伊犁地区种胚发育过程中脂肪酸构成变化Fig. 5 Fatty acid composition during seed development in Yili region
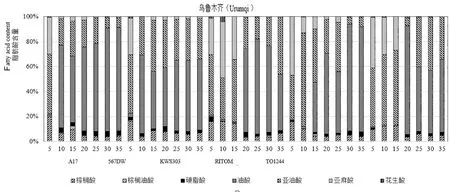
图6 乌鲁木齐种胚发育过程中脂肪酸构成变化Fig. 6 Fatty Acid Composition during seed development in Urumqi
2.3 脂肪发育过程中脂肪酸各组分的相互关系
研究表明,品种A17中硬脂酸和油酸呈正相关为第一类。其余脂肪酸为第二类,第二类中除棕榈油酸和亚麻酸、花生酸呈负相关以外,其余均呈正相关。品种567DW硬脂酸和油酸呈正相关,为第一类,其余脂肪酸为第二类。第二类中除棕榈油酸和油酸呈正相关外,其余均与第一类中的脂肪酸呈负相关。品种KWS303棕榈油酸、硬脂酸和油酸之间呈正相关属于第一类。棕榈酸、亚油酸、亚麻酸和花生酸之间呈正相关为第二类,第一类和第二类之间呈负相关。品种RITOM的硬脂酸和油酸呈正相关,为第一类。其余脂肪酸为第二类,第二类中除棕榈油酸和花生酸呈负相关外,其余脂肪酸之间均呈正相关。品种TO1244硬脂酸和油酸之间呈正相关为第一类。棕榈酸、棕榈油酸、亚油酸和亚麻酸之间互为正相关为第二类,花生酸为第三类,与第一类中的油酸和第二类中的棕榈油酸、亚麻酸呈正相关,与其他脂肪酸均呈负相关。各品种中第一类和第二类之间均呈负相关。A17和RITOM品种的脂肪酸各组分相关规律有一定的相似之处,其棕榈油酸均与油酸、硬脂酸和花生酸呈负相关,这种现象与这2个品种为高油酸品种有关。表2~4

表2 种胚发育过程中脂肪酸相关系数Table 2 Correlation coefficient of fatty acids during seed development
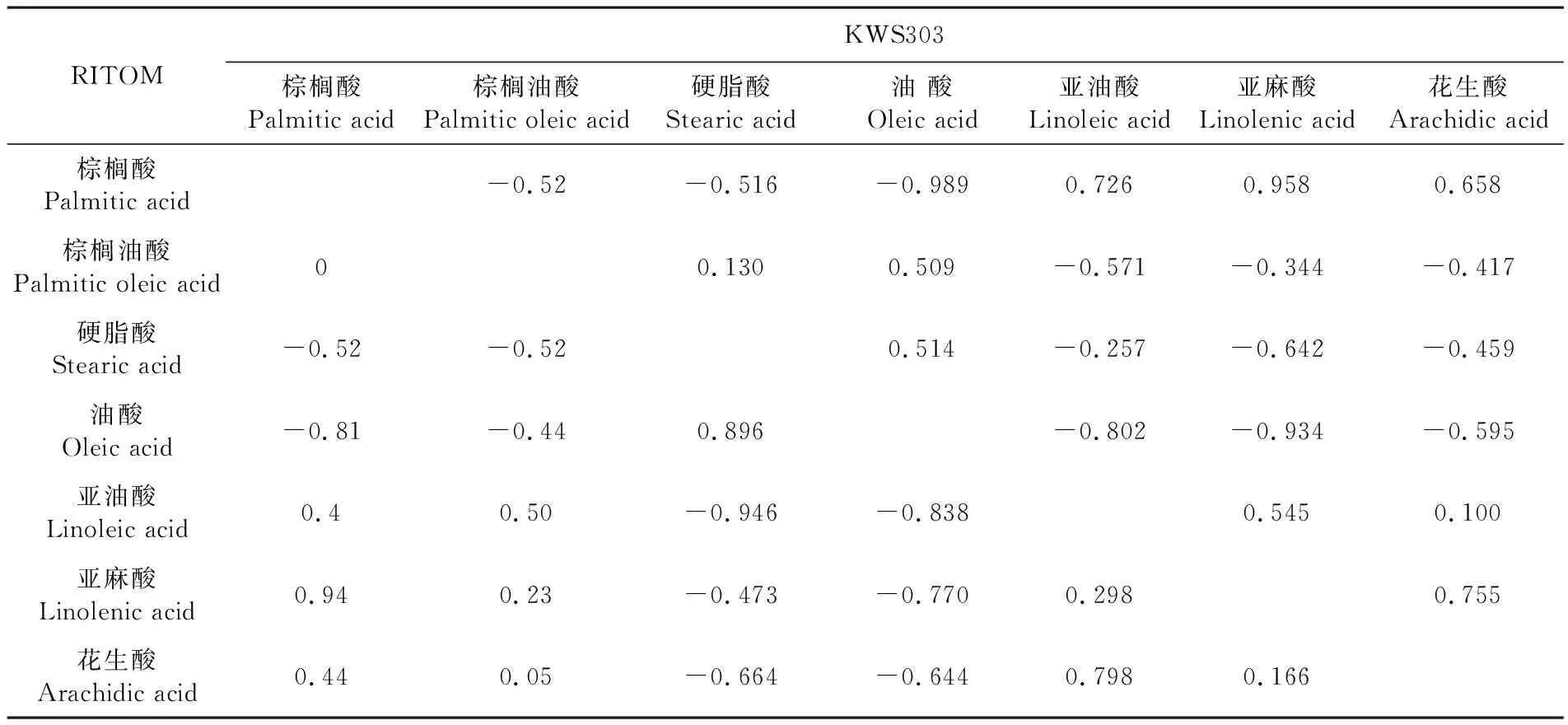
表3 种胚发育过程中脂肪酸相关系数Table 3 Correlation coefficient of fatty acids during seed development

表4 种胚发育过程中脂肪酸相关系数Table 4 Correlation coefficient of fatty acids during seed development
2.4 脂肪酸组分的主成分
研究表明,提取前2个主要成分,累积贡献率达74.237%,第一主成分的贡献率为58.793%,第二主成分的贡献率为15.444%。
其中第一个主成分的值与棕榈酸、亚油酸、亚麻酸和花生酸呈正相关,同硬脂酸和油酸呈负相关,对棕榈油酸的影响较小。棕榈酸、亚油酸、亚麻酸和花生酸的含量越高,则硬脂酸和油酸的含量会相应的减少。鉴于第一主成分中油酸的特征向量绝对值最大,故称其为油酸因子。
第二主成分与棕榈油酸和亚油酸呈正相关,与棕榈酸、硬脂酸、油酸、亚麻酸和花生酸呈负相关。由于花生酸在与第二成分呈负相关的脂肪酸中特征向量绝对值最大,所以花生酸可能对棕榈油酸和亚油酸的抑制作用较大,对其余脂肪酸的影响并不显著,可称第二主成分为花生酸指数。表5
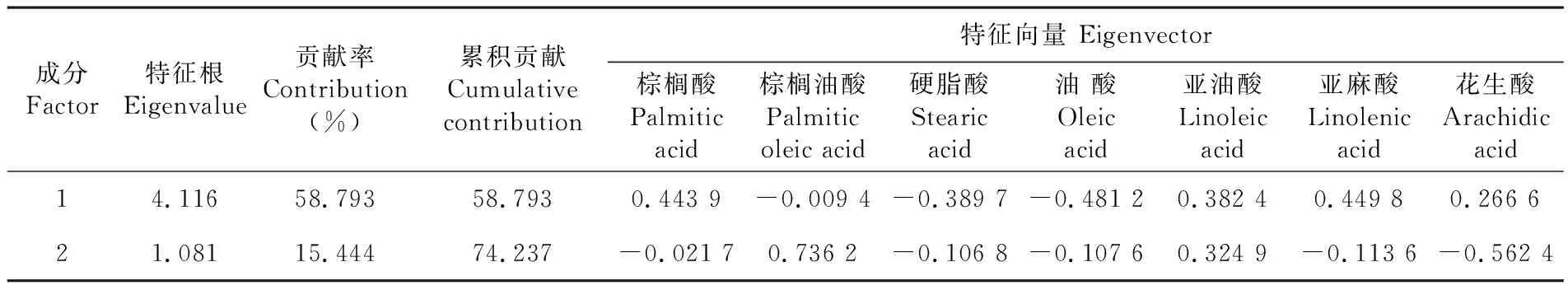
表5 种胚发育过程中脂肪酸主成分Table 5 Principal component analysis of fatty acids during seed development
3 讨 论
油料作物脂肪酸含量以及各组分所占比例,是衡量其油用价值的重要标准,研究油料作物脂肪酸形成规律有着重要的意义,不同油料作物的脂肪酸积累模式不尽相同。花生[10]、芝麻[11]和油菜[12]的脂肪酸总量积累模式呈现先上升至最大值再小幅下降的趋势,研究结果显示,向日葵种胚形成前期脂肪酸总量合成速率较高,在后期合成速率明显低于前期,这与花生、芝麻和油菜的脂肪酸积累模式有相似之处,造成这种现象的原因是种胚成熟后期水分丧失,植株体中营养运输困难,需消耗部分油脂[13],所以在种胚成熟后要适当提前收获,向日葵油脂含量之所以回落不明显也与适时收获有一定的关联。
研究还证实大部分品种在种胚发育前期的油酸合成速率要高于后期。前人研究证明,环境温度对种胚中油酸的积累有重要的影响,温度越高对种胚中油酸的积累越能起到积极的作用[14,15]。在向日葵种胚成熟后期,温度明显降低可能是导致种胚中油酸积累速率减慢的原因之一。RITOM是高油酸品种,在阿勒泰地区和乌鲁木齐均显示高油酸含量,但在伊犁地区油酸含量却远远低于这2个地区,可能与伊犁地区温度较低有关。
在种胚发育的不同时期,脂肪酸组成也有变化,最初脂肪酸有7种,到了种胚发育中期检测不到花生酸,种胚快成熟期,大多数品种的硬脂酸含量几乎为0,与芝麻[11]的积累模式有类似之处,但芝麻在中途消失的脂肪酸为棕榈油酸和亚麻酸。
油酸较其他不饱和脂肪酸更稳定,食用营养价值高,高油酸育种是目前向日葵育种的一大热点。研究发现硬脂酸和油酸呈正相关,油酸和亚油酸含量呈负相关,硬脂酸和油酸的合成是同步进行的,这与李晓丹[8]在花生中的研究结果相反,可能在不同的油料作物中油酸积累模式有差异。油酸因子主成分显示棕榈酸、亚油酸、亚麻酸和花生酸的含量越高,则硬脂酸和油酸的含量会相应的减少,这种结果提示育种家在高油酸育种过程中,提高油酸的同时可能会伴随产生高硬脂酸的结果。
4 结 论
4.1 随着向日葵种胚的发育,脂肪酸总量稳步上升,但在种胚成熟末期,脂肪酸总量增长缓慢,甚至在个别材料中出现回落趋势。向日葵最终脂肪酸总量与种胚发育阶段脂肪酸的快速增长持续时间密切相关。
4.2 分别测定3个点的5个品种不同时期的籽粒样品的脂肪酸组分含量:向日葵种胚属于高不饱和脂肪酸食品,向日葵种胚中含量最高的不饱和脂肪酸是油酸,其次是亚油酸和亚麻酸,在种胚中不饱和脂肪酸合成途径中油酸合成占主要部分。向日葵种胚油酸的合成可能从花后5 d才开始,高油酸品种可能在后期保持较高的油酸合成速率。
4.3 对向日葵中各脂肪酸组分含量进行相关分析,各品种之间的脂肪酸相关规律不尽相同,品种A17中硬脂酸和油酸呈正相关为第一类,其余脂肪酸为第二类;品种567DW中硬脂酸和油酸呈正相关,为第一类,其余脂肪酸为第二类;品种KWS303中棕榈油酸、硬脂酸和油酸之间呈正相关属于第一类,棕榈酸、亚油酸、亚麻酸和花生酸之间呈正相关为第二类。品种RITOM中硬脂酸和油酸呈正相关,为第一类,其余脂肪酸为第二类。品种TO1244中硬脂酸和油酸之间呈正相关为第一类,棕榈酸、棕榈油酸、亚油酸和亚麻酸之间互为正相关为第二类,花生酸为第三类。各品种中第一类和第二类之间均呈负相关。A17和RITOM品种的脂肪酸各组分相关规律有一定的相似之处,它们的棕榈油酸均与油酸、硬脂酸和花生酸呈负相关,这种现象可能与这2个品种为高油酸品种有关。
4.4 第一主成分为油酸因子,贡献率为58.793%,棕榈酸、亚油酸、亚麻酸和花生酸的含量越高,则硬脂酸和油酸的含量会相应的减少。第二主成分为花生酸指数,贡献率为15.444%,花生酸可能对棕榈油酸和亚油酸的抑制作用较大,对其余脂肪酸的影响并不显著。
