干旱荒漠区欧李品种光合生理日进程与产量
2020-09-08姚泽姜生秀严子柱刘光武
姚泽,姜生秀,严子柱,刘光武
(1.甘肃省治沙研究所/甘肃省沙生植物工程技术研究中心,甘肃 兰州 730070;2.甘肃省荒漠化与风沙灾害防治国家重点实验室,甘肃 兰州 730070;3.甘肃民勤荒漠草地生态系统国家野外科学观测研究站,甘肃 民勤 733000)
欧李Cerasus humilis属蔷薇科樱属落叶灌木,山西农业大学从野生欧李中培育出多个栽培品种,逐渐发展成为一种新兴果树[1]。欧李集果、花、药多种用途为一体,经济价值很高[2-4]。欧李耐瘠薄,抗寒,抗旱,栽培技术简单,是干旱半干旱地区防风固沙、水土保持、城市绿化的先锋树种[5]。
光合作用是一切物质和能量的来源,是作物高产的生理基础。目前,有关欧李的研究多集中在引种栽培[6]、形态解剖[7]及营养成分[8]等方面,而且多在干旱半干旱区进行,关于欧李光合特性与产量关系的研究还比较少。本文以干旱荒漠区4 个欧李品种为研究对象,比较不同月份光合生理日进程及其与产量的关系,以阐明其差异性,为干旱荒漠区欧李丰产栽培提供依据。
1 研究区概况
甘肃民勤治沙综合试验站,102°58′E、38°34′N,海拔1 378 m,属典型温带大陆性干旱荒漠气候。夏季酷热,冬季寒冷,日温差大,年均气温7.6 ℃,极端低温-30.8 ℃,极端高温40℃,≥10 ℃活动积温3 036.4 ℃;气候干燥,降水稀少,蒸发强烈,年均降水113.2 mm,年均蒸发2 604.3 mm,蒸发量是降水量的23 倍,空气相对湿度47%,空气干燥度5.1,最高18.7;光热充足,年均日照2 799.4 h,无霜期175 d;冬季盛行西北风,全年风沙日83 d,多集中在2~5 月,年均风速2.5 m/s,最大风速23.0 m/s。
试验地土壤为碱性沙土,沙层深厚,肥力很差,全盐0.146%,有机质0.198%,全氮0.008%,全磷0.116%,pH 8.3。
2 材料与方法
2.1 材料
供试欧李品种4 个:农大5 号、农大6 号、农大7 号、生态早熟种,苗木引自山西省太谷县欧李培育基地,每个品种500 株,苗龄2 a,栽植株行距0.8 m×1.0 m,2016 年定植。试验地土质条件基本相同,栽培技术和管理水平一致。
2.2 试验方法
LI-6400XT 便携式光合仪(LI-COR,USA)测定净光合速率、胞间二氧化碳浓度、气孔导度、蒸腾速率等,自然光源,透明叶室。每个供试品种定株测定5 株,样株长势一致;每株(每次或测期)定叶测量3 片样叶,样叶同枝(或异枝),梢顶下第5~8 片叶,消除了叶龄差异,样株无病虫害、长势一致。2019 年6~9 月选择晴朗天气测定光合参数,每月测定1 次,每次测定3 d,每天测定时间7:00~19:00,每2 h 测定1 次,每个样叶重复记录5 组数据,取平均值。
每月每次光合参数测定完毕后,定株观测株中选择3 株,每株同方位同层次取样叶5 片,密封袋带回室内,YMJ-C 叶面积扫描仪测定叶面积。
9 月10 日~15 日,每个供试品种随机抽取100 株逐株测定产量。生态早熟种8 月上旬至9月上旬果实成熟,其余供试品种果实开始成熟的日期较晚,依次为农大6 号、农大7 号、农大5 号。
采用Excel 对所测数据进行整理分析。所有数据采用Prisim 5.0 软件进行分析,采用单因素方差分析、Tukey 法检验和多重比较。
3 结果与分析
3.1 光合生理日进程
图1 显示,净光合速率日进程曲线,4 个供试品种的7 月线均高居上位,6 月、8 月、9 月线互相接近甚至交错,处于下位,说明供试品种7 月份的光合生产能力最高,在营养生长和生殖生长中贡献最大。4 个供试品种的6 月、7 月线均在13:00时出现低谷,两侧则凸起呈双峰状;9 月线峰谷现象消失,说明6 月、7 月份供试品种每日13:00 时出现光合午休现象,9 月份光合午休现象消失。7月线13:00 时的谷底,农大5 号显著高于其余月份同期测定值,农大6 号、农大7 号和生态早熟种则接近其余月份同期测定值,说明农大5 号光合午休程度较轻。
8 月、9 月欧李果实陆续成熟[9]。图1 显示,净光合速率日进程曲线,4 个供试品种的8 月、9月线差异较大,农大5 号8 月线呈微弱双峰状位居9 月线之上;农大6 号8 月线呈单峰状交错落于9 月线之下,农大7 号8 月线呈微弱双峰状位居9 月线之上并部分重叠,生态早熟种8 月线呈微弱双峰双谷状位居9 月线之下,其第2 峰与6月、7 月同期测定值接近。供试品种8 月、9 月净光合速率日进程曲线的上述差异,与果实成熟期紧密相关。生态早熟种果实8 月上旬开始成熟,成熟的果实不再积累光合产物,植物体可能因此下调光合水平,导致其8 月线低于9 月线。同理,农大5 号果实成熟期最迟,8 月份尚未成熟的果实继续积累光合产物,导致其8 月线显著高于9月,甚至高于6 月。农大6 号、农大7 号果实成熟期介于生态早熟种和农大5 号之间,其8 月、9 月线的高低关系也介于二者之间。总之,供试品种8 月、9 月光合生理的上述变化和差异,在研究区欧李生产中具有一定意义,值得予以关注。
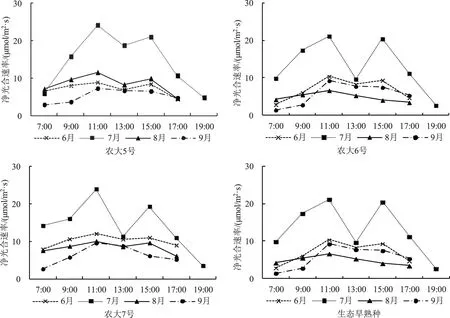
图1 净光合速率历月日进程
蒸腾是植物必不可少的生理过程,其主要生理作用在于降低植物体温、增加植物水分吸收和增加无机离子的吸收和运输[10]。图2 显示,蒸腾速率日进程线,4 个供试品种7 月线均呈弧状单峰线高居上位,并且均在13:00 时达到或接近最大值;6 月线均在13:00 时出现微弱谷底;8 月、9月线均在13:00 时前达到高峰。农大号5、农大7号各月线相对集中;农大6 号和生态早熟种各月线则比较分散,其中8 月线显著低于其他月线。农大5 号、农大6 号、农大7 号各月线均高于生态早熟种。说明:8 月份欧李品种间蒸腾日变化存在显著差异;相对于其他供试品种,生态早熟种测试期内蒸腾量较小。
图3 显示,气孔导度日进程曲线,农大5 号、农大6 号、农大7 号的6 月、7 月线均呈双峰单谷状且谷底出现在13:00 时,9 月线均呈单峰状且峰顶出现在11:00 时;8 月线,各供试品种均处于较低位置,但农大5 号呈双峰状且谷底出现在13:00时,农大6 号、农大7 号呈单峰状且峰顶出现在11:00 时;生态早熟种各月线均呈单峰或微弱双峰状,峰顶出现在11:00 时或9:00 时。农大5 号、农大6 号、农大7 号各月线均高于生态早熟种。说明:8 月份欧李气孔开放程度较小且品种间变异较大;相对于其他供试品种,生态早熟种测定期内气孔开放程度较小。

图2 蒸腾速率历月日进程

图3 气孔导度历月日进程
图4 显示,胞间二氧化碳浓度日进程曲线,各供试品种均呈两边高中间底的凹线,6 月线均相对较高;7 月、9 月线在各供试品种中变化较大但均位于6 月线上下;8 月线,农大5 号、农大6 号和生态早熟种均处于最低位置,谷底甚至接近0值,农大6 号呈单谷状,农大5 号和生态早熟种则呈双谷状,但农大7 号的8 月线接近6 月线处于较高位置。说明:8 月份欧李胞间二氧化碳浓度日变化剧烈,品种间变化方式存在明显差异,其他月份表现为先降后升。

图4 胞间二氧化碳浓度历月日进程
综合考察图1、图2、图3、图4 可以看出,各供试品种7 月净光合速率日进程曲线显著高于其余月线且13:00 时的峰间谷底较深,午休现象明显,但蒸腾速率、气孔导度和胞间二氧化碳浓度日进程曲线相应变化并不明显,7 月线相比其余月份高低位置和13:00 时的测定大多并不突出,只有农大6、7 号的气孔导度日进程曲线在13:00时的谷底比较显著。说明午休现象除受气孔开放和胞间二氧化碳浓度影响外,可能还与属于干旱荒漠气候的研究区7 月份光照强烈有关。8 月份各供试品种的4 个日进程曲线存在较大差异,表现出比较明显的品种特性,暗示这种品种特性与果实成熟有关,因为生态早熟种和农大6 号果实在8 月陆续成熟,农大5 号、农大7 号果实则在8下旬开始成熟。生态早熟种胞间二氧化碳浓度日进程曲线与其余供试品种相近,净光合速率、蒸腾速率和气孔导度日进程曲线则相对较低,说明生态早熟种对干旱荒漠气候适应能力较强,有较多的光合产物用于调节其生态适应性,导致净光合产物较低。
3.2 叶和果实大小
由表1 可以看出叶面积在农大6、7 号之间无统计学差异,两者其测定数值的差异源于随机因素;农大5 号、生态早熟种与农大6 号、农大7 号,三者互相之间均有统计学差异,其测定数值的差异不是随机误差所致,而是品种特性所致。说明:在叶片大小方面,4 个供试品种可以分为3 组,农大6 号、农大7 号为1 组,农大5 号和生态早熟种各为1 组。表1 数据显示,农大5 号叶面积测定数值最大,生态早熟种最小,农大6 号、农大7 号居中,说明叶片农大5 号>农大6 号、农大7 号>生态早熟种。
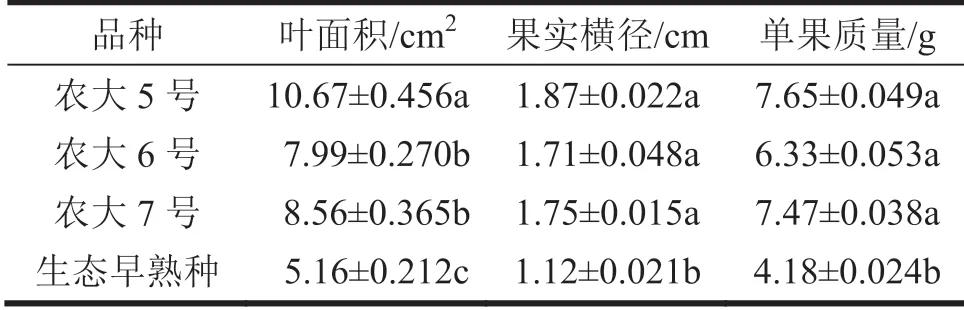
表1 供试品种叶和果实大小
由表1 可以看出,果实横径和单果质量,农大5 号、农大6 号、农大7 号3 个供试品种之间均无统计学差异,说明其测定数值的差异源于随机因素;生态早熟种与其余3 个供试品种均有统计学差异,说明其测定数值的差异源于品种差异。说明:在果实大小方面,4个供试品种可以分为2组,农大5 号、农大6 号、农大7 号为1 组,生态早熟种为1 组。表1 数据显示,农大5 号、农大6 号、农大7 号果实横径和单果质量测定值较大,生态早熟种测定值较小,说明生态早熟种果实较小。
总之,农大5 号、农大6 号、农大7 号叶片和果实均比生态早熟种大,说明前3 个供试品种栽培性状更明显,生态早熟种则带有较强的原始性状。
3.3 产量
图5 显示,农大5 号、农大7 号和生态早熟种结果株率接近或超过95%,农大6 号结果株率较低,不足90%。
图5 显示,单株产量,农大5 号与农大7 号之间、农大6 号与生态早熟种之间无统计学差异,说明其单株产量测定数值的差异源于随机误差;农大5 号、农大7 号与农大6 号、生态早熟种之间有统计学差异,说明其单株产量测定数值的差异源于品种差异。说明:在果实产量方面,4 个供试品种可以分为2 组,农大5 号、农大7 号为一组,农大6 号、生态早熟种为一组。图5 数据显示,农大5 号、农大7 号单株产量测定值较大,农大6 号、生态早熟种测定值较小,说明农大5 号、农大7 号果实生产能力优于农大6 号、生态早熟种。4 个供试品种中,农大6 号单株产量标准误明显较大,说明单株产量变异幅度较宽,生产稳定性较差。
农大6 号成熟期较早,结果株率和单株产量均较低;果实成熟前的8 月份净光合速率较低,而蒸腾速率却较高,光能和水分利用率均较低。因此,农大6 号经济性能较差。农大5 号7 月份光合午体现象较轻,叶片和单果质量均较大,结果率较高,其经济性能优于农大7 号。
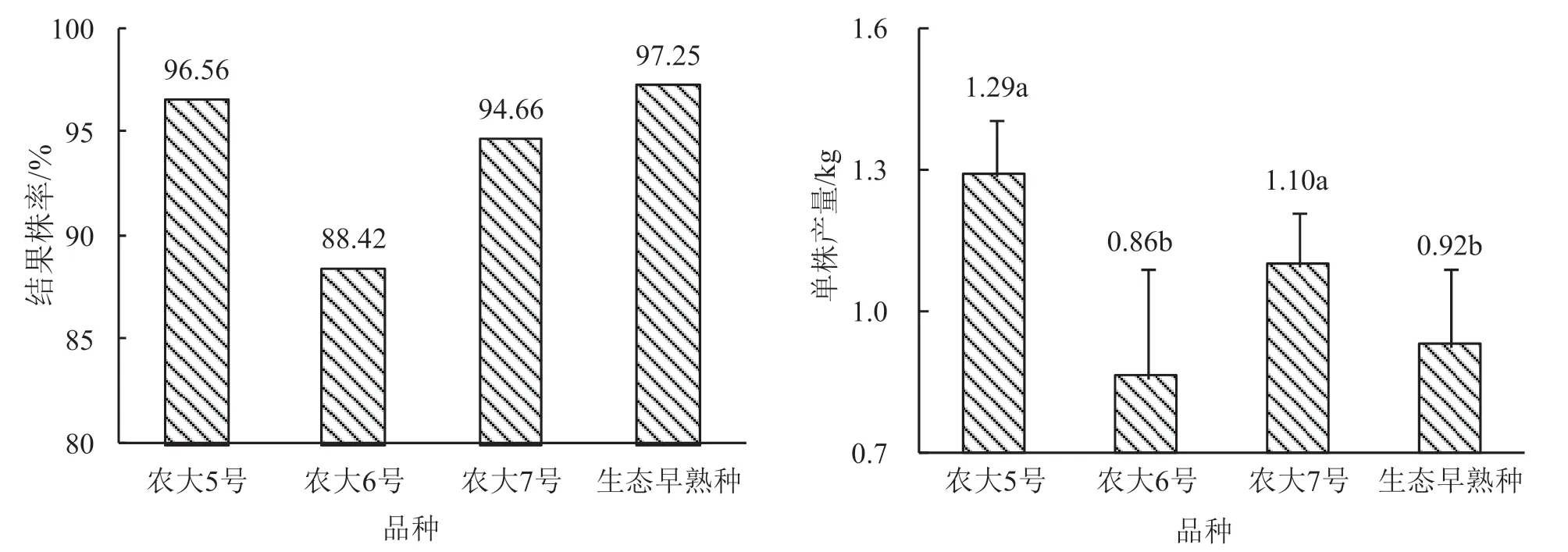
图5 供试品种结果情况
4 结论与讨论
农大5 号、农大7 号2 个品种产量较高,可以在与民勤相似气候类型的干旱荒漠区欧李栽培生产中推广应用,其中以农大5 号为优,栽植3 a 平均单株产量1.29 kg;生态早熟种对于干旱荒漠气候适应性较强,产量较低,可以适当用于相同气候区欧李生态化栽培生产。
4 个欧李品种在7 月份净光合速率最高,在营养生长和生殖生长中贡献最大,但光合午休现象比较明显,在欧李栽培中应加强7 月份的水肥管理,以削弱光合午休效应,提高产量。
8 月份,4 个欧李品种的净光合速率、蒸腾速率、气孔导度和胞间二氧化碳浓度日变化表现出比较明显的品种特性,推测与果实成熟期有关,由于在8 月份,民勤干旱荒漠区欧李果实陆续成熟,其光合生理对产量有一定影响。因此,值得深入研究欧李不同品种光合生理现象。
在自然条件下,植物的光合特征与光合有效辐射、蒸腾速率、气孔导度、胞间二氧化碳浓度等因子密切相关[11],净光合速率与气孔导度、蒸腾速率在一定范围内变化是一致的[2]。从本研究结果可以看出,4 个欧李品种在不同月份的净光合速率、气孔导度、蒸腾速率变化趋势基本一致。蒸腾速率主要受大气温度和空气相对湿度的影响,4个欧李品种蒸腾速率不断升高,在11:00 和15:00左右出现高峰。因为气温最高、光强最强的时段是在11:00~15:00,高温和强光导致叶温迅速升高,叶片内外蒸气压梯度增加,蒸腾速率加快,表皮细胞和保卫细胞直接向大气蒸腾水分,迫使整个叶片水势下降,气孔阻力增大[12]。气孔导度是植物气孔传导二氧化碳和水汽的能力,气孔导度越大,代表气孔开张越大,植物通过改变气孔的开张度等方式来控制与外界的二氧化碳和水汽交换,从而调节光合速率和蒸腾速率,以适应不同的环境条件[13]。欧李胞间二氧化碳浓度变化趋势与净光合速率相反,呈“W”和“V”字形曲线。通常光合速率的降低有两方面的原因:气孔限制和非气孔限制,根据Farquhar 等的观点,如果净光合速率和胞间二氧化碳浓度变化方向相同,并且气孔导度降低时净光合速率的降低是气孔因素造成的;如果净光合速率和胞间二氧化碳浓度变化方向相反,并且气孔导度增大,则净光合速率的下降主要是由非气孔限制,即叶肉细胞同化能力的降低引起的。可见,6~8 月份4 个欧李品种光合午休现象是非气孔因素造成的。
光是影响植物光合作用的重要因子,不同月份下光强的变化对欧李品种光合特性有较大的影响[14]。4 个欧李品种在7 月份均表现出较强的光合作用能力,7 月日均净光合速率均最高,均显著高于9 月份,但农大7 号9 月日均净光合速率大于8 月,这是因为8 月出现光合午休现象。4 个月份中农大7 号的日均净光合速率均最高,农大7 号和农大5 号之间无显著性差异,均显著高于农大6 号,生态早熟种高于农大6 号。由此可以看出,农大7 号和农大5 号光合作用性能较好,光合生理具有一定的可塑性,能够充分地利用当地的光照条件。
