嗜热性木质纤维素酶在纤维素乙醇生产中的研究进展
2020-08-28刘登刘均洪
刘登 刘均洪
(1. 山东理工职业学院化工学院,济宁 272067;2. 青岛科技大学化工学院,青岛 266042)
随着全球工业化进程的推进,能源需求越来越大,而传统化石燃料的储备却日渐枯竭,加上全球范围内温室效应的加剧和环保压力的增加,各国正不断寻求化石燃料的替代能源。生物乙醇因其原料广泛性、可再生性、环境友好性以及技术可行性逐渐受到各国科研人员的重视,被认为是传统液体燃料最有潜力的替代产品[1]。目前,生物乙醇已经在全球实现了商业化,尤其是美国和巴西,不过,商业化的生物乙醇现阶段主要是以可食用的淀粉基原料为底物,这在一定程度上造成了粮食危机,当非洲等贫困地区还存在大量饥饿人口时,淀粉基生物乙醇的生产和应用便受到各国政府的限制。而纤维素乙醇是以不可食用的木质纤维素为原料,其全球储量巨大,价格便宜且可再生,不会造成任何社会和经济问题,这使得纤维素乙醇未来的商业前景极其广阔。
然而,目前纤维素乙醇过高的生产成本在很大程度上限制了它的商业化进程。从纤维素乙醇生产的工艺经济可行性分析发现,造成其生产成本过高的原因,主要是在木质纤维素原料的生物乙醇转化过程中存在许多技术瓶颈,这些技术瓶颈不仅造成了大量的能源或试剂消耗,还导致部分原料的浪费和乙醇收率的严重下降[2]。为了解决这些问题,经济有效的生物催化剂(木质纤维素酶)的选择和利用被认为是纤维素乙醇未来的关键。可是纤维素乙醇规模化生产中的极端条件又极大限制了常规嗜温酶的活性和稳定性。针对这种状况,嗜热性木质纤维素酶因其高催化活性、热稳定性和对化学变性剂的抵抗力等独特优势引起了科研人员的极大关注[3]。本文将从如何提高嗜热酶产量、活性和稳定性方面介绍嗜热性木质纤维素酶在纤维素乙醇生产应用中的研究进展。
1 纤维素乙醇生产中存在的技术瓶颈
木质纤维素是地球上储量丰富且可再生的生物质原料,作为纤维素乙醇的原始底物潜力巨大[4]。从木质纤维素原料到生物乙醇的生物转化技术路线早已证实是可行的,纤维素生物乙醇也被认为具有广阔的商业应用前景。然而,至今为止纤维素乙醇的市场占有率却非常低,Elsayed等[5]通过对纤维素乙醇生产流程的经济性分析发现,过高的预处理运营成本是影响其经济可行性的关键因素。研究表明,纤维素乙醇生产的工艺路线虽然已经打通,但生物转化过程中却依然存在着许多技术瓶颈,这些技术瓶颈一方面制约着生物乙醇的底物转化率和产品收率;另一方面也导致了它的生产成本一直居高不下[6]。
木质纤维素生物质从结构上来看,主要是由纤维素(35%-50%),半纤维素(30%-35%)和木质素(10%-40%)组成[7]。其中,纤维素是由D-葡萄糖单元通过是由β(1→4)糖苷键组成的高分子线性均聚物,可以通过3种酶(内切葡聚糖酶、外切葡聚糖酶和葡萄糖苷酶)实现它的完全解聚,是目前产纤维素乙醇微生物的主要发酵底物;半纤维素是由不同类型的五碳糖和六碳糖构成的复杂异质高聚物,它的解聚需要多种酶(如木聚糖酶、阿拉伯糖苷酶、阿魏酸酯酶和甘露聚糖酶等)的联合作用,主要水解产物为以木糖为主的五碳糖,在现阶段纤维素乙醇的生产中利用率较低;木质素则是由3种主要单体(对香豆醇、松柏醇和芥子醇)形成的复杂酚类聚合物,其水解通常也涉及3种主要的酶(木质素过氧化物酶、锰过氧化物酶和漆酶),其水解产物无法被产醇微生物利用。在木质纤维素生物质中,纤维素多糖链之间通过氢键彼此连接形成结晶纤维素,半纤维素和其他多糖特别是果胶再与结晶纤维素形成胶结层,该结构最终再被木质素包围形成微纤维,许多这样的微纤维一起构成木质纤维素[8]。
木质纤维素具有上述致密的刚性结构,其在作为纤维素乙醇的原料使用时必须进行适当地预处理,通过破坏半纤维素和木质素形成的物理屏障以提高结晶纤维素的表面可及性。在纤维素乙醇的生产过程中(原料收集、预处理、水解糖化、发酵和蒸馏),原料的预处理目前尚存在许多技术瓶颈。通常的预处理方法包括物理法、化学法和生物法,这些技术方法主要涉及到酸、碱、氨纤维爆炸、离子液体处理、有机溶剂、臭氧分解、蒸汽处理、湿式氧化、微波辐射和酶等。然而,高昂的运营成本、大量溶剂的使用、抑制剂的形成、生物活性的损失、戊糖的浪费、产物抑制以及乙醇收率的降低,都限制着纤维素乙醇的商业化部署[9],尽管发现酶法降解木质纤维素是比物理法和化学法更有效的方法,但中温酶对工业条件的不适应性和高成本限制了它的应用[10],这迫使科研人员不得不继续寻求更加经济高效的预处理方法。于是,近几年嗜热酶便逐渐引起了各国研究人员的关注。本文将根据最新文献研究对嗜热性木质纤维素酶的特点和研究进展进行重点介绍。
2 嗜热性木质纤维素酶的特点
生物法预处理木质纤维素转化为可发酵糖的过程中,涉及到一整套复杂的木质纤维素酶,它包括纤维素酶系、半纤维素酶系和木质素酶系,每组酶系又是由许多具有不同功能和底物特异性的酶构成,如内切葡聚糖酶、外切葡聚糖酶、葡萄糖苷酶、木聚糖酶,阿拉伯糖苷酶、甘露聚糖酶、锰过氧化物酶和漆酶等。木质纤维素便是在这些酶的共同作用下将纤维素释放出来并转化为可发酵糖和其他降解产物的。然而,由于工业生产中的高温和极端pH等苛刻条件,使得常规嗜温性酶系的活性和稳定性都受到限制[11],为了改善这种状况,科研人员对嗜热性酶的研究逐渐增多。
嗜热性酶来自于嗜热微生物,能耐受80-120℃的高温和广泛的pH(0.5-11)值,这种特性允许生产过程可以维持在相对较高的温度下进行。在较高的温度下,底物溶解度增大、反应速率提高、杂菌污染几率下降、挥发性物质的快速挥发减少了对发酵微生物的产物抑制、乙醇收率增加、避免了中间过程的冷却处理、节约了成本、缩短了反应时间[12]。另外,嗜热性木质纤维素酶在高温下能够保持良好的催化性能和稳定性、具有一定程度的抑制剂耐受性。基于这些优点,研究人员相信嗜热性木质纤维素酶的应用有助于纤维素乙醇生产的工艺经济性[13]。
然而,受限于嗜热性微生物苛刻的生存条件和培养规模,目前嗜热性木质纤维素酶的产量无法满足纤维素乙醇生产的工业化需求,加上工业化生产环境与其最适工作条件具有偏差,嗜热性木质纤维素酶的活性和稳定性也受到影响[14]。所以,对于科研工作者来说,选择合适的方法提高嗜热性木质纤维素酶的产量、增大酶的活性和稳定性、降低它的生产成本便是下一步的研究重点。
3 纤维素乙醇生产中嗜热性木质纤维素酶的研究进展
利用嗜热性木质纤维素酶生产纤维素乙醇,既能克服嗜温酶在工业化生产中存在的各种缺陷,又能增大底物溶解率、加快反应速率、降低杂菌污染风险、节约系统冷却成本以及提高产品收率,因此展现出了巨大的商业应用潜力[15]。然而,受限于它的高成本,如何提高嗜热性木质纤维素酶的产量、活性和稳定性以加速其商业化部署便成为了科研人员的主要研究方向。图1展示了目前嗜热性木质纤维素酶在纤维素乙醇生产应用中的主要研究热点。
3.1 嗜热性木质纤维素酶的筛选
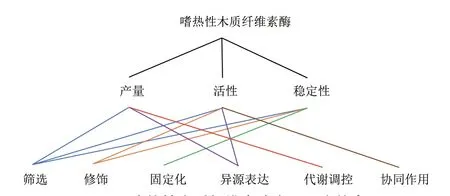
图1 嗜热性木质纤维素酶主要研究热点
嗜热性酶来源于嗜热性微生物,为了获得优良的嗜热性微生物菌株,研究人员采用了多种筛选策略。常规做法就是从各种不同的极端环境进行微生物的分离和鉴定,Saurabh等[16]从印度Leh和Ladakh地区的温泉水中分离出11种能够降解纤维素的细菌菌株,其中Bacillus subtilisBHUJPV-H19 在最适温度50℃时的β-葡萄糖苷酶活性为5.65 U/mL;Aikawa等[17]从牛粪堆肥中分离出一株富含糖苷家族GH11、GH2以及木聚糖酶的菌株Herbivorax saccincolaA7,它在pH 9和55℃时表现出较高的木质纤维素分解活性;Tan等[18]从牛粪便中筛选出一株Chryseobacterium属的嗜热性菌株,该菌株能产生具有纤维素酶-木聚糖酶双功能活性的嗜热性酶,在pH 9和90℃下对羧甲基纤维素的内切葡聚糖酶活性为3 237 U/mg,在pH 8和90℃下对桦木木聚糖的木聚糖酶活性为1 793 U/mg,将该酶作用于由稻草组成的蘑菇培养底物时,与某商业纤维素酶和木糖酶相比,葡萄糖产量是后者的9.2-15.7倍,木糖产量是后者的3.5-4.3倍;Shahi等[19]从黄石国家公园的酸性温泉中分离出一株Acidothermus cellulolyticus11B,该菌株能产生一种具有木聚糖内切酶、阿拉伯呋喃糖苷酶和乙酰木聚糖酯酶活性的三功能酶,该酶在70℃和pH 6.0时表现出很高的活性。上述研究表明,通过传统的筛选方法可以从极端环境中获取到具有优良性能的嗜热性木质纤维素酶,但是这种方法筛选过程比较繁琐且筛选通量较低,为了提高筛选效率,研究人员开发出了一种高通量的筛选方法即宏基因组筛选。Ariaeenejad等[20]根据骆驼瘤胃宏基因组学数据,使用五阶段计算筛选程序从骆驼瘤胃大量微生物菌群酶中快速找到了具有很宽pH值(6-11)和温度(30-90℃)范围的木聚糖酶。有研究人员表示,筛选性能优良的嗜热性微生物菌株是获得具有高产量、高活性和热稳定性嗜热性木质纤维素酶的前提。所以,在大自然这种独特而多样性的进化环境中,坚持传统的野生菌株筛选始终是一种不可或缺的手段。同时,开发和应用宏基因组学和现代测序技术等高通量筛选策略,探索嗜热性微生物群落多样性的微生物环境,快速预测和鉴定样品中所有微生物及其酶的种类,将极大提高筛选效率,有助于在较短时间内发现一批全新的嗜热性木质纤维素酶,从而强化其在纤维素乙醇生产中的应用[21]。
3.2 嗜热性木质纤维素酶的修饰
在纤维素乙醇的工业化生产中,极端的环境条件使工作用酶活性和稳定性都大幅度下降,这在一定程度上增加了纤维素乙醇的生产成本。研究人员发现,通过对酶的遗传或化学修饰可以提高其催化活性或改善因pH、热或抑制剂等引起的稳定性和寿命的下降,从而可以间接降低用酶成本。Brogan等[22]通过表面阳离子化反应对嗜热性葡糖苷酶进行化学修饰,然后将阳离子化葡糖苷酶与表面活性剂乙醇酸聚乙氧基月桂醚结合制备了一种酶-聚合物表面活性剂纳米结构,经修饰的葡糖苷酶可以将热稳定性提高到137℃,对纤维素的酶活提高了30倍。Darby等[23]将结构引导的小分子激活剂整合到一个特定位点以共价修饰Bacteroides thetaiotaomicron的糖苷水解酶BtGH84,结果表明,修饰后的酶催化活性提高了35倍,同时还增强了对某些抑制剂的耐受性。Boonyapakron等[24]在嗜热性木聚糖酶Xyn12.2的β-折叠核心的N末端和α-螺旋之间引入了两个二硫键,修饰后的酶在pH 9.0、50℃时催化效率增加了4.2倍,60℃时增加了3倍。Tajwar等[25]将嗜热性Clostridium thermocellum的2个碳水化合物结合模块(CBM6和CBM22)分别融合到由Thermotoga maritima产生的木聚糖酶XynB的催化域(XynB-C),结果显示,修饰后的酶XynB-CB22和XynB-CB6对不溶性桦木木聚糖的活性分别提高了3.24倍和2倍60℃下持续4 h后,前者的活性保持100%,后者活性损失11%。因此可以确定,从微生物中获取的天然嗜热性酶或常温酶,利用科学的方法进行适当改性后,其活性和稳定性将表现为更加有利于工业化生产,这在商品酶产量无法增加,价格无法下降的前提下,提高了它的性价比,相当于间接的降低了纤维素乙醇的生产成本。将来,在深入研究嗜热性酶的蛋白质晶体结构的基础上,构建基于结构和序列的嗜热性木质纤维素酶模型,将大大加快特定残基和结构域的靶向修饰,这种方法在改善嗜热性或嗜温性酶的特性方面展现出了广阔的发展空间。
3.3 嗜热性木质素纤维素酶的固定化
在生物催化过程结束后,酶的回收率是影响产品成本的另一因素。研究表明,固定化酶比游离酶具有更好的可回收性。此外,酶的固定化也已被证明是在低于最佳条件下提高酶活性和稳定性的有效方法之一。有研究人员将漆酶固定在碳纳米管上,结果固定化后的漆酶催化活性大幅度提高。Fernández-Fernández等[26]将 由Myceliophthora thermophila产生的漆酶固定在乙醛-琼脂糖珠上,从而提高了其在70℃下的活性和热稳定性。Gupta和Bisaria[27]将3种 商 业 化 的 内 切 葡 聚 糖酶(2 800±100 IU/mL),β-葡萄糖苷酶(570±25 IU/mL)和木聚糖酶(4 250±150 IU/mL)通过戊二醛化学交联固定化制备成交联酶聚集体(Cross-linked enzyme aggregates,CLEA),与游离酶相比,CLEA更稳定,在70℃时它们的滤纸酶活性(Filter paper enzyme activity,FPA)、内切葡聚糖酶、β-葡萄糖苷酶和木聚糖酶的半衰期分别比游离酶高出2.30、1.56、3.07和1.67倍。在以羧甲基纤维素和木聚糖为底物的水解过程中,每次水解持续24 h,温度控制在70℃,经过5个连续循环后,CLEA的总体糖化产率为43.3%,而游离酶为31.8%,另外5个循环后CLEA还保留了77.4%的内切葡聚糖酶和85.9%的木聚糖酶活性。同样,Periyasamy等[28]也把木聚糖酶、纤维素酶和β-1,3-葡聚糖酶进行了交联固定化,结果表明,固定化酶具有了更高的热稳定性、回收率和底物水解率。另外,Jia等[29]开发了一种新型的磁性交联纤维素固定化酶聚集体,并将其用于竹子木质纤维的生物转化,与游离纤维素酶相比,固定化纤维素酶在更宽的温度范围和pH值下显示出了更好的活性,并且每次水解反应完成后,都可以用永磁体对固定化酶分离回收,经过6次水解循环后,其活性仍能保持初始活性的74%。所以,在嗜热性木质纤维素酶产量有限、价格过高的现状下,利用酶的固定化技术不仅能够一定程度上提高酶的活性,还能大大提高酶的利用率,在目前以及将来都不失为一种有效而经济的方法。对于科研人员而言,选择或开发具有生物亲和性的交联载体、研究有利于嗜热性酶特性交联度、探索具有最佳协同作用的交联酶聚集体便成为了纤维素乙醇能源领域下一步的研究重点。
3.4 嗜热性木质纤维素酶的异源表达和代谢调控
3.4.1 嗜热性木质纤维素酶在嗜温性宿主中的异源表达 由于嗜热微生物的最佳环境条件很难在实验室中模仿,无法进行大规模培养,对于工业应用而言,天然产生的嗜热性酶的产量通常太低,因此将嗜热性木质纤维素酶在合适的微生物宿主中异源表达非常 必 要。Benedetti等[30]从Thermotoga neapolitana基因组中克隆了两个编码半纤维素酶的基因,并在Escherichia coli中表达,结果不但在正常的培养条件下提高了酶的产量,酶的活性和最适温度也有所提高。Haq和Akram[31]克隆了超嗜热真菌Thermotoga naphthophilaRKU-10T的 内 切1,4-β-木 聚 糖 酶(TnXynB)基因,构建了表达载体pET-21a-TnXynB,并使之在Escherichia coliBL21 CodonPlus中高效稳定的过表达,当经IPTG(0.5 mmol/L)热激(42℃21 h)诱导培养(0.8 OD600nm)后,于22℃摇床(200 r/min)培养72 h,观察到最佳的细胞外TnXynB表达,异源表达的TnXynB蛋白经纯化23.38倍后,以桦木木聚糖为底物时,在95℃和pH 6.0时表现出最佳催化活性。热稳定性分析表明,异源表达的TnXynB在温度50-85℃、pH 5.5-8.0的条件下保持8 h活性没有任何损失,而在pH 4.0-4.5时稳定性较差。Han等[32]将来自Chaetomium thermophilum的嗜热性纤维二糖水解酶CtCel6经合理设计后,使之在Pichia pastoris中过表达,对纯化后的异源CtCel6进行酶学特性测定后发现,其对β-D-葡聚糖、磷酸溶胀纤维素(PASC)和羧甲基纤维素钠(CMC-Na)的催化活性分别提高了1.82、1.65和1.43倍,在80℃和90℃下的半衰期(t1/2)分别延长了1.42倍和2.40倍。上述研究表明,嗜热性木质纤维素酶在嗜温性宿主中的异源表达不仅可以在常规条件下提高酶的产量,还能很大程度上提高酶的活性和稳定性,而今后如何筛选和优化异源表达宿主及其培养条件,以及如何构建更加具有适应性的表达载体系统将会是科研人员的热点课题。
3.4.2 嗜热性木质纤维素酶在植物中的异源表达 既然木质纤维素的预处理过程是现阶段纤维素乙醇生产的主要经济负担,目前预处理的主要思路也是集中在酸解、碱解或酶解等外部方法上,并且这些方法进展也比较缓慢,那么从植物本身入手,通过基因工程技术在植物组织中异源表达编码木质纤维素降解酶的基因,从而从植物内部克服木质纤维素的顽固性,以此来实现预处理过程的经济化,便成为了一种绝妙的替代或辅助方法[33]。不过,嗜温性酶的植物内异源表达可能会导致与细胞内毒性有关的生理问题,如在正常的生长温度下,会对植物的生长和发育产生强烈的负面影响,另外,嗜温酶的性能参数与工业预处理条件(高温、极端pH值或有机溶质)也不匹配,应用效果不太理想,所以,研究人员便进行了嗜热性木质纤维素酶在植物中的异源表达尝试。Mir等[34]在拟南芥中表达了密码子优化的重组超嗜热内切葡聚糖酶(EG)和木聚糖酶(Xyn)基因,结果显示,植物生长或产量没有出现明显的损失,在转基因拟南芥的干燥茎中观察到最高的酶活性,表明该酶在茎衰老和贮藏期间未发生降解,同时,相对于野生型对照植物,来自转基因拟南芥的生物质显示出改善的糖化效率。Castiglia等[35]把编码嗜热菌的内切葡聚糖酶、内切β-1,4-木聚糖酶和β-葡萄糖苷酶的基因在烟草质体中表达,其蛋白得率范围为总可溶性蛋白(TSP)的2%-75%,其中,内切葡聚糖酶的积累(2%TSP)使质体发育受损,出现了幼苗致死表型,内切β-1,4-木聚糖酶和β-葡萄糖苷酶经热处理后的产量高达105.4 U/mg和255.4 U/mg,保留了天然嗜热酶的主要特征,对植物的生长没有不利影响。当在60℃的温度下,用质体衍生的木聚糖酶和β-葡萄糖苷酶替代商业酶进行巨型芦苇生物质水解时,显示出了比商业酶更高的水解速率以及木糖和葡萄糖产率。Xiao等[36]在转基因杂交杨中表达了来自Thermotoga neapolitana的嗜热内切葡聚糖酶(TnCelB),结果发现,在叶片和发育中的木质部具有较高的TnCelB积累,该部位的木质素含量和纤维素结晶度明显降低,对生物质收获并进行热处理后,糖化效率明显提高。由上可知,植物中异源表达嗜热性木质纤维素酶既可以作为一种获取嗜热性酶的来源,还可以从本质上改变植物木质纤维素的组成含量,减少发酵前添加外源酶的数量。在未来的研究中,通过对植物中这种异源表达机制的深入研究,如果能找到更加科学有效的方法来适度增加植物体内这些酶的积累以及减少因表达木质纤维素酶而造成的植物毒性,那么这种技术对纤维素乙醇的商业化进展将会产生巨大的促进作用。
3.4.3 嗜热性木质纤维素酶的代谢调控 嗜热性酶与常规嗜温酶一样,通常是以诱导酶的形式分泌,因此不管是同源表达还是异源表达,它们的产量甚至活性都受到激活和抑制机制的调节控制。如果能对这种调控机制深入研究并加以利用,嗜热性木质纤维素酶的产量必将大幅度提高,纤维素乙醇的商业价格也将会具有较大的竞争力。例如,研究发现在Trichoderma reesei中,含有5个编码木聚糖酶的基因和16个编码半纤维素酶的基因,这些基因的表达主要涉及到3个正调控因子(XYR1,ACE2,HAP2 / 3/5复合体)和两个负调控因子(ACE1,CRE1)[37]。Aghcheh等[38]研究 后发现,转录因子LAE1和VEL1可以控制纤维素酶的合成,而vel1和lae1的缺失则会导致纤维素酶表达的完全丧失。Zhang等[39]在Trichoderma reeseiRUT C30中引入了人工转录激活因子xyr1-cre1b和新型调控因子Trvib-1,重组后的Trichoderma reeseiVib-1在纯纤维素和其他可溶性诱导剂的存在下导致纤维素酶产量增加了两倍。Sun等[40]把Aspergillus nidulans中负责表达6-磷酸果糖氨基转移酶(glutamine-fructose-6-phosphate aminotransferase,GFAT)的 基 因gfaA删除,然后把来自Volvariella volvacea的调控因子gfat整合到载体pAL5和自主复制载体Prg3-AMA1-NotI中以分别构建gfat载体pALG和pAMAG,将两个表达载体转入到A. nidulans后,成功实现了嗜热性漆酶基因lcs的细胞内和细胞外过表达。Jain等[41]为了揭示Cu2+浓度对丝状真菌相关木质纤维素酶产量的影响,进行相关研究后发现,高浓度的铜离子能够引起微生物生物量的显著减少,但可以通过激活多种途径增加纤维素酶、木聚糖酶和漆酶的产量,并通过提高细胞壁的通透性来促进它们的胞外分泌。该研究虽然不是针对嗜热性酶,但是依据嗜热性微生物和嗜温性微生物代谢途径的相似性,该研究非常值得借鉴。另外,随着分子生物学和代谢工程技术的发展,有研究人员认为,通过引入调控元件、上调乙酸盐途径和ATP合酶亚基来激活嗜热性木质纤维素酶过表达,可以快速实现商业酶成本的下降[42-43]。
3.5 嗜热性木质纤维素酶协同作用
木质纤维素的复杂成分和结构使得单独一种酶无法实现其完全降解,必须通过多种酶的联合使用,才能将其快速高效的转化成可发酵糖。然而,由于自身底物的特异性,属于不同家族的酶可以显示出协同或反协同作用,通过开展深入研究,优化多酶混合物的最佳组合,实现不同酶之间的协同促进作用,对于提高底物转化率和产品收率具有重要意义。Aulitto等[44]以角豆树为底物对两种嗜热酶内切1,4-β-甘露聚糖酶(DturCelB)和α-半乳糖苷酶(TtGalA)进行比较协同研究,研究显示,当DturCelB和TtGalA以1∶1的组合进行测定时,没有表现出协同作用,以3∶1的组合进行测定时,表现出了一定的的协同作用,而以瓜尔豆骨架(DturCelB和TtGalA对瓜尔豆的单独比活度比角豆树低)为底物时,两者以1∶1和3∶1进行组合时,都表现出良好的协同促进作用。Zhou等[45]从嗜热性微生物Myceliophthora thermophila中得到了一种新型嗜热性溶解性多糖单加氧酶(MtLPMO9L),与纤维二糖水解酶(CBHI和CBHII)进行协同实验后发现,不同底物情况下MtLPMO9L对CBHI和CBHII表现出了不同的协同作用。当MtLPMO9L和CBHI的浓度比较低时(1∶10)时,两者对磷酸膨胀纤维素(PASC)表现出轻微的协同作用,而当两种酶的浓度比增加时(10∶1),对PASC和微晶纤维素(Avicel)具有明显的抑制作用;至于MtLPMO9L和CBHII则在不同浓度时对PASC和Avicel上均观察到协同作用,不过与Avicel相比,其对PASC的协同作用更为明显。除了类似上述嗜热性木质纤维素酶之间的协同作用外,有研究发现[46-47],辅助水解酶或辅助非水解酶、部分表面活性剂及牛血清白蛋白等对嗜热性木质纤维素酶也具有一定的协同促进作用。虽然关于嗜热性木质纤维素酶之间的协同研究文献数量有限,但通过上述研究可以确定,针对不同来源的木质纤维素底物,选择合适的嗜热酶及确定合适的比例进行联合使用,增强嗜热性酶之间的正协同作用,必能在一定程度上降低纤维素乙醇的生产成本。
4 结语
综上所述,纤维素乙醇作为化石能源极具潜力的替代品,其生产过程中存在的技术瓶颈严重限制了它的商业化步伐。利用嗜温酶预处理木质纤维素转化为可发酵糖的工艺中,嗜温酶的高成本和对工业化生产中极端环境的不适应性,成为了影响纤维素乙醇经济可行性的重要因素之一。嗜热性木质纤维素酶因其更高的热稳定性、更宽pH范围、更强的抑制剂耐受性、更低的杂菌污染风险、更高的反应速率以及更低的冷却成本,在纤维素乙醇生产过程中展现出了独特的优势。为了尽早实现嗜热性木质纤维素酶在纤维素乙醇中的商业化应用,科研人员分别从它的筛选、修饰、固定化、异源表达、代谢调控和协同作用等不同方向进行了大量研究,并在一定程度上实现了产量、活性和稳定性的提高。然而,在嗜热性木质纤维素酶的研究和应用过程中还存在许多困难,如固定化酶活性的下降、植物中异源表达时的细胞毒性等。但是,可以相信,随着研究的不断深入,这些困难都会逐一解决,纤维素乙醇的商业竞争力也会得到大幅度提高。
