施氮对干旱河谷岷江柏(Cupressus chengiana)幼苗光合生理特征的影响
2020-08-28冯秋红缪国辉徐峥静茹史作民李登峰
冯秋红,缪国辉,徐峥静茹,史作民,缪 宁,李登峰
(1.四川省林业科学院 四川卧龙森林生态系统定位站,四川 成都 610081;2. 森林和湿地生态恢复与保育四川重点实验室,四川 成都 610081;3. 理县林业和草原局,四川 理县 623102;4. 中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室,北京 100091;5. 四川大学生命科学学院生物资源与生态环境教育部重点实验室,四川 成都 610064;6. 四川农业大学,四川 成都 611103)
【研究意义】岷江柏(Cupressuschengiana)是柏科柏属乔木,为中国珍稀濒危种,分布于岷江、大渡河和白龙江流域的干旱河谷地带。因其具有良好的水土保持和水源涵养作用,成为长江上游高山峡谷地区干旱河谷地段水土保持的重要树种和荒山造林的先锋树种,一般生于海拔1200~2900 m的干燥阳坡[1-3]。近年来,由于人为干扰而遭到严重破坏,加之人为砍伐及气候影响,岷江柏生存环境日益恶劣,天然林日渐稀少。氮素是影响植物生长发育的重要营养因子。施氮肥可以促进植物苗高、地径的生长,增强植物生理代谢,增加苗木生物量,提高苗木质量和产量[4-6]。植物叶片中约75 %的氮素存在于叶绿体中[7],其中30 %~50 %被碳同化的关键酶——核酮糖-1,5-二磷酸羧化酶(Rubisco)所占据,还有一部分用于光捕获和电子传递等相关蛋白的合成[8]。因此,氮素与植物光合作用密切相关,研究施氮对岷江上游干旱河谷地区乡土树种岷江柏生长和光合的生理响应具有重要意义。【前人研究进展】岷江柏的相关研究多见于对其分布、生境和种群结构的研究[9-11],对岷江柏育苗、造林及恢复等技术方面也多有报道[12-13]。邵芳丽等[14]的研究表明,氮肥对岷江柏幼苗的净光合作用速率均有显著影响,且该影响随着水分梯度的变化而改变。亦有研究表明,在植物受到干旱胁迫时,施用氮肥将首先促进植物的抗旱性,如提高水分利用效率、增加氮素在根茎甚至根系的分配等[15-18]。【本研究切入点】目前,干旱区岷江柏人工林氮肥施用尚无统一标准,而且对干旱河谷地区岷江柏幼苗施氮后光合特性的研究鲜见报道。【拟解决的关键问题】因此本试验通过设置不同施氮量对岷江柏幼苗氮磷含量、氮磷比、气体交换及叶绿素荧光参数的影响,探讨岷江柏幼苗的养分、光合生理特征随施氮梯度的响应规律,为干旱河谷地区乡土树种——岷江柏种群的恢复与日常管护提供科学数据,并为干旱河谷地区植被恢复技术体系建立提供基础数据,为长江上游森林质量提升与生态屏障建设奠定理论基础。
1 材料与方法
1.1 试验地概况
试验地位于四川省阿坝藏族羌族自治州理县的薛城林场,东经103°21′50″,北纬31°34′38″,海拔1800 m。林场属山地型立体气候,春夏季降水量多,冬季无霜期短,年降雨量在650~1000 mm,年平均气温6.9~11 ℃,无霜期221 d。
1.2 供试材料
供试的岷江柏种子于2013年10-11月采自四川省小金地区。播前用0.5 %高锰酸钾溶液浸泡2 h,然后进行催芽处理。于2014年4月播种于阿坝州理县薛城林场的苗圃苗床中。营养土选用沙壤55 %、黄壤40 %和过磷酸钙5 %配制,土壤用1~2 mm的铁筛筛选,每立方米营养土用100 g硫酸亚铁和50 g六六六粉进行土壤消毒,防止猝倒病及地下虫。2015年5月,进行移苗工作。共移栽120株长势良好、生长情况相似、无病虫害的幼苗,一苗一盆,盆内浇透水。移苗后,全光环境下适应,期间死亡的幼苗,重新移栽补齐。
1.3 试验设计
试验于2015年7-10月在阿坝州薛城林场的大棚中进行。利用尿素[H2NCONH2(CO(NH2)2]进行人工施氮处理,试验采用单因素随机区组设计,分4个处理,施氮水平依次为:0.00 mg·cm-3(N0)、0.75 mg·cm-3(N1)、1.50 mg·cm-3(N2)、2.25 mg·cm-3(N3)。施肥在连续晴天、浇水后的第2天傍晚进行,施肥时在离幼苗6~8 cm处挖出一个深10 cm的环形沟,将肥料均匀施于环形沟内,覆土。之后,每周对苗木进行统一浇水1次。试验测定开始于施肥后2个月,测定周期为7 d,测定期间不再对植物进行浇水。
1.4 试验指标测定
1.4.1 气体交换参数 在施氮后2个月(9月),分别测量了岷江柏幼苗在不同施氮水平上的叶片气体交换参数值。在各梯度实验中,每个处理选取4株植株,每株选择向阳的完全展开的成熟鳞叶2~3片于上午9:00到下午3:00在野外大气环境条件下,控制叶室内CO2浓度为400 μmol·mol-1,温度设定为25 ℃、湿度控制在75 %±5 %、空气流速为0.5 L·s-1,采用Li-6400 (LI-COR,Lincoln,Nebraska,USA)通过改变光强进行光响应曲线的测量,获得不同处理类型植株的光饱和点,具体的光强梯度为:2000、1800、1600、1400、1200、1000、800、600、400、200、0 μmol·m-2·s-1。得到光饱和点后,在其他条件不变的情况下,将光强设为饱和光强,得到净光合速率(A)、蒸腾速率(T)、气孔导度(Gs)、水分利用效率(WUE)等指标。每个叶片重复读数10次以上。叶片叶面积的测定通过WINFOLIA叶面积测量系统(Regent Instruments Inc.,Quebec,Canada)完成。
1.4.2 叶绿素荧光参数 在天气晴好的上午,在野外大气环境条件下,使用Li-6400的荧光叶室进行测量。将植株在自然条件下光照活化30min,测定光下最小荧光(Fo′)、光下最大荧光(Fm′)和稳态荧光(Fs),并计算PSII实际光化学效率(ΦPSII)和光合电子传递速率(ETR)等参数。每个处理测定4个植株,每株植株测定2~3个叶片,每叶片记录3次,
结果取平均值。
ΦPSII=(Fm′-Fs)/Fm′
(1)
ETR=PAR×ΦPSII×0.84×0.5
(2)
1.5 数据处理
运用非直角双曲线修正模型结合SPSS 18.0软件的非线性回归,计算光响应曲线的相关参数和模拟光响应曲线[19]。
采用Excel2010进行数据的录入和前期处理,采用SPSS 18.0软件进行单因素方差分析(one-way ANOVA) 和Pearson相关分析。采用LSD法检验比较各处理间的差异显著性,采用Duncan法进行多重比较。
2 结果与分析
2.1 施氮对岷江柏幼苗氮、磷含量及氮磷比的影响
由表1可以看出,施氮后岷江柏幼苗叶片氮含量较N0有所增加,但不显著;施氮也增加了根茎氮含量,且在N1、N2水平上达到显著水平(P<0.05),最高可达14.227 mg·g-1(N2),较N0提高了5.62 %;不同于岷江柏幼苗氮含量的变化,随着外界供氮量的增加,其叶片磷含量呈逐步下降趋势,且施氮处理植株的叶片磷含量均显著低于N0。而在不同施氮处理间,N3磷含量最低,较N0降低了36.89 %,且显著低于N1,与N2间无显著差异;由表1可以看出,岷江柏叶片氮磷比随氮含量的增加而增加,且在各施氮水平上均显著(P<0.05)高于N0。
2.2 施氮对岷江柏幼苗气体交换参数的影响
由表2可以看出,岷江柏幼苗的A随着施氮量的增加,呈现先增加后降低的趋势,N1最高,且显著高于其他处理类型(P<0.05),而其他处理之间无显著差异;对于Gs而言,随着施氮量的增加,岷江柏幼苗的Gs在N0、N1和N2间变化不显著,在N3水平下呈现显著降低,且显著低于其他施氮水平(P<0.05)。
随着施氮量的增加,岷江柏幼苗的T先升高再降低,其中N1最高,显著高于N0,N3最低,且显著低于其他处理类型,N2与N0、N1之间无显著差异;同时,WUE随着施氮量的增加而呈现波动趋势,其中N3显著高于其他处理,N1显著高于N0和N2,而后两者间无显著差异。
2.3 施氮对岷江柏幼苗叶绿素荧光参数的影响
ΦPSII为实际光化学量子产量,反映了PSII反应中心在部分关闭情况下实际原初光能捕获效率,反映叶片用于光合电子传递的能量占所吸收光能的比例,即实际光能利用[20-21];ETR为表观光合电子传递效率,它与植物的光合速率有很强的线性关系,是一个表征植物光合能力高低的变量[22]。由表3可见,随着施氮量的增加,岷江柏幼苗叶片的ΦPSII、ETR呈现逐渐升高的趋势,且除N1外,其他处理均达到显著水平(P<0.05),ΦPSII、ETR在N3水平分别达到0.106和92.752,较之N0增加了16.483 %、16.662 %。说明随着施氮量的增加,叶片PSⅡ光能捕获效率逐渐升高,光能利用率升高。

表3 不同施氮水平岷江柏幼苗叶绿素荧光参数
2.4 施氮水平岷江柏幼苗叶片主要生理指标相关分析
通过对不同施氮水平下岷江柏叶片主要生理指标进行相关分析(表4),表明岷江柏幼苗的磷含量、Gs、T与施氮水平呈极显著负相关关系(P<0.01),而叶片氮磷比、水分利用效率、ΦPSII、ETR与施氮水平呈极显著正相关关系(P<0.01)。叶片磷含量与WUE呈显著极负相关关系(P<0.01),而与ΦPSII呈显著负相关关系(P<0.05);叶片氮磷比与WUE呈显著极正相关关系(P<0.01),而与ΦPSII、ETR呈显著正相关关系(P<0.05);A与Gs、T呈现极显著的正相关关系(P<0.01),此外,Gs与T呈极显著正相关关系(P<0.01),与ΦPSII、ETR呈现显著负相关关系(P<0.05);WUE与ΦPSII呈极显著正相关关系(P<0.01),与ETR呈现显著正相关关系(P<0.05);同时,T与ΦPSII、ETR均呈现显著负相关关系(P<0.05)。最后,ΦPSII与ETR呈现极显著正相关关系(P<0.01)。
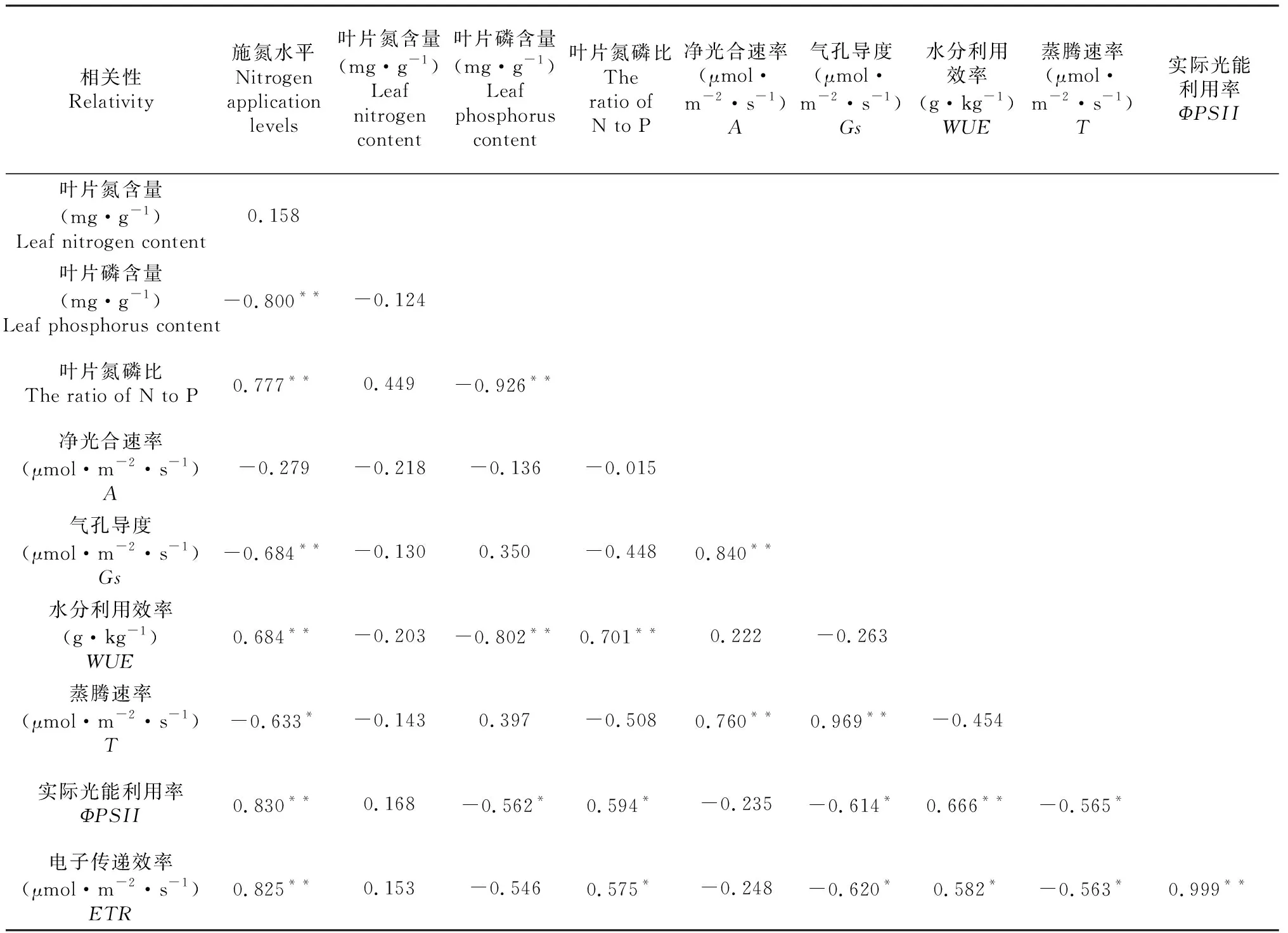
表4 不同施氮水平岷江柏幼苗生理指标相关分析
3 讨 论
氮是植物体最重要的结构元素之一,是蛋白质的基本构成单元,参与了生物结构的构成和生命活动,对植物机体十分重要。氮素在植物体的各个器官之间的分配和循环对于植物的生长和生理活动也具有重要的影响。在本研究中,施氮处理并没有显著提高岷江柏幼苗叶片的氮素含量,但施氮却显著提升了根茎的单位质量氮素含量(其中N1和N2水平达到显著水平),可见,相较于叶片,岷江柏的根茎氮含量对施氮具有更显著的正向响应,这可能表明试验植株仍处于干旱胁迫状态。前人研究表明,处于干旱胁迫下的植物,会将更多的养分物质用来缓解干旱,即促进根系生长,进而获取更多的水分利用同位素标记的方法对干旱胁迫下胡杨幼苗的相关研究也得到了相似的结论[15,17]。同时,施氮处理还显著降低了叶片磷含量[23]、进而显著提升了叶片氮磷比。岷江柏叶片氮磷比的变化范围为8.402~13.127,均小于14,说明本研究中岷江柏幼苗生长主要受N素限制[24-25],但随着施氮量的增加,该限制逐渐得以缓解。
氮是氨基酸和核酸的必要组成成分之一,植物叶片的氮素绝大部分都存在叶绿体中,而叶绿体又是光合作用的主要场所,因此,氮素含量的变化在一定程度上影响着植物的光合作用[26]。叶绿素荧光动力学参数包含着光合作用过程的重要信息,在探测逆境对光合作用影响方面具有独特的作用,能够反映光合系统“内在性”特点[27]。前人研究[14]表明,植物的叶片氮含量、光合效率与施氮量呈正线性相关,但也有研究[28-29]表明植物的叶氮含量、光合能力与施氮量之间并不呈线性关系,即达到一定施氮量后,植株的光合能力和生长可能会对施氮响应不显著甚至受到抑制。本研究中,在施氮N1水平下,虽然岷江柏叶片氮含量无显著增加,且叶片磷素含量显著降低,但气孔导度仍有所提升,蒸腾速率显著提升,进而光合速率却显著增加,这可能与随着施氮量增加而显著改善的叶绿素荧光参数有关,即,岷江柏幼苗叶片的ΦPSII和ETR均随着施氮量的增加而显著增加,表明施氮能促进PSII反应中心的开放程度、实际光能捕获效率和电子传递速率[30],进而促进光合速率[19,29-30],此阶段为非气孔限制;随着施氮量的持续增加(N2、N3水平),叶片氮氮含量仍没有显著增加,且磷素含量持续降低,虽然光合荧光参数持续增加,但气孔导度逐步降低,导致光合速率却开始降低,水分利用效率升高(N3水平),植物进入气孔限制阶段。这可能与植物启动干旱胁迫下抗旱反应有关,相关研究表明,干旱胁迫下适量施氮有利于提升植物抗旱酶指标,进而提升其主动抗旱能力[18],从而促使植物降低气孔导度和蒸腾速率[31-33,29-30],提升水分利用效率[34-35]。毕竟,在干旱地区,与生长相比,干旱胁迫下的节水抗旱显然占据了更加重要的位置[23],所以在施氮量持续增加时(N2和N3水平),虽然岷江柏幼苗叶片的ΦPSII和ETR持续升高,但光合速率却显著降低。
4 结 论
综上所述,在干旱河谷地区,施氮没有提升岷江柏叶片氮含量,但却促进了根茎氮含量,同时抑制了叶片磷的吸收,氮磷比显著增加。本研究中岷江柏幼苗的氮磷比小于14,主要受N限制,适量的施氮可以缓解该趋势;与此同时,施氮显著促进了岷江柏的光合系统的PSII反应中心的开发程度和实际光能捕获效率,但净光合速率却随着施氮量的增加先升高再降低,这可能与植物随着施氮量而逐步提升的主动抗旱节水应急有关,即,当光合速率持续增加而带来更大的水分消耗时,岷江柏通过降低气孔开放程度(气孔导度降低),进而控制机体水分丧失(蒸腾速率降低),从而达到提升水分利用效率,提升抗旱能力的目的。可见,对干旱地区岷江柏而言,少量施氮可以促进其生长,随着施氮量的增加,植物抗旱能力提升,且以降低光合生长为代价。
