滆湖藻密度的时空分布及驱动因子分析
2020-08-26沈丽娟周崴沈伟
沈丽娟 周崴 沈伟
(1. 江苏省常州环境监测中心,江苏常州 213001;2. 江苏省环境保护水环境生物监测重点实验室,江苏常州 213001)
1 引言
滆湖位于江苏省常州市西南,东南通太湖,西连长荡湖,南接宜兴市,是苏南地区仅次于太湖的第二大湖泊,面积146 km2,是苏南太湖湖群的重要组成部分,具有饮用、灌溉、航运、游览和水产增养殖等多种功能[1]。湖区地形多为低洼平原,沿岸水系发达,水网交错,来水主要是西北部的地表径流及工业废水和生活污水排放,近年来随着污染负荷和富营养化程度不断加大,滆湖在夏季频繁暴发蓝藻水华,破坏了健康平衡的水生生态系统,降低了水资源利用效能,影响滆湖作为常州市备用饮用水水源地的功能。
浮游植物作为湖泊水体中主要的初级生产者,是食物链的基础环节,易受环境中各种因素的影响而在较短的时间内发生变化,相对于其他生物类群更能及时反映水域生态系统的情况[2]。藻密度(Algal density)是单位体积内藻细胞的数量,是最能直接说明水体中藻类数量和水华暴发强度的指标。虽然叶绿素a 也可间接反映藻类浓度,但是由于其容易受其他色素(如大型水生植物)或色素衍生物的干扰,在测定过程中又容易受到样品保存时间、检测环境的影响[3]而造成评价结果的不可靠,所以本研究采用藻密度而非叶绿素a 作为研究指标。
对于藻密度与环境因子关系的研究方法,需根据不同研究目的而定。若旨在揭示不同藻类的藻密度分布与主要环境因子之间的对应关系,可以采用典范对应分析(CCA)[4];研究藻密度与水质的简单关系可以采用相关分析[5]。研究多种变量对藻密度分布情况的影响,可以考虑多元线性逐步回归法和偏相关分析,前者是在因变量(藻密度)与自变量(水质因子、气象因子)之间建立多元回归方程,通过逐步回归法确定主要影响因子;后者则通过固定因变量与其中1 个自变量以外的其他变量对它们的影响后,研究这2 个变量之间的相关关系,它反映了事物间的本质联系[6]。
2 材料与方法
2.1 样品采集与检测
在滆湖内共设置20 个采样点,在2 年内每季度采样1 次,采集水面下0.5 m 处水样。其中S38 高速公路以北的北部湖区设置6 个采样点,中部湖区设置10 个采样点,属于宜兴市管辖的南部湖区设置4个采样点。采样点设置具体见图1。
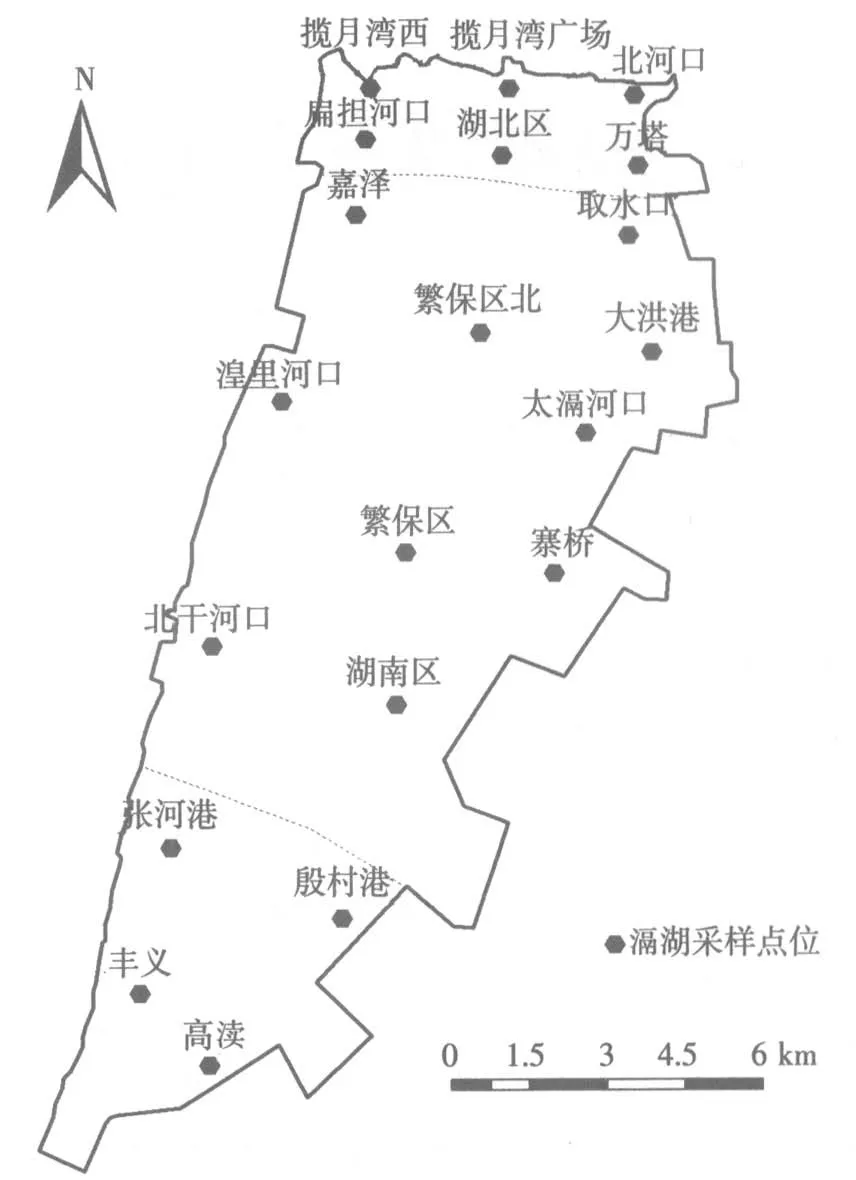
图1 滆湖分区及采样点分布示意
水样分析指标主要有pH、溶解氧(DO)、水温(T)、透明度(SD)、总氮(TN)、硝酸盐氮(NO3-N)、氨氮(NH3-N)、总磷(TP)、磷酸盐(PO43-)、高锰酸盐指数(CODMn)。pH,DO,T 采用YSI 6600 型多参数水质监测仪现场测定,SD 采用塞氏盘法测定,TN,NO3-N,NH3-N,TP,PO43-和CODMn参照《水和废水监测分析方法》[7]测定。浮游植物采样及鉴定方法:在水下0.5 m 处取1 L 水样,立刻加入15 mL 鲁哥试剂固定浮游植物,带回实验室,静置48 h 后浓缩至30 mL,在显微镜下进行浮游植物种类和藻密度的计数。
2.2 统计分析方法
数据统计分析和制图使用Excel 2013,SPSS 21.0 和ArcGIS 10.3 软件。
3 结果与分析
3.1 滆湖藻密度时空分布特征
滆湖全湖及各湖区藻密度均值及变幅见表1。由表1 可见,2013—2014 年期间,滆湖藻密度平均值为(3 506±5 350)×104cells/L,变幅为62×104~39 491×104cells/L。

表1 滆湖全湖及各湖区藻密度均值及变幅 ×104 cells/L
运用ArcGIS10.3 软件插值分析模块中的克里金法绘制滆湖2013 年和2014 年全年藻密度平均值的空间分布图(图2)。2013 年和2014 年滆湖藻密度空间分布趋势大致相同,均为由北向南藻密度逐渐升高,这与陈立婧[8]2004 年4 月—2006 年1 月在滆湖所做研究的藻密度空间分布趋势完全相反。K-W 检验显示北部湖区与中部湖区和南部湖区之间藻密度差异极显著(P<0.01),中部湖区与南部湖区之间藻密度差异接近显著(P=0.056)。2013 年和2014 年藻密度最高值均出现在南部湖区,有所不同的是,2013年南部西岸丰义区域藻密度最高,2014 年南部东岸殷村港区域藻密度最高。

图2 滆湖藻密度年均值空间分布
滆湖各湖区2013—2014 年藻密度的时间分布状况见图3。冬季各湖区的藻密度普遍较低,且差距不大。春季藻密度开始快速上升,南部湖区藻密度的增长要明显快于中部湖区和北部湖区,2013 年夏季藻密度达到高点后回落,而2014 年夏秋季节藻密度均低于春季。Wilcoxon 秩和检验显示,年际间季节差异均极显著(P<0.01),其中冬、春季节2014 年藻密度显著高于2013 年,夏、秋季节则相反;2013 年年内,春季和秋季之间藻密度差异不显著(P=0.09),其他季节互相之间藻密度差异均显著(P<0.05);2014年年内,秋季和冬季、夏季之间藻密度差异不显著(P>0.05),其他季节互相之间藻密度差异均极显著(P<0.01)。
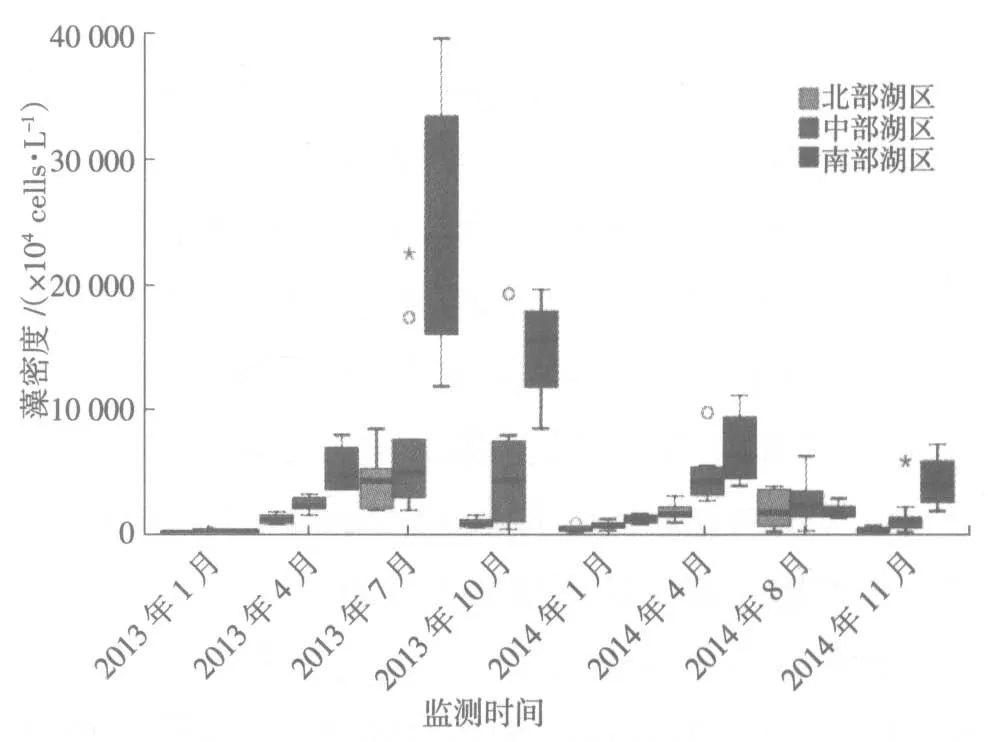
图3 滆湖藻密度时间分布
通过对滆湖浮游植物组成的分析发现,滆湖浮游植物的演替规律大致为:冬、春季节以绿藻和硅藻为主,夏、秋季节蓝藻占绝对优势,季节性差异明显。滆湖浮游植物种类繁多,蓝藻门、绿藻门、硅藻门在全年均占有较大比重,蓝藻门、绿藻门、硅藻门和黄藻门的最高值出现在南部湖区,其他门类的藻类最高值出现在中部湖区,夏秋季节蓝藻门密度占浮游植物总密度比例均在70%以上,见图4。

图4 滆湖藻密度物种组成
3.2 多元线性逐步回归法确定驱动因子
多元线性逐步回归确定驱动因子的方法已广泛应用于环境预测、环境监测、环境评价等多个环境领域[9]。由于多元线性逐步回归分析要求数据呈正态分布,而滆湖藻密度数据呈明显的正偏态分布,对藻密度数据进行对数变换后,通过正态性检验。由于ρ(PO43-)与ρ(TP)的相关系数高达0.886,ρ(NH3-N)和ρ(NO3-N)与ρ(TN)的相关系数也分别达到了0.769 和0.773,为消除因子间的多重共线性,不能将以上环境因子全部列入备选驱动因子。
由于分析方法的原因,TN 和TP 的测定过程中将水样中悬浮态的蓝藻、泥沙所含的氮、磷均包含在内[10],而NH3-N,NO3-N 测定中的预处理过程少了蓝藻、泥沙的干扰,更能真实反映水体营养盐含量,通常浮游植物也倾向吸收水体中的NH3-N 和NO3-N来合成细胞所需要的氨基酸等物质[11]。
藻类作为水生生态系统中的生产者,通过光合作用吸收CO2,生成O2。天然水体中的pH 值主要受CO2含量的控制,在藻类生长旺盛的水域,为进行光合作用藻类需从水体中吸收大量的CO2,使得水体酸度降低,pH 值升高,并释放O2[11]。因此在富营养化水体中,O2和CO2主要受生物过程的控制[12],藻类数量的多少、生命活动的旺盛程度必然会影响水体的pH 值变化。因此水中DO 和pH 的变化是藻类生命活动的反映,难以用于预测藻类生长情况,不能作为驱动因子。
基于以上原因,设置因变量Y 为log10(藻密度),自变量个数m 为6,自变量X1为T(水温),X2为SD(透明度),X3为ρ(NH3-N),X4为ρ(NO3-N),X5为ρ(PO43-),X6为ρ(CODMn)。
运用SPSS 软件对数据进行回归分析,回归方程模型见表2。

表2 藻密度与环境因子的回归方程
多元线性逐步回归模型见表3。

表3 多元线性逐步回归模型结果
由表2 可知,在多元线性逐步回归分析中,全湖和所有湖区回归方程中均选入了环境因子X1,除北部湖区以外的回归方程都选入了环境因子X6,北部湖区和中部湖区的回归方程还分别选入了环境因子X4和X3。
回归方程模型拟合度检验表明,多重相关系数和多重判定系数说明这些回归方程均有较好的拟合度,北部湖区、南部湖区和全湖分别有69.4%,79.7%和65.1%的藻密度可以通过2 个环境因子用模型解释,中部湖区有64.1%的藻密度可以通过3 个环境因子用模型解释。
环境因子驱动力大小的比较不能直接采用回归系数,而要采用标准化系数,标准化系数的绝对值越大说明驱动因子的驱动力越大,全湖和南部湖区藻密度的环境因子驱动力强弱顺序为:X1(T)>X6(CODMn),中部湖区藻密度的环境因子驱动力强弱顺序为:X1(T)>X6(CODMn)>X3(NH3-N),北部湖区藻密度的环境因子驱动力强弱顺序为:X1(T)>X4(NO3-N)。
3.3 偏相关系数分析
简单的高度相关并不意味着因果性[13],运用简单相关计算2 个变量的相关关系时,常常因为第3个变量的影响,使简单相关系数不能真正反映2 个变量间的线性关系的程度和方向[14],而偏相关系数是在排除其他变量影响后计算得到的,具有显著性统计意义的偏相关系数能反映事物间的本质联系,并已成功应用于筛选驱动因子的研究中[6]。
基于与多元线性逐步回归分析同样的原因,在偏相关分析中以log10(藻密度)作为因变量Y,运用SPSS 软件对数据进行偏相关性分析。结果表明,在全湖和各个湖区分别有2 个环境因子与Y 都有极显著的相关性关系(P≤0.01),其中北部湖区为T 和NO3-N,其他湖区和全湖均为T 和CODMn,见表4。

表4 藻密度与环境因子的偏相关性分析
4 讨论
4.1 驱动因子及驱动力大小
通过以上2 种统计分析方法,除中部湖区外,均确定了相同的驱动因子,全湖和南部湖区为T 和CODMn,北部湖区为T 和NO3-N。在中部湖区,多元线性逐步回归法确定的驱动因子有3 个:T,CODMn和NH3-N,而偏相关系数法确定的仅为T 和CODMn。
对于驱动力大小的判定,多元线性逐步回归法由于采用标准化系数的方法,不仅与自变量的回归系数有关,而且与这个自变量的波动程度有关,因此可能会有偏差,将样本数据中波动程度较大的自变量赋予过高的驱动作用力[15]。而偏相关系数法对于驱动力大小的判定则不会受自变量样本波动性的影响。在中部湖区,NH3-N 的变异系数CV 高达106%,其他环境因子的变异系数CV 则均在56%以下,因此比较2 种方法的结果可以确定NH3-N 由于数据波动程度较大,而被误选入回归方程,中部湖区藻密度的驱动因子顺序确定为T 和CODMn。
从驱动力大小的顺序来看,中部湖区和南部湖区2 种统计方法的结果一致,为T>CODMn,而北部湖区和全湖的2 种统计方法计算的驱动力大小顺序相反,但实际的系数差距不大,可以视为驱动力相近。该结果说明对于滆湖藻密度影响最大的驱动因子是T 和CODMn,对藻密度起到决定性的作用。
浮游植物的动态演替规律是多个环境因子在时间和空间序列上作用的结果[16],不同的湖泊具有不同的主要影响因子。温度是各种浮游生物时空分布的重要决定因素,不同的浮游植物都有其生长、繁殖的最适温度范围。研究表明,水温对浮游植物的生长、繁殖具有显著的作用[17-18]。不同季节温度影响了藻类的光和作用、呼吸作用和生长速率,温度升高是蓝藻形成优势的重要原因之一,特别是夏季,蓝藻更适应高温环境[19],蓝藻在19.5 ℃以后占优势[20]。CODMn反映了水体有机污染情况,对苏州平原河网区浅水湖泊的研究显示,CODMn对浮游植物的分布具有一定的影响,绿藻能耐受较高的CODMn,有机污染严重的湖泊中绿藻的丰度比较大[21]。滆湖蓝藻密度和绿藻密度分别占藻密度的49%和26%,夏季更是高达75%和17%,体现了T 和CODMn驱动下蓝绿藻的优势地位。
4.2 未入选的环境因子
营养盐对藻类生长的影响作用一直受到湖泊学研究者的关注,2 种统计分析方法均显示NH3-N 和PO43-不是藻密度的驱动因子,NO3-N 仅在相对独立的北部湖区是藻密度的主要驱动力。随着湖泊富营养化,特别是P 浓度的增加,通常会导致水体中蓝藻在浮游植物群落演替中占优势[22]。温带湖泊TP 浓度在0~30 μg/L 时,蓝藻占优的几率为10%以下,30~70 μg/L 时为40%,接近100 μg/L 时,风险达到80%左右[23]。陈宇炜等在太湖梅梁湾研究显示,氮、磷与藻类叶绿素a 和总生物量以及各门类生物量均无显著的线性相关关系,说明N,P 浓度的变化对梅梁湾藻类的生长没有直接的影响[24]。由于近年来营养盐的过量输入,滆湖N,P 含量已处于很高的浓度(总氮均值2.99 mg/L,总磷均值0.10 mg/L),已超过浮游植物的同化能力,不再是藻类生长的限制因子。
同时SD 也没有入选驱动因子,这是由于浅水湖泊有着与深水湖泊完全不同的一些特点。滆湖与太湖均属典型的浅水湖泊,滆湖平均水深相比太湖而言更小,与深水湖泊相比,浅水湖泊一般水底较平、水浅,底泥更频繁地受到风浪等水动力作用的扰动,SD 水平普遍较低,且波动幅度不大,在各湖区间差距不大。
4.3 其他可能影响藻密度的驱动因子
各湖区及全湖多元线性回归方程对藻密度的解释度在64%~80%之间,说明还有其他驱动因子对藻密度影响较大。
4.3.1 水动力条件及气象因素
水文条件影响到蓝藻的生长和分布,蓝藻的生长需要稳定的水体[20]。中部湖区回归方程解释度最低,这可能是由于滆湖是典型的吞吐型湖泊,主要的出入湖河流集中在中部湖区,水力滞留时间较短,流速较快,水动力条件较为复杂,单纯使用统计方法无法很好地模拟蓝藻生物量的分布和变化,而南部湖区出入湖河流较少,流速较低,有利于浮游植物生长。表层水体中藻类密度更多地受到风浪、湖流等外部条件的影响,而多元逐步回归无法模拟风浪、湖流等动力效应[3]。在夏季微囊藻为优势种的时期,具有伪空泡的微囊藻群体,其受风浪的影响漂移并在下风处聚集的特性在逐步回归统计中同样无法模拟计算。
4.3.2 鱼类及浮游动物摄食
由于藻类是湖泊的初级生产者,其他营养级的生物摄食藻类也是影响藻密度和藻类群落结构的重要因素。滆湖是重要的渔业水域,围网养殖面积较大,滆湖南部进行的围网实验显示,围网内浮游植物度显著低于围网外[25]。一般说来,蓝藻、绿藻、裸藻被视为鲤科鱼类难于消化的种类;硅藻、金藻、甲藻、隐藻为易于消化的种类[8]。孙育平[26]进行的围隔实验表明,鲢使浮游植物群落以甲藻—硅藻—绿藻型向绿藻—硅藻—蓝藻型模式演替,优势种向可食性差或具快速生长能力的种类演替。浮游动物的选择性摄食对浮游生态系统各种群的共存以及藻类的快速进化有着重要的影响。浮游动物摄食的选择性包括2 个方面:有毒藻类产生的毒素能抑制浮游动物的摄食,使浮游动物能避开摄食有毒藻类;同时,由于浮游动物偏好摄食无毒藻类,也减少了对有毒藻类的摄食。周健等[27]的研究表明,太湖夏季后生浮游动物摄食并不能控制太湖蓝藻水华,相反,后生浮游动物特别是大型浮游动物能促进蓝藻水华的形成。同时表明,后生浮游动物群落结构可能是影响微囊藻水华形成的重要因素。
4.3.3 高等水生植物竞争
2010 年后滆湖北部地区开展了大规模的整治工程,包括前置库入湖污染负荷控制[28]、底泥疏浚、大型仿生式机械捞藻和水生植被重建,由于芦苇、菱角等高等水生植物的竞争,相同月份水生植被重建区浮游植物生物量要比敞水区低63.9%~91.1%[29],这可能是北部湖区藻密度在全湖最低的重要原因。
5 结论
(1)滆湖全湖藻密度平均值为(3 506±5 350)×104cells/L,,变幅为62×104~39 491×104cells/L。
(2)滆湖藻密度空间分布特征为由北向南逐渐升高,北部湖区与中、南部湖区之间藻密度差异极显著。
(3)滆湖藻密度时间分布特征为冬季各湖区的藻密度普遍较低,且差距不大。春季藻密度开始快速上升,南部湖区藻密度的增长要明显快于中部湖区和北部湖区,2013 年夏季藻密度达到高点后回落,而2014 年夏秋季节藻密度均低于春季。浮游植物的演替规律为:冬、春季节以绿藻和硅藻为主,夏、秋季节蓝藻占绝对优势,季节性差异明显。
(4)滆湖全湖及中、南部湖区藻密度的驱动因子是T 和高CODMn,北部湖区藻密度的驱动因子是T和NO3-N,所有驱动因子对滆湖藻密度的影响均为正效应。其他可能的驱动因子包括水动力条件及气象因素、鱼类及浮游动物摄食和高等水生植物竞争。在富营养化严重的滆湖,N,P 营养盐已经不再是浮游植物生长的主要驱动因子。
