不同返青休牧期土壤微生物碳代谢特征及其影响因素研究
2020-08-20王晓丽温军马玉寿王彦龙施建军周华坤
王晓丽,温军,马玉寿,王彦龙,施建军,周华坤
1. 青海省畜牧兽医科学院/青海大学/省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016;2. 流域水循环与生态省级重点实验室/青海省水资源高效利用工程技术研究中心/青海省水利水电科学研究院有限公司,青海 西宁 810001;3 青海省寒区恢复生态学重点实验室/中国科学院西北高原生物研究所,青海 西宁 810008
三江源区高寒草甸生态环境脆弱,对自然环境和人类生产活动的干扰较为敏感(杜玉娥等,2011)。研究表明三江源区高寒草甸持续退化(赵新全等,2005;Fan et al.,2010;徐翠等,2013),高原地区过度放牧是草地退化的直接原因(曹广民等,2007;樊江文等,2011;韦惠兰等,2016)。面对草地退化这一严峻形势,科学制定放牧制度,合理利用高寒草甸牧草资源,才能达到保护天然牧场和生态环境的目的。
青藏高原地区,研究人员通常将放牧草场分为暖季草场和冷季草场进行研究(董全民等,2012)。由于牧草在春夏和秋冬之交生长缓慢,再生性能差,此时最易发生过牧,牧草过高的利用率严重影响了牧草的再生,引起草地的严重退化(胡金梅等,2008)。春季作为关键忌牧期,是牧草贮藏营养物质和生长的关键时期,对放牧的时间、强度十分敏感。基于植物生长发育节律在草地放牧敏感期舍饲休牧才是“时间机制”的基本内涵(李青丰等,2005)。
春季休牧(返青期休牧)在内蒙古典型草原被证明可以有效保护草地生态环境,平衡畜牧业发展和草地生态环境之间的关系,有利于草地植被的可持续利用(李青丰,2005;赵钢等,2006;朱立博等,2008)。研究不同休牧时间对草地的影响,对休牧的可行性进行分析,休牧期间采用低投入维持性饲养,显著降低饲养成本,可提高草地生态效益、畜牧业生产实践可行性和经济合理性(李青丰等,2005)。学者们从种群分布格局、种间关系和碳储量等方面进一步研究了春季休牧对草原植被和土壤的影响。研究表明,植物群落和土壤有机碳储量随着休牧年限的延长而增多,但在春季休牧时资源限制了群落内植物种群的生长,种群因占有资源能力不同而产生竞争作用。在休牧后有放牧干扰时,家畜牧食行为成为限制植物生长的关键因素,资源的限制作用降低(李玉洁等,2013a;卫智军等,2016;吕世杰等,2016)。三江源区高寒草甸返青期休牧的研究工作尚处于初级阶段,且该区域高寒草甸生态系统独特、原始和脆弱,亟待在调控载畜量的基础上根据区域物候期和牧草返青期科学合理地制定春季休牧制度。土壤微生物作为生态系统重要的分解者,在物质循环和能量转化中占有重要地位(Somova et al.,2001;Schimel et al.,2012)。与土壤理化性质相比,土壤微生物对外界环境的变化更为敏感,常被用作表征土壤质量的灵敏性指标(Nielsen et al.,2002)。尽管基于生物标志物的测定方法(如磷脂脂肪酸PLFAs法)和分子生物学方法(DGGE等)可以反映微生物的群落结构信息,但却无法获得有关微生物群落总体活性与代谢功能的信息,Biolog方法弥补了这一不足(Garland et al.,1991)。
因此,本文拟利用Biolog方法研究三江源区高寒草甸生态系统不同返青期休牧时间下土壤微生物生物生物量碳氮及其碳代谢特征,不仅可以了解高寒草甸生态系统在不同返青休牧期下微生物的变化特征,而且通过监测土壤理化性质试图揭示微生物碳代谢对返青期休牧过程的响应机理。本研究可为三江源区高寒草甸的合理利用提供有效措施,为草地畜牧业的可持续发展提供理论依据,对优化放牧制度、揭示生态系统退化原因以及提升生态系统功能具有重要的科学意义。
1 试验区域与研究方法
1.1 试验地概况
试验地位于三江源国家自然保护区青海省果洛州玛沁县大武镇永宝村,地理位置为34°24′14″N,100°23′31″E,海拔 3920 m 左右。该区域属于典型高原大陆型气候,寒冷、干旱、多风,太阳辐射强,日照时间短,昼夜温差大。年均温为−3.9 ℃,最冷月平均气温为−12.6 ℃,最热月平均气温为9.7 ℃;年降水量513.2—542.9 mm,多集中在6—9月,年蒸发量2471.6 mm。牧草生长季约156 d,全年无绝对无霜期.草地为不同退化程度的高寒矮嵩草(Kobresia humili)草甸,主要草本植物为莎草科、蓼科、菊科和蔷薇科等。土壤为高山草甸土。
1.2 试验设计及测试方法
2017年5月10日三江源区果洛地区的返青期开始,选择作为冬春草场地势平坦的高寒草甸样地该草地原属于永宝牧场,近30年来,一直作为冬春草场使用,在夏秋季节是休牧的。如表1所述,分别设置CK(不休牧)、休牧20 d、休牧30 d、休牧40 d和休牧50 d,共5个处理。所有处理的休牧开始时间不一致,但结束时间统一为转场到夏季牧场的时间6月30日,其中休牧50 d的时间为5月10日—6月30日,休牧40 d的时间为5月20日—6月30日,休牧30 d的时间为5月30日—6月30日,休牧20 d的时间为6月10日—6月30日。对照 CK为不休牧,通过控制牧草利用率在 30%—50%来控制,放牧强度约为 0.89—1.45 cattle·hm−2,属中轻度放牧。每个处理设置3个重复样地。

表1 试验中各处理的休牧和放牧时间Table1 The times of no-grazing and grazing in the treatments
2017年8月在每个样地选择5个样方采集表层0—15 cm的土壤样品,每个样方的土样由3.5 cm直径的土钻采集5钻的土混合而成。在采样过程中为避免牲畜排泄物导致的对土壤特性的影响,绝对避开牲畜排泄物的,同时避开牲畜排泄物留存痕迹。同时,用便携式土壤三参数仪测定土壤的温度。土壤样品带回实验室后,过2 mm的土筛,捡出石粒、植物根系及枯落物等,并分成两份,一份在室内自然风干后,用于测定土壤理化性质等指标;一份在4 ℃冷藏保存,用于测定土壤微生物生物量碳氮和土壤微生物碳源利用特征及其多样性等指标。土壤有机碳(SOC)采用重铬酸钾氧化法测定,全氮(TN)和速效氮(NH4+-N和NO3−-N)采用全自动间断化学分析仪(CleverChem)测定,全磷(TP)采用钼锑抗比色法测定,速效磷(AP)采用碳酸氢钠浸提-钼锑抗比色法测定,全钾(TK)采用火焰光度法测定(杨剑虹等,2008),土壤微生物生物量碳氮采用氯仿熏蒸提取法测定(Brookes et al.,1985)。
土壤微生物碳源利用特征及其多样性分析采用 Biolog-Eco法测定(Choi et al.,1999;Miki et al.,2018)。采用 Biolog-Eco平板法测定的数据通过计算平均颜色变化率(AWCD)、McIntosh指数(U)、Shannon物种多样性指数(H)和Simpson优势度指数(D)来衡量土壤微生物群落碳代谢能力。其中每孔平均颜色变化率(AWCD)反映了土壤微生物群落碳源代谢能力的高低,是土壤微生物活性和多样性大小的一个重要指标。一般认为变化幅度较大的样品具有较高的碳源利用能力,也往往具有较高的微生物丰度。对第10天的土壤微生物31种碳源的碳代谢指标 AWCD进行了多样性指数的分析,包括微生物群落Shannon多样性指数、Simpson优势度指数和 McIntosh指数代表微生物群落功能多样性(Garland et al.,1991;Garland,1997)。
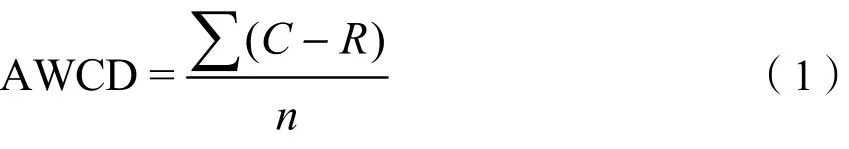
式中,C为31孔每孔的吸光值;R为对照孔吸光值;n为碳源种类数31。
McIntosh指数(U)来衡量群落均一性程度;

式中ni为31种碳源的平均值。
采用Shannon物种多样性指数(H)和Simpson优势度指数(D)反映群落中最常见的物种,评估微生物群落优势度;

式中,Pi为每个孔吸光度/所有吸光度之和。
1.3 数据处理及统计分析
用SPSS 22.0和Excel 2013对数据进行统计分析,其中不同休牧期间的差异比较采用单因素方差分析(One-way ANOVA),采用 LSD法或 Tamhane’s进行多重比较。利用R软件的ggplot 2包做了培养第 10天微生物利用 31中碳源的气泡图。利用canoco for window 4.5对拐点时间第10天的土壤微生物碳代谢指标 AWCD做主成分分析(principal component analysis,PCA),并且将31中碳源分成六类碳源,即糖类、氨基酸类、酯类、醇类、胺类和羧酸类。然后用第 10天的土壤微生物碳代谢指标 AWCD与土壤理化性质做冗余度分析(redundancy analysis,RDA),同时进行蒙特卡洛(Monte Carlo)检验选出在0.05水平上显著的环境变量。
2 结果与分析
2.1 不同返青休牧期下土壤碳氮及微生物生物量碳氮特征
不同返青期休牧下土壤有机碳(SOC)差异显著(P=0.000),且休牧 20 d(43.57 g·kg−1)显著高于其他处理,休牧50 d反而最低(22.83 g·kg−1)。土壤全氮(TN)在不同处理间差异显著(P=0.003),但与SOC的规律不同,休牧20 d、40 d和CK显著高于休牧30 d和50 d。土壤碳氮比(SOC:TN)在不同返青期休牧下差异也显著(P=0.005),休牧20 d和30 d显著高于对照和休牧40 d和50 d,不同处理间SOC:TN值在14.84—23.82之间。
不同返青期休牧下土壤微生物生物量碳(MBC)差异不显著(P=0.711),但在CK、20 d、30 d、40 d和50 d处理下呈现先增加后降低的趋势,在 30 d处理下 MBC最大,为 56.57 mg·kg−1,在 CK处理下最小,为31.09 mg·kg−1。不同返青期休牧下土壤微生物生物量氮(MBN)差异也不显著(P=0.064),但休牧20 d和40 d的显著高于CK和休牧50 d处理。MBN在休牧20 d和休牧40 d处理下分别为 27.20 mg·kg−1和 30.07 mg·kg−1,在 CK 和50 d 处理下分别为 12.81 mg·kg−1和 11.22 mg·kg−1。但MBC:MBN在不同处理间差异显著(P=0.046),休牧30 d和50 d显著高于休牧20 d和40 d,其中休牧50 d和30 d为3.83和3.82,休牧20 d和40 d为1.53和1.67(表2)。土壤微生物墒(MBC:SOC)在不同返青期休牧下差异不显著(P=0.346)。

表2 不同返青期休牧土壤碳氮及微生物生物量碳氮特征Table 2 The properties of soil C and N and soil microbial biomass Cand N in different grazing-rest times
2.2 不同返青休牧期土壤微生物碳代谢分析
图1为不同返青期休牧处理下土壤微生物群落代谢活性(AWCD)随培养时间的变化曲线,在培养初始的 24 h内土壤微生物活性较低,24 h后AWCD值快速增长,在7 d的时候增加较为缓慢,在第8天出现第二个快速增长期,在第10天达到最大值,但在第11天各处理的AWCD有下降趋势。不同返青期休牧下土壤微生物利用碳源能力的顺序为 CK>30 d>40 d>50 d>20 d,AWCD值分别为(0.85±0.09)、(0.82±0.13)、(0.75±0.08)、(0.70±0.05)和 (0.68±0.01),但各处理间差异并不显著(P>0.05)。

图1 不同返青期休牧下土壤微生物群落平均颜色变化率(AWCD)Fig. 1 Soil microbial AWCD in the different grazing-rest times
高寒草地土壤微生物对不同种类碳源的利用能力是不同,B4在CK显著高于50 d(P=0.048),C2在 20 d显著高于 40 d(P=0.015)和 50 d(P=0.028),D2在 50 d显著低于 20、30、40 d(P=0.012,P=0.018,P=0.013),H4在40 d显著高于50 d(P=0.032),F2在50 d显著低于CK、20 d和 30 d(P=0.016,P=0.020,P=0.035),B3 在 30 d显著高于CK和20 d(P=0.026,P=0.029),H3在50 d显著高于20 d和30 d(P=0.046,P=0.037)(图2)。
土壤微生物对 6类碳源的利用能力也是不同的,依次为羧酸类>氨基酸类>糖类>酯类>醇类>胺类,但不同返青期休牧处理下,只有醇类的代谢差异显著(P=0.028),且休牧20 d显著高于40 d和50 d。其他5类碳源的代谢在不同返青期休牧处理下差异不显著(P>0.05)。不同类的碳源代谢在不同返青期休牧处理下变化很大,很不稳定,并未表现出一定的规律(图3)。

图2 不同返青期休牧下土壤微生物对31类碳源的利用(240 h)Fig. 2 The ability of soil microbial community to utilize the 31 carbon sources in the different grazing-rest times

图3 不同返青期休牧下土壤微生物对六类碳源的利用(240 h)Fig. 3 The ability of soil microbial community to utilize the six kinds of carbon sources in the different grazing-rest times

图4 不同返青期休牧下土壤微生物群落功能多样性指数的比较Fig. 4 Functional diversity indices for soil microbial community in the different grazing-rest times
多样性指数可以反映微生物群落代谢功能的多样性,图4显示土壤微生物群落功能Simpson(D)、Shannon(H)、McIntosh指数(U)和均匀度指数(J)4个指数均在前7天增加较快,7 d以后增加缓慢,且在不同的返青期休牧处理下差异不显著(P>0.05)。Simpson(D)指数在培养的第1天表现出20 d>50 d>40 d>30 d>CK的规律,且值在0.83—0.89之间,但在培养的第11天不同返青期休牧处理下表现出几乎相等的值,约为0.95—0.96(图4A)。Shannon(H)在培养第1天也表现出20 d>50 d>40 d>30 d>CK规律,值在2.25—2.59之间,但在培养的第 11天不同返青期休牧处理下也表现出几乎相等的值,约为 3.15—3.25之间(图 4B)。McIntosh指数(U)在培养的第1天表现出20 d>40 d>CK>30 d>50 d的规律,且值在0.03—0.08之间,但在培养第11天不同返青期休牧处理下表现出30 d>CK>20 d>40 d>50 d的规律,值在4.84—5.64之间(图4C)。均匀度指数(J)在培养第1天表现出20d最高,CK最低的规律,在培养第11天CK是较高的(图4D)。
2.3 土壤微生物特征及其与土壤性质的关系
不同样点的主成分分析(PCA)结果表明:不同返青期休牧处理下土壤微生物群落碳代谢在PCA1和 PCA2上差异都不显著(P>0.05),但在PCA2上,休牧20 d显著高于休牧40 d(P=0.05)。第一主成分和第二主成分的贡献率分别为 75.69%和9.75%,累计贡献率为85.44%(图5)。我们将土壤理化性质及微生物碳氮指标与微生物群落碳代谢功能进行冗余度分析(RDA)分析(图6),结果表明:土壤理化性质及微生物碳氮指标解释了微生物群落碳代谢的91.1%的变异,其中第一主成分解释了80.9%,第二主成分解释了10.2%。土壤有机碳SOC解释了8%的变异,且显著影响微生物群落碳代谢(P=0.038)。另外,全磷(TP)解释了19%的变异(P=0.096),微生物量碳氮比MBC:MBN解释了15%的变异(P=0.072),硝态氮(NO3−-N)解释了14%的变异(P=0.098),微生物生物量碳MBC解释了9%的变异(P=0.124),但这些因子对微生物群落碳代谢的影响差异并不显著。

图5 不同休牧时间下土壤微生物碳代谢功能主成分分析Fig. 5 The principal component analysis (PCA) of soil microbial carbon metabolism function
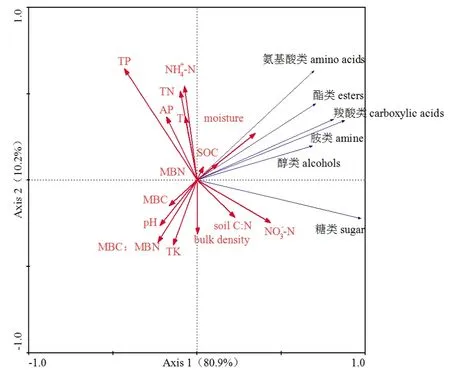
图6 土壤和微生物性质与微生物碳代谢功能的冗余分析Fig. 6 The redundancy analysis (RDA) of soil, microbial properties and the carbon metabolism function
3 讨论
在高寒草甸生态系统,放牧是主要的干扰因子,牲畜可以通过排泄粪便直接的影响土壤生态系统,同时也可以通过采食植物间接的影响土壤生态系统。土壤是草地生态系统重要组分之一,土壤的发育和肥力状况对植物及家畜的生长发育产生重要的影响。土壤生态系统中生活着丰富的微生物类群,其种类、数量、分布等生命活动规律都与土壤中的物质、能量转化、肥力及植物生长有着密切的关系(Bunemann et al.,2004;Schimel et al.,2005)。在我们的研究中,不同的返青休牧期主要差异在于对照是放牧处理,而休牧20、30、40、50 d分别是对草地进行休牧的时间处理,因此,我们不仅要考虑放牧对高寒草地土壤微生物群落的影响,而且要考虑在放牧或者休牧后植物及其根系变化而间接引起的土壤微生物的变化。
家畜等大型食草动物通过改变草地地上和地下凋落物的数量和品质,以及排放尿液和粪便来间接影响植被和生态系统特征(Persson et al.,2005;侯扶江等,2006;张成霞等,2010),适度的放牧干扰有利于地表凋落物的分解和输入,增加土壤养分含量,有利于土壤微生物的繁殖(Xiao et al.,2012;姚国征等,2017;吝亚杰,2017)。我们的研究结果显示在对照不休牧样地土壤微生物群落代谢活性较强,表明放牧活动能提高高寒草甸土壤微生物群落代谢特征,这与翟文婷等(2017)在环青海湖地区高寒草原的结果一致。曲同宝等在松嫩草地上的研究也表明适牧处理下的土壤细菌能充分利用碳水化合物、氨基酸、多聚物、酚酸类及胺类等碳源,休牧处理下的土壤细菌对碳水化合物、氨基酸及多聚物能够充分利用,而重牧处理下的土壤细菌只对羧酸类碳源利用有相对优势(曲同宝等,2012),而长时间的休牧(休牧3—9年)使草原土壤微生物代谢功能增强,土壤微生物繁殖快、数量大,从而促进土壤微生物量碳、氮含量的增加(李玉洁等2013b;薛亚芳等,2018)。由此可见,土壤微生物对碳源的利用与放牧强度和休牧时间有很大的关系,考虑到我们的研究地处于轻度退化高寒草甸,因此在进行放牧的对照处理时,采用轻中度的放牧处理。主要是由于牲畜所产生的粪便能够直接增加土壤中营养元素的含量,加快生态系统的营养循环速率,也有可能是轻中度放牧增加地上植物生产力,刺激光和固定的碳输入到地上根系,增加根系分泌物,从而提高了土壤微生物群落碳代谢活性(Zhao et al.,2017)。但相对于休牧样地,土壤微生物群落代谢活性并未呈出显著的差异,Zhao et al.(2017)的研究结果也表明,尽管放牧条件下土壤微生物群落在土壤碳动态过程中扮演着重要的角色,但轻中度放牧对土壤微生物及其真菌细菌群落无显著影响。Qi et al.(2011)等在中国内蒙古半干旱草原放牧研究中结果表明土壤微生物碳代谢AWCD值在长期休牧样地和连续放牧样地间也无显著差异,但AWCD值和香农维纳指数(H′)随着放牧强度的增加而增加。
从不同返青休牧期对微生物群落代谢功能多样性的影响来看,在培养的第一天休牧20 d的处理下,Simpson(D)、Shannon(H)、McIntosh指数(U)都较其他处理高。休牧20 d处理是返青期后期,在早期(5月10日—6月10日)进行了为期30 d的放牧活动,虽然植物在幼苗期受家畜啃食和践踏的影响,但可能在后期得到充分补偿生长,同时土壤可能受到家畜粪便微生物类群的影响,使得微生物种类和活性有一定的增加,在后期休牧的时候,碳代谢功能得到充分发挥。我们的结果也显示休牧20 d处理下微生物利用醇类物质显著高于休牧40 d、休牧50 d和对照的放牧样地。但凡被微生物用来构成细胞物质或代谢产物中碳架来源的营养物质统称为碳源。微生物能利用碳源的种类及形式极其广泛多样,其中糖类是微生物最广泛利用的碳源,其次是醇类、有机酸类和脂类(鲁顺保等,2012)。微生物对有机物质的分解两类:一、有氧分解,即进入环境的含碳有机物与微生物接触,在有氧条件下这些有机物便成为好氧微生物的营养基质而被氧化分解。二、无氧分解,即当大量有机物进入环境时,由于好氧细菌的活动消耗大量氧气,造成了局部的厌氧环境,使厌氧微生物取代了好氧微生物,而对有机物进行厌氧分解。醇类则需要转化为葡萄糖,进而被分解为无机物质(吴向华等,2012)。由此我们推测,20 d休牧处理前期的放牧活动提高了偏好利用醇类物质的微生物类群种类和活性,从而导致在20 d休牧处理下表现出较高的功能多样性。我们反观对照的放牧处理,其微生物群落代谢功能多样性 Simpson(D)、Shannon(H)、McIntosh指数(U)和均匀度指数(J)在培养第11天的时候相对于其他处理都是较高的。这与陇东典型草原上研究结果一致,丰富度指数和香浓指数在有一定放牧的小区最高,可能是由于土壤硝化细菌种群数量和硝化势随着放牧强度增加了(刘天增,2012)。在不同类型的草地生态系统中,放牧均能增加土壤的硝化活性(Patra et al.,2005)。土壤微生物群落代谢活性(AWCD)和功能多样性在放牧处理下是协同发展的,放牧使微生物活性和种类都增加了。
尽管 Biolog方法能得到很大的微生物代谢功能信息量,但无法得到微生物群落结构的信息。因此,我们仅通过此方法分析不同返青期休牧处理下微生物活动与碳代谢功能,而不考虑微生物类群的变化,尤其跟土壤碳氮比等理化性质密切相关的土壤微生物真菌细菌比的变化。我们用土壤理化性质与土壤微生物碳代谢功能信息做冗余度分析(RDA)试图去寻找微生物群落碳代谢的解释因子,结果发现土壤有机碳显著影响了微生物群落碳代谢特征。相似地,Degen et al.(2000)的研究发现土壤有机碳显著影响碳代谢多样性。李飞等(2018)在高寒湿地和草甸的研究也表明碳氮比、含水量、有机碳、全氮、容重、覆盖度、氮磷比和pH对土壤微生物碳代谢功能多样性有显著的影响。我们的研究中土壤全磷、微生物碳氮比、硝态氮和微生物量氮也影响微生物群落碳代谢特征,但并未达到显著水平。
虽然 Biolog的结果无法直接说明休牧后土壤质量得到一定的恢复或者改善,但土壤微生物墒(微生物量碳/有机碳)被解释为基质的有效性和微生物固持土壤碳的量,反映了土壤中活性有机碳所占比例,是监测土壤有机质的一个有用参数(徐一兰等,2016;易桂田等,2017)。微生物墒高是由于微生物利用有机质的效率高,可作为检测土壤被扰动的指数,微生物墒逐步提高意味着土壤扰动逐步降低而土壤恢复逐步提高,微生物细胞固持更多的有机碳,碳的周转率逐步提高。我们的研究中休牧50 d的微生物熵最大,因此,土壤微生物熵在不同休牧处理间所呈现的规律间接证明了休牧降低土壤扰动,土壤在慢慢恢复。
4 结论
综上所述,我们对高寒草甸生态系统设置返青期不同休牧期的试验处理,通过测定土壤微生物群落碳代谢特征及其土壤理化各因子,结果表明对照不休牧样地土壤微生物群落代谢活性较强,适度的放牧活动能提高高寒草甸土壤微生物群落代谢特征。休牧20 d处理下,微生物利用醇类物质显著高于其他处理。土壤有机碳显著影响了微生物群落碳代谢特征。休牧50 d的微生物熵最大,证明休牧降低土壤扰动,土壤在慢慢恢复。我们的研究可为高寒牧区放牧休牧措施提供土壤方面的基础信息,并且从土壤方面对休牧措施给出科学理论基础。
