果糖处理对冷藏雷竹笋品质和木质化的影响及其调控机制研究
2020-08-20周大祥汪开拓匡文玲雷长毅邱玲岚黎春红蒋永波
周大祥,汪开拓,2*,匡文玲,3,雷长毅,2,邱玲岚,黎春红,蒋永波
1(重庆三峡学院 生物与食品工程学院,重庆,404000) 2(重庆三峡学院 环境与化学工程学院,重庆,404000)3(重庆市万州区食品药品检验所,重庆,404000)
雷竹(Phyllostachyspraecoxf. Preveynalis)为禾本科竹亚科刚竹属竹种,是中国特有优质食用竹种,广泛种植于我国江南、华南及西南各省,雷竹幼体(鞭梢和嫩芽)即为雷竹笋[1]。雷竹笋具有滋味鲜美、硬脆爽口、富含各种维生素及矿物质等优良食用和营养特性,深受市场欢迎。雷竹笋笋体娇嫩,可食用率高,但采割时形成的笋体根部大面积机械伤可导致明显的笋体失重(失水)现象,加快笋体呼吸强度和生理衰老速度,并刺激创面及笋体中心处木质素及纤维素的大量积累,从而使笋体出现质地生硬、粗糙少汁和表面褐变等典型木质化败坏症状,严重降低了雷竹笋食用品质和商品性[2]。低温冷藏(0~5 ℃)虽可有效控制雷竹笋采后失重现象,下调雷竹笋呼吸强度和生理代谢水平,但低温环境也可诱导笋体中木质素合成相关酶活性的上升,促进笋体细胞壁中木质素积累,不利于延长冷藏周期[3]。现阶段研究发现,采用涂膜、气调包装、低压、热激以及外源化学物质处理(如赤霉素、草酸和臭氧等)等方法均可延缓笋体低温木质化症状的发展;但这些处理步骤较繁琐,且笋体木质素单体类物质合成的抑制机制不清晰[4]。
苯丙烷类代谢途径是植物酚酸、花青素、类黄酮、植保素和木质素等多酚类次生代谢产物合成的主要途径,该途径中的主要酶系活性或基因表达丰度在植物体遭受机械伤、病原菌侵染或环境胁迫时显著上调,启动相关逆境响应反应[5]。某些激发子(如茉莉酸甲酯、油菜素内酯或β-氨基丁酸)也可显著诱导采后杨梅[6]、草莓[7]和葡萄[8]等果实植保素、酚酸、类黄酮或花色苷的合成,增强果实贮藏期间抗病性或抗氧化活性。但也有研究发现,杨梅果实[9]和葡萄细胞[10]在花色苷类物质或植保素(白藜芦醇及其脱氢二聚体)大量合成的同时,也伴随着蔗糖及尿苷二磷酸葡萄糖(uridine diphosphate glucose,UDPG)含量的显著下降,其中UDPG正是苯丙烷类代谢途径中合成酚类物质C6-C3-C6骨架的主要底物,可见,蔗糖与苯丙烷类代谢存在对UDPG的竞争性作用。此外,木质素是由香豆醇、松柏醇和芥子醇等单体构成的多聚体,其3种单体在通过苯丙烷类代谢途径合成的过程中也需要大量UDPG来组成碳骨架[11]。因此,木质素合成与蔗糖代谢也可能存在密切联系。本研究拟通过果糖处理来特征性分析采后雷竹笋冷藏期间苯丙烷类与蔗糖代谢的关联性,以期从物质代谢的角度分析雷竹笋木质素合成机理,为延缓雷竹笋采后木质化败坏进程提供可行性方案。
1 材料与方法
1.1 材料、试剂与仪器
雷竹笋于2018年3月中旬和2019年3月上旬采收自重庆市江津区石蟆镇杨柳村雷竹笋种植园,采收后6 h内运回实验室。先剔除切割面不整齐、笋壳脱落或尚有霉菌斑点的雷竹笋,再选择长度和直径均一、外观灰褐色的成熟笋体,平铺并通风散去残余田间热。挑选出的雷竹笋切去老蔸,仔细去壳后备用。
浓H2SO4、丙酮、邻菲罗啉、果糖(分析纯),重庆西南化学试剂公司;葡萄糖、果糖、蔗糖和UDPG标样,美国Sigma公司;RNAprep Pure Kit,北京天根公司;chamQ universal SYBR qPCR master mix、HiScript III RT SuperMix for qPCR,南京Vazyme公司;甲醇、甲酸和乙腈均为色谱纯,中国国药集团有限公司。
PAL-1型手持数显折光仪,日本Atago公司;GL-20G-II型冷冻离心机,上海安亭科学仪器厂;DW-86L型超低温冰箱,海尔公司;TA-XT2i型质构仪,英国Stable Micro System公司;LC-20A型高效液相色谱仪,日本岛津公司;ABI QuantStudio7型实时荧光定量PCR仪,美国Thermo Fisher公司。
1.2 果糖处理
参考XU等[12]的方法进行果糖处理。第一批试验主要分析不同浓度果糖处理对雷竹笋品质的影响,以寻找最适处理浓度。将去壳雷竹笋随机分为5组,悬空平铺于镂空铁丝网上,随后分别用0(对照)、10、20、30或40 mmol/L的果糖溶液进行喷淋处理5 min(20 ℃),期间缓慢翻转笋体以保证喷淋均匀,随后将雷竹笋置于封闭的无菌工作台上充分晾干(避免病原菌和虫蝇污染)。处理结束后,将雷竹笋分装于60 μm厚的聚乙烯袋中,每袋5根笋体,袋口用橡皮筋缠绕2圈,于4 ℃、相对湿度80%~90%条件下贮藏20 d,每5 d取鲜样测定木质化及品质参数。各处理组8袋雷竹笋,重复3次,整个实验重复2次。
第二批试验主要分析果糖处理减轻雷竹笋冷藏期间木质化败坏的机理。将去壳雷竹笋随机分为5组,分别用0(对照)、10、20、30或40 mmol/L的果糖溶液进行处理,方法同上。雷竹笋在4 ℃、相对湿度80%~90%条件下贮藏20 d,每5 d取鲜样经液氮速冻后置于-80 ℃超低温冰箱中保存,测定可溶性糖含量以及相关基因表达水平。各处理组10袋果实,重复3次,整个实验重复2次。
1.3 测定指标及方法
1.3.1 硬度和出汁率的测定
笋体中部硬度参考汪开拓等[13]方法采用TA-XT2i型质构仪进行测定,结果以N/cm2表示。出汁率参考CAO等[14]的方法,将笋体组织圆盘(直径6 mm、高8 mm)置于已经称量的塞有无菌棉的50 mL离心管(m1)中,笋体圆盘及m1质量共计为m2,于2 000×g离心20 min后,对仅装有笋体组织的离心管称重(m3),并按公式(1)计算出汁率:
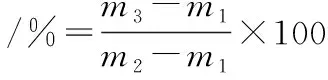
(1)
1.3.2 褐变指数及失重率的测定
褐变度的测定参考JIANG等[15]的方法,略有调整。称取2 g鲜笋肉,研磨后加入10 mL的0.1 mol/L磷酸盐缓冲液(phosphate buffer saline,PBS,pH 7.5),充分匀浆后10 000×g离心25 min(4 ℃),于410 nm处测定上清液吸光值,并以吸光值表示样品褐变度。失重率采用差重法进行测定,并根据公式(2)计算失重率:

(2)
1.3.3 可溶性固形物(toatl soluble solid,TSS)、游离蛋白、抗坏血酸(ascorbic acid,AsA)及1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除能力的测定
笋体TSS和游离蛋白含量参考杨光等[16]的方法进行测定,AsA含量参考邻菲罗啉比色法[17]进行测定,TSS、游离蛋白、抗坏血酸含量分别以溶质的质量百分比(%)、mg/g FW、μg/g FW表示;DPPH自由基清除能力的测定参照RUFINO等[18]的方法,结果按自由基清除百分率来表示。
1.3.4 木质素及纤维素含量的测定
木质素和纤维素含量按照CAO等[14]的方法进行测定,结果均以g/kg FW表示。
1.3.5 可溶性糖组分及UDPG含量的测定
取5 g冻样于液氮下研磨并用30 mL体积分数95%冷乙醇反复萃取3次,萃取液于10 000×g离心15 min(4 ℃)。合并上清液并于旋转蒸发仪(37 ℃)进行真空浓缩,将蒸干后的残留物溶于去离子水,再经0.45 μm的纤维膜微滤后分别按照WILSON等[19]和GOULARD等[20]的HPLC分析法对样品中可溶性糖组分及UDPG含量进行测定。流动相由乙腈(A,75%)和去离子水(B,25%)组成,流速0.8 mL/min,进样量10 μL,柱温35 ℃。HPLC装备Zorbax糖分析柱(250 mm×4.6 mm,10 μm填料粒径)。以外标法计算样品中可溶性糖组分及UDPG含量,结果均以mg/g FW表示。
1.3.6 蔗糖及苯丙烷类代谢途径关键基因表达量测定
取5 g冻样于液氮充分研磨后用RNAprep Pure Kit提取总RNA,再使用HiScript III RT试剂盒将RNA反转录为cDNA第一条链。参照蔗糖合酶分解方向(SS1)、蔗糖合酶合成方向(SS2)、蔗糖磷酸合酶(SPS)、磷酸蔗糖磷酸酶(SPP)、苯丙氨酸解氨酶(PAL)、4-香豆酰CoA连接酶(4CL)、肉桂醇脱氢酶(CAD)、阿魏酸5-羟化酶(F5H)、咖啡酰CoA-O-甲基转移酶(CCOMT)和过氧化物酶(POD)基因序列设计特异性引物(表1),以单链cDNA为模板,以管家基因Actin为内参照进行SYBR Green实时荧光定量PCR(qPCR)试验。根据各基因的循环阈值(Ct值)用定量法(2-ΔΔCt)分析各基因mRNA表达丰度,将对照表达量设置为1以校准各基因的相对表达量[21]。

表1 相关基因特异性引物序列Table 1 Sequences of the gene primers used in this study
1.4 数据分析
数据采取Duncan多重比较法(SPSS 13.0)进行差异显著性检验,5%记录为显著水平(P<0.05),并用Origin 8.5软件作图。
2 结果与分析
2.1 外源果糖处理对冷藏雷竹笋硬度、出汁率、褐变度和失重率的影响
如图1所示,雷竹笋在冷藏期间硬度、失重及笋体褐变情况呈现出逐渐上升的趋势,而出汁率则逐渐下降,表明冷藏期间雷竹笋木质化败坏症状逐渐加剧。10、20或30 mmol/L果糖处理对雷竹笋硬度、失重率和褐变度的上升,以及出汁率的降低有显著(P<0.05)的延缓作用,其中20 mmol/L果糖处理效果最为明显;4 ℃下贮藏20 d后,经20 mmol/L果糖处理的雷竹笋硬度、失重及褐变度仅为对照的76%、68%和79%,而出汁率则为对照的1.57倍。但40 mmol/L果糖处理反而加剧了笋体木质化败坏的症状。

图1 外源果糖处理对冷藏雷竹笋硬度(a)、出汁率(b)、褐变度(c)及失重率(d)的影响Fig.1 Effects of fructose treatments on the firmness (a),extractable juice rate (b),degree of browning (c) and rate ofweight loss (d) in bamboo shoots (Phyllostachys praecox) during cold storage
2.2 外源果糖处理对冷藏雷竹笋品质参数的影响
如表2所示,采用10或20 mmol/L果糖处理可有效提高冷藏雷竹笋游离蛋白和抗坏血酸含量,并增强笋体DPPH自由基清除能力,其中20 mmol/L较10 mmol/L果糖处理效果更为显著。在冷藏结束时,经30或40 mmol/L果糖处理的雷竹笋中抗坏血酸含量显著(P<0.05)高于对照水平,并且经30 mmol/L果糖处理的雷竹笋游离蛋白含量也显著(P<0.05)高于对照水平,但DPPH自由基清除率与对照相比无显著(P>0.05)差异。此外,10~30 mmol/L果糖处理均显著(P<0.05)提高了冷藏雷竹笋TSS含量。

表2 外源果糖处理对雷竹笋冷藏20 d后品质参数的影响Table 2 Effects of fructose treatments on the quality parameters in bamboo shoots after 20 d of cold storage
2.3 外源果糖处理对冷藏雷竹笋木质素及纤维素含量的影响
如图2所示,雷竹笋木质素和纤维素含量随冷藏时间延长而逐渐上升,外源果糖处理(10、20或30 mmol/L)抑制了冷藏期间雷竹笋木质素及纤维素的积累,40 mmol/L果糖处理反而加速了笋体木质素及纤维素的积累。冷藏20 d后,40 mmol/L果糖处理的雷竹笋木质素及纤维素含量分别上升至冷藏前的2.71和3.37倍,而20 mmol/L果糖处理组仅上升至冷藏前的1.82和1.95倍。

图2 外源果糖处理对冷藏雷竹笋木质素(a)及纤维素含量(b)的影响Fig.2 Effects of fructose treatments on the contents of lignin(a) and cellulose (b) in bamboo shootsduring cold storage
2.4 外源果糖处理对冷藏雷竹笋可溶性糖组分及UDPG含量的影响
雷竹笋在4 ℃贮藏期间,葡萄糖及果糖含量在前5 d逐渐下降,随后呈现上升趋势;蔗糖含量持续积累;而UDPG在前10 d逐渐上升,随后缓慢降低(图3)。40 mmol/L果糖处理加剧了雷竹笋冷藏期间可溶性糖组分及UDPG含量的变化;经外源果糖处理(10、20或30 mmol/L)的雷竹笋中葡萄糖、果糖和蔗糖含量在整个冷藏期间均显著(P<0.05)高于对照,然而其UDPG含量则低于对照,其中20 mmol/L果糖处理的冷藏雷竹笋中可溶性糖组分及UDPG含量与对照间的差异最为显著(P<0.05)。
2.5 外源果糖处理对冷藏雷竹笋蔗糖代谢关键基因表达量的影响
蔗糖合酶合成方向(SS2)可催化果糖和UDPG合成蔗糖,同时SPS可将UDPG和6-磷酸果糖合成为6-磷酸蔗糖,再经SPP的作用最终降解形成蔗糖;另一方面,蔗糖可经蔗糖合酶分解方向(SS1)的作用分解为葡萄糖和果糖,并经由糖转运蛋白转运至胞内[22]。如图4所示,外源果糖处理(10、20或30 mmol/L)诱导了冷藏雷竹笋PpSS1表达量的下调,并显著(P< 0.05)上调了PpSS2、PpSPS、PpSPP表达量,其中20 mmol/L果糖处理对雷竹笋冷藏期间蔗糖代谢关键基因表达量的调控作用最为显著(P<0.05);而40 mmol/L果糖处理对冷藏雷竹笋蔗糖代谢关键基因的调控与10、20或30 mmol/L果糖处理恰好相反。

图3 外源果糖处理对冷藏雷竹笋葡萄糖(a)、果糖(b)、蔗糖(c)及UDPG含量(d)的影响Fig.3 Effects of fructose treatments on the contents of glucose (a),fructose (b),sucrose (c) and UDPG (d) in bambooshoots during cold storage

图4 外源果糖处理对冷藏雷竹笋PpSS1(a)、PpSS2(b)、PpSPS(c)和PpSPP(d)表达丰度的影响Fig.4 Effects of fructose treatments on the expression levels of PpSS1 (a),PpSS2 (b),PpSPS (c) andPpSPP (d) in bamboo shoots during cold storage注:不同小写字母表示差异显著(下同)
2.6 外源果糖处理对冷藏雷竹笋苯丙烷类代谢关键基因表达量的影响
木质素的合成途径主要由苯丙氨酸途径和木质素合成的特异途径所组成,其中PAL作为苯丙烷途径的关键限速酶;4CL位于苯丙氨酸生物合成途径的分支点,主要参与次生代谢产物合成并调控植物木质素代谢;CAD、CCOMT能催化木质素前体物质转化为木质素单体;F5H主要调控木质素单体的生物合成;POD是催化木质素生物合成途径最后一步的关键酶,这几种酶与木质素的合成密切相关[11]。如图5所示,外源20 mmol/L果糖处理显著(P<0.05)下调了冷藏雷竹笋苯丙烷类代谢关键基因(PpPAL、Pp4CL、PpCAD、PpF5H、PpCCOMT和PpPOD)的表达量;10或30 mmol/L果糖处理也一定程度诱导了苯丙烷类代谢关键基因表达量的下调;而40 mmol/L果糖处理对雷竹笋冷藏前10 d内苯丙烷类代谢关键基因的表达有促进作用。

图5 外源果糖处理对冷藏雷竹笋PpPAL(a)、Pp4CL(b)、PpCAD(c)、PpF5H(d)、PpCCOMT(e)和PpPOD(f)表达丰度的影响Fig.5 Effects of fructose treatments on the expression abundance of PpPAL (a),Pp4CL (b),PpCAD (c),PpF5H(d),PpCCOMT (e) and PpPOD (f) in bamboo shoots during cold storage
3 讨论
3.1 最适果糖处理浓度的筛选
可溶性糖作为果蔬感官品质的主要物质来源,与果蔬采后生理、功能性或营养成分、风味、呈色等重要品质参数密切相关;并作为植物体中能量来源和渗透调节物质,具有为各种代谢过程提供前体物质的功能[23]。相关研究表明,采后蔗糖处理可有效降低青花菜线粒体膜电位并阻止细胞色素C从线粒体中释放,从而延缓青花菜程序性死亡进程和生理衰老[24];蔗糖或葡萄糖处理则可维持青花菜中叶绿体超微结构的完整性,抑制叶绿素降解相关酶基因表达,延缓叶绿素降解,抑制花蕾黄化症状的发展[12,25];可溶性糖(蔗糖、果糖和葡萄糖)复合处理则能上调拟南芥花色苷合成相关基因的表达,促进花色苷的合成[26]。在这些研究中,果糖及其他可溶性糖既能直接通过蔗糖代谢来实现对果蔬品质的调控,也可作为激发子来诱导糖代谢关键酶(如己糖激酶)活性以启动下游酶蛋白的磷酸化作用,从而延缓生理衰老进程或促进次生代谢产物合成。本研究中,在所有处理浓度中,20 mmol/L果糖处理可最为显著地抑制雷竹笋冷藏期间木质素和纤维素的积累,延缓笋体硬度、失重率和褐变度的上升,并提高出汁率(图1和2);在冷藏结束的时候,经20 mmol/L果糖处理的雷竹笋中TSS、游离蛋白、抗坏血酸含量及自由基清除能力均显著高于其他浓度处理或对照水平(表2)。因此,20 mmol/L果糖处理可有效抑制雷竹笋木质化败坏进程,减慢笋体品质劣变速度,从而延长冷藏周期。由于果糖处理可使蔗糖代谢偏向于合成方向从而大量消耗UDPG,因此可减少木质素底物含量,进而限制雷竹笋木质素的合成[22]。另有研究发现,果糖处理能直接诱导费菜中苯丙烷类代谢酶活性的上升,并提高类黄酮合成量和抗氧化活性[27],这可能与果糖处理直接诱导己糖激酶基因的表达,并促进苯丙烷类代谢途径关键酶的活化有关。
3.2 果糖处理抑制雷竹笋木质素败坏的机理分析
植物木质素为多种苯丙烷单体的共聚物,其单体主要通过苯丙烷类代谢途径,以L-苯丙氨酸为底物合成中间体UDPG,再经羟基化、甲基化和连接反应最终生成;该途径的主要酶系有苯丙氨酸解氨酶(PAL)、4-香豆酰CoA连接酶(4CL)、咖啡酰CoA-O-甲基转移酶(CCOMT)、阿魏酸5-羟化酶(F5H)、肉桂醇脱氢酶(CAD)、过氧化物酶(POD)[5]。蔗糖或己糖是果实中碳水化合物运输的主要形式,常被用来研究对果实生长发育和基因表达的调控作用。蔗糖代谢相关酶活性与果实可溶性糖积累之间存在密切联系,其中包括蔗糖合酶(SS)、蔗糖磷酸合酶(SPS)和磷酸蔗糖磷酸酶(SPP)等酶系[22];UDPG则在蔗糖代谢途径中作为单糖基合成蔗糖或作为供体分解为单糖[28]。对模式植物拟南芥的研究表明,当植株受到病原菌侵染时,其组织中UDPG急剧消耗以合成植保素或花色苷类等抗菌物质,控制病害症状的发展[29]。因此,苯丙烷类次生代谢产物与蔗糖在合成路径上存在对底物UDPG的竞争。在本研究中,当施加外源果糖处理后,雷竹笋蔗糖代谢途径中的PpSS2(合成方向)、PpSPS和PpSPP基因表达量在整个冷藏期间显著上调而PpSS1(分解方向)基因表达量显著下调(图4),使蔗糖代谢通路偏向于合成方向,从而引起笋体蔗糖含量上升和UDPG含量下降(图3);同时,笋体苯丙烷类代谢途径中主要关键酶基因(PpPAL、Pp4CL、PpCAD、PpF5H、PpCCOMT和PpPOD)表达丰度在贮藏期间均显著下降(图5),并伴随着木质素合成量的下降,这极有可能是由于UDPG过度用于蔗糖合成从而对木质素合成底物产生竞争性消耗。前期研究也证实,葡萄悬浮细胞经高浓度苯丙噻唑硫代乙酸甲酯或β-氨基丁酸处理后,其细胞内蔗糖分解酶活性显著上升,分解出UDPG以供给茋类植保素和花色苷的合成,从而降低细胞中可溶性糖含量和细胞生长速率[10,30]。这些结果说明,外源果糖处理可通过调控蔗糖代谢的方式来降低UDPG含量,从而减少雷竹笋中木质素生成量,减轻冷藏周期内笋体木质化败坏症状,延长贮藏周期或货架期。
4 结论
20 mmol/L果糖喷淋处理可有效抑制雷竹笋冷藏期间笋体硬度上升、褐变和少汁等木质化败坏症状的发生,降低笋体失重率,维持较高的出汁率和感官品质,从而有效延长雷竹笋冷藏周期。20 mmol/L果糖处理可通过调控雷竹笋蔗糖代谢途径的方式,竞争性抑制笋体木质素的合成,但果糖在此过程中是否发挥其他的调控功能(如己糖信号分子作用)仍有待研究。
