螺旋筛板气升式反应器在真菌发酵中的应用
2020-08-19李想陈瑜琦王子凡郑志永陈海琴高敏杰詹晓北
李想,陈瑜琦,王子凡,郑志永,2*,陈海琴,高敏杰*,詹晓北
1(糖化学与生物技术教育部重点实验室(江南大学),江苏 无锡,214122) 2(江南大学 环境与土木工程学院,江苏 无锡 214122)3(江南大学 食品学院,江苏 无锡,214122)
螺旋筛板气升式反应器(airlift reactor with helical sieve plate,ALR-HSP)是本研究室在经典气升式反应器(conventional airlift reactor,CALR)的基础上研发出来的,靠导流筒装置的引导使多相进行循环混合的反应器,它与CALR的主要区别在于其导流筒上升段安装有螺旋上升的多孔筛板[1-2]。在空气-水试验中,ALR-HSP展现出比CALR更好的传质性能和节能的特性,装有螺旋筛板的这种结构能够有效阻止气泡聚并,增大气液比表面积从而提高传质效率,在相同通风量条件下气含率和体积传质系数分别增加38%~53%和76%~144%[3]。
气升式反应器虽然具有结构简单、剪切力小和能耗低的特性,但由于其能调控的参数较少,操作鲁棒性差,传质效率较低,因此工业上只有少量菌株能够用气升式反应器来培养。导流筒上安装螺旋筛板后,反应器的传质性能得到提高,操作鲁棒性明显改善,这增加了工业菌株用气升式反应器来培养的适用性。真菌具有生长速度较慢、耗氧量较少和对剪切力比较敏感的特性,是潜在的可以使用气升式反应器来进行培养的菌株。
高山被孢霉(Mortierellaalpine)是1株生产多不饱和脂肪酸(polyunsaturated fatty acids,PUFAs)的丝状真菌[4-5]。其对剪切力敏感[6],但由于CALR不能满足其耗氧需求,因此高山被孢霉的培养依旧选择机械搅拌式反应器。在采用机械搅拌式反应器培养时转速不宜过高,这在一定程度上减弱了机械搅拌罐在操作鲁棒性上较强的优势,使用ALR-HSP有望解决这个问题。另外毕赤酵母是一种应用广泛的外源蛋白表达系统,也是一种在工业发酵中常用的菌株,能生产出大量的重组蛋白质[7-8]。同样因CALR的供氧能力有限,不能满足菌株的耗氧需求,因而使用机械搅拌罐[9-10]。本文采用传统气升式反应器和带有螺旋筛板气升式反应器,对高山被孢霉和毕赤酵母进行培养,分别考察这2株菌的发酵过程参数,分析螺旋筛板在耗氧微生物发酵中产生的影响。
1 材料与方法
1.1 材料
1.1.1 实验装置
发酵用ALR-HSP是以比欧SSTC型65 L气升式反应器(瑞士比欧生物工程公司)作为改造对象,在反应器内安装自行设计的导流筒、开孔率61%的螺旋筛板与圆锥面形的气体分布器,具体样式如图1所示。本实验的反应器有2个,一个是导流筒上有螺旋筛板的反应器ALR-HSP,作为实验组;另一个是导流筒上没有螺旋筛板的反应器CALR,作为对照组。实验装置主要结构参数如表1所示。
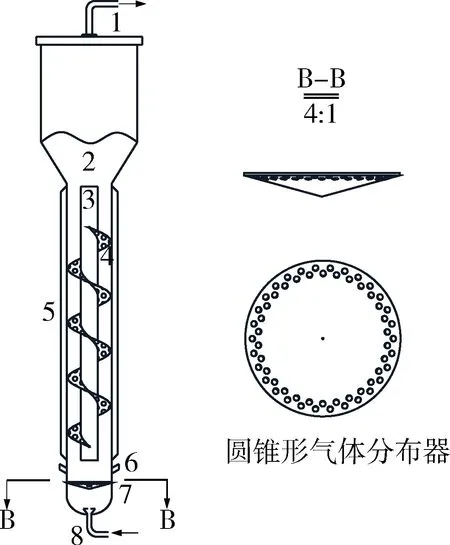
1-出气口;2-气液分离区;3-导流筒;4-螺旋筛板;5-夹套;6-溶氧电极;7-圆锥形气体分布器;8-进气口图1 螺旋筛板气升式反应器Fig.1 Experimental device of helical sieve airlift
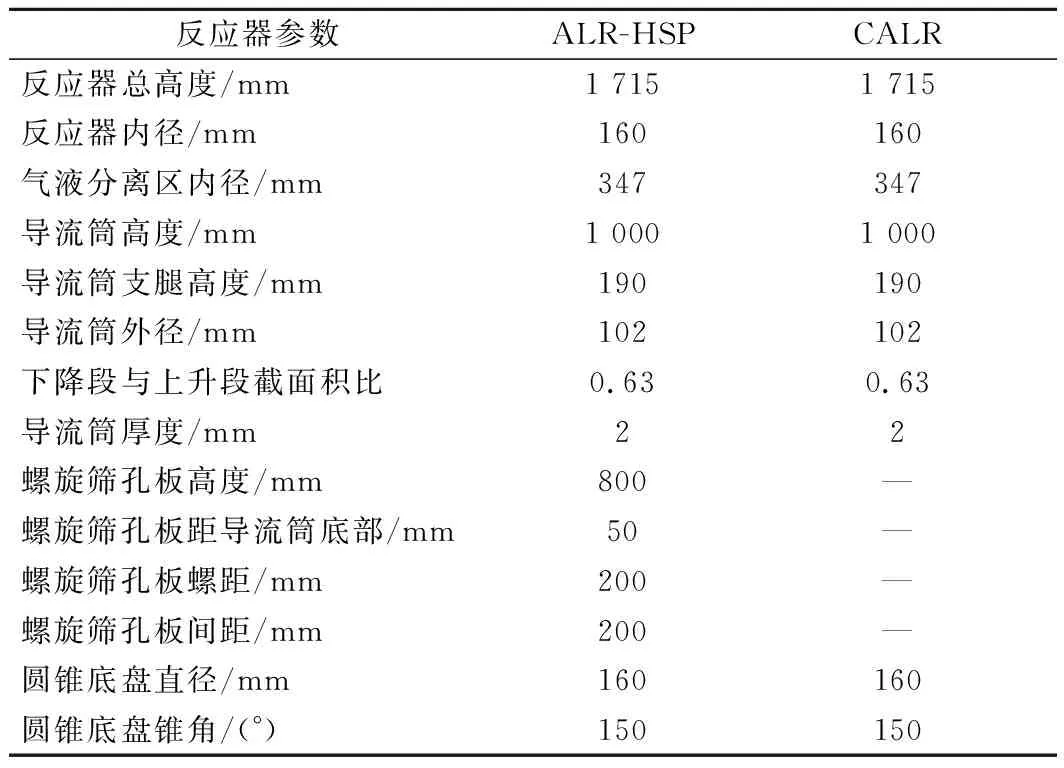
表1 实验气升式反应器的结构参数Table 1 Structure parameters of airlift experiment reactor
1.1.2 菌株与试剂
高山被孢霉(MortierellaalpineATUS2222)由江南大学食品学院赠送。毕赤酵母(PichiapastorisKM71)购于Invitrogen公司。
酵母粉,北京索莱宝科技有限公司;甘油、K2SO4、KH2PO4、MgSO4、KOH、磷酸等,国药集团化学试剂有限公司。
1.1.3 培养基
1.1.3.1 高山被孢霉培养基
种子培养基(g/L):葡萄糖 20,酵母提取物 5,KH2PO41,MgSO4·7H2O 0.51,KNO310。
发酵培养基(g/L):葡萄糖 30,酵母提取物 1.5,酒石酸铵 2.0,KH2PO47,MgSO4·7H2O 1.5,Na2HPO410,无水CaCl20.1,FeCl3·6H2O 8 mg/L,ZnSO4·7H2O 1 mg/L,CuSO4·5H2O、Co(NO3)2·6H2O、MnSO4·5H2O 0.1 mg/L。
1.1.3.2 毕赤酵母培养基
酵母浸出粉胨葡萄糖(YPD)种子培养基(g/L):酵母粉 10.0,葡萄糖 20.0,蛋白胨 20.0。
发酵培养基(g/L):甘油 26.0,K2SO41.0,MgSO41.0,CaSO40.1,KOH 20.0,(NH4)2SO45.0,PTM1 10 mL,H3PO420 mL/L,pH 6.0。
1.2 实验方法
1.2.1 培养方法
1.2.1.1 高山被孢霉
种子液制备:从斜面上挑取活化后的菌种接种至装有100 mL种子培养基的250 mL三角瓶中,200 r/min、28 ℃培养48 h后,按体积分数1%的接种量接种,使用分散器打散的菌丝体片段,并传代2次,每次均在200 r/min、28 ℃条件下培养36 h,即得到种子液。
发酵罐培养:将36 L的初始发酵培养基置于65 L的气升式反应器内,灭菌,接种前调节发酵培养基维持在28 ℃、pH 6.0。加入4 L的种子液,在恒定通气量10 L/min条件下进行培养。
1.2.1.2 毕赤酵母
种子液制备:将保存在甘油管中的菌体接种到斜面培养基上,放入30 ℃培养箱培养2 d。将单个菌落接入装有50 mL培养基的500 mL三角瓶,温度30 ℃,转速220 r/min,培养18 h,直至OD600达到2.0。
发酵罐培养:将28.5 L的初始发酵培养基置于65 L的气升式反应器内,灭菌,接种前调节发酵培养基维持在30 ℃、pH 6.0。加入1.5 L的种子液,在恒定通气量40 L/min条件下进行培养,待甘油耗尽后停止培养。
1.2.2 细胞浓度测定
将用去离子水洗涤过的高山被孢霉进行冷冻干燥,采用干重法测定菌体浓度[11],采用浊度法测定毕赤酵母菌体浓度[12]。
1.2.3 碳源质量浓度测定
1.2.3.1 葡萄糖质量浓度测定
采用SBA-40E型生物传感仪(山东省科学院生物研究所)测定[13]。
1.2.3.2 甘油质量浓度测定
发酵液中残余甘油浓度通过HPLC法测定[12]。
1.2.4 NH4-N含量测定
采用水杨酸钠-次氯酸钠比色法[14]。
1.2.5 发酵过程中OUR及kLa测定
通过LKM2000A型尾气分析仪(韩国 LOKAS)记录发酵尾气中O2的分压,按照公式(1)和(2)分别计算O2消耗速率(oxygen uptake rate,OUR),体积传质系数(kLa),计算公式如下:
(1)
(2)
式中:OUR,O2消耗速率,mmol/(L·h);Qg,通气流量,m3/s;φO2,氧气的体积分数,%;V,装液体积,m3;ρ*,饱和氧质量浓度,mg/L;ρL,实际溶氧质量浓度,mg/L。
1.2.6 脂质含量测定
样品制备方法如下:将冷冻干燥的菌丝体碾碎,精密称取约50 mg置于样品瓶中,提取脂肪酸及甲酯化具体操作见文献[15]。
采用QP2010型气相色谱-质谱(日本 Shimadzu公司)分析脂肪酸。色谱柱为DB-WAXetr(30 m×0.32 mm,0.25 μm)。柱箱温度在150 ℃下保持3 min,然后以10 ℃/min升至190 ℃,随后以5 ℃/min升至220 ℃持续16 min。质谱在m/z50~550的扫描范围内运行,离子源温度为220 ℃,电子轰击源能量为70 eV,进样量为1 μL。按照上述分析方法得到不饱和脂肪酸甲酯的总离子流图,如图2所示。并通过与商业脂肪酸甲酯标准(GLC.463,Nu-Chek,Elysian,MN,美国)进行比较,鉴定脂肪酸甲酯,鉴定结果如表2所示。

图2 高山被孢霉产油脂的总离子流图Fig.2 Total ion chromatograms of lipids produced byM. alpine
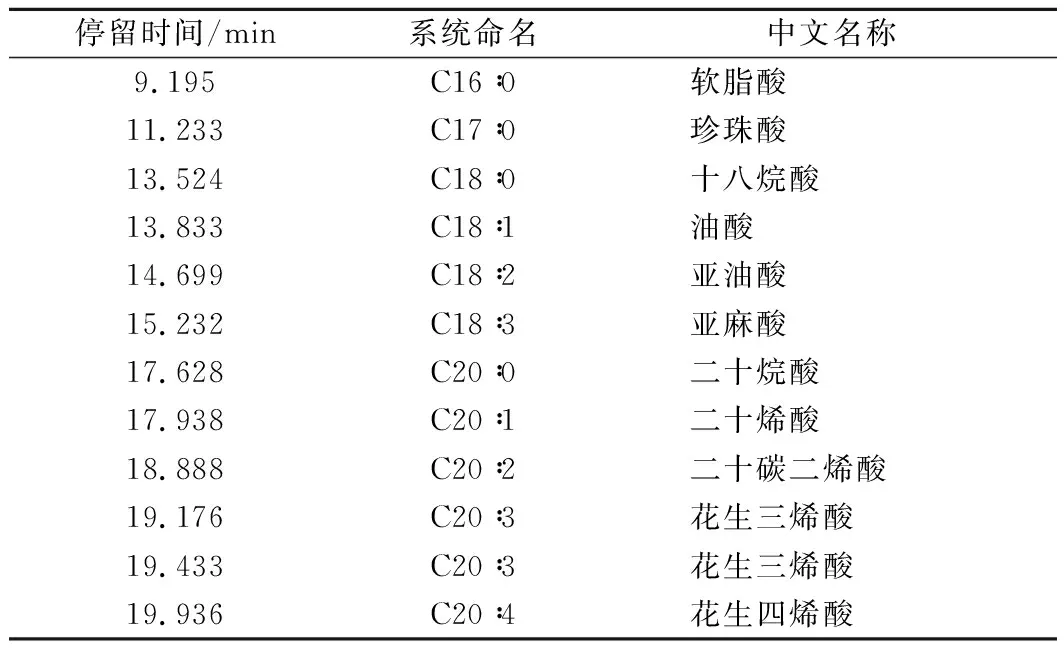
表2 高山被孢霉合成的油脂组分分析Table 2 Components of lipids produced by M. alpine
2 结果与讨论
2.1 发酵过程曲线
高山被孢霉和毕赤酵母的发酵过程曲线如图3所示。高山被孢霉的生长可以分为延滞期、对数期(8~40 h)、稳定期3个阶段(图3-a,b)。反应器在对数期,菌体量增多代谢加快,导致2个反应器的溶解氧(dissolved oxygen,DO)浓度均迅速下降,培养基中的底物浓度迅速降低。在对数生长后期,2反应器的DO值差别显著,ALR-HSP和CALR的DO值分别在35%和10%左右后趋于稳定。在培养40 h后,培养基中氮源的耗尽限制了菌体生长,菌体浓度趋于稳定,菌体生长进入稳定期。培养结束后,ALR-HSP中的菌体浓度比CALR增加约10%。菌体浓度增加可能是在ALR-HSP中氧气供应更为充足,更适合高山被孢霉的生长[16]。
毕赤酵母生长过程可分为延滞期、对数期、稳定期3个阶段(图3-c,d)。菌体进入对数生长期(12~32 h)后,代谢加快,耗氧量增加,2个反应器的DO值均开始下降,并出现明显差异。菌体在此阶段耗氧量大,而气升式反应器的供氧能力有限,致使2反应器的溶氧先后降为0。直到36 h后,ALR-HSP和CALR的DO值开始上升,此时甘油质量浓度过低限制了菌体的生长,菌体进入稳定期(36~48 h)。最终ALR-HSP中菌体的浓度比CALR增长约20%。其原因可能是CALR中菌体在溶氧限制条件下产生了大量副产物导致细胞对底物的得率系数较低[7,17-18]。

a-高山被孢霉溶解氧体积分数和生物量曲线;b-高山被孢霉葡萄糖质量浓度和NH4-N质量浓度曲线;c-毕赤酵母溶解氧体积分数和生物量曲线;d-毕赤酵母甘油质量浓度和NH4-N质量浓度曲线实心-ALR-HSP;空心-CALR;三角形-菌体浓度;菱形-溶解氧体积分数;圆形-NH4-N质量浓度;正方形-葡萄糖/甘油质量浓度图3 高山被孢霉和毕赤酵母在ALR-HSP和CALR中的发酵过程曲线Fig.3 Process curves for the M. alpine and P. pastoris cultivation in the ALR-HSP and CALR
2.2 生长速率与比生长速率
高山被孢霉在2种反应器内培养的生长速率曲线和比生长速率分别如图4-a,b所示。菌体进入对数期后,生长速率迅速增加,并在28 h达到峰值后开始变慢。根据测定溶氧生长极限值(当DO值低于生长限制值时,菌体生长受到限制)的方法[16]测得本实验所用菌体的生长极限值约为30%。在营养物充足的情况下,菌体生长速率和比生长速率降低是由于DO限制造成[19]。ALR-HSP培养菌体的最大生长速率和最大比生长速率分别比CALR提高25%和30%,这说明了ALR-HSP在氧传递性能上的优越性。图4-c,d为毕赤酵母在两种反应器内培养的生长速率曲线和比生长速率曲线。菌体生长到12 h后开始进入对数生长期,此时生长速率迅速增加,到20 h后,设备的最大供氧能力限制着菌体的生长速率。由于ALR-HSP在供氧能力优越性,使得ALR-HSP的细胞生长速率和比生长速率分别比CALR提高40%和20%。
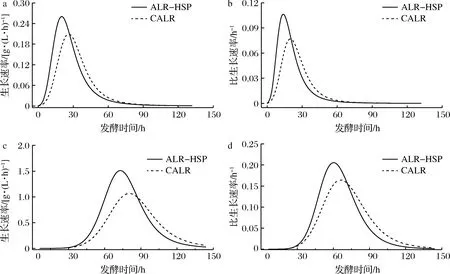
a-高山被孢霉生长速率曲线;b-高山被孢霉比生长速率曲线;c-毕赤酵母生长速率曲线;d-毕赤酵母比生长速率曲线图4 高山被孢霉和毕赤酵母在ALR-HSP和CALR中培养的生长速率与比生长速率Fig.4 Growth rate and specific growth rate for the M. alpine and P. pastoris cultivation in the ALR-HSP and CALR
2.3 高山被孢霉单位菌体脂肪酸组成分析
各脂肪酸的量随着发酵时间不同而发生变化,脂肪酸总量随发酵时间延长增多,最后趋于恒定(图5)。相同培养时间下(发酵时间大于48 h),ALR-HSP中高山被孢霉的脂肪酸总含量明显高于CALR。2个反应器对高山被孢霉中脂肪酸组成的影响如表3所示。从表3中可知,ALR-HSP中的菌体产油脂的总量比CALR提高40%,菌体对碳源的得率提高了6%,总油脂量对碳源的得率提高了45%。菌体中产生几种主要脂肪酸含量存在显著差异,对比脂肪酸种类发现,ALR-HSP中菌体产生的脂肪酸的不饱和度比CALR中的高。其原因可能是在供氧充足时,菌体更容易合成不饱和度高的脂肪酸[20-21],这说明ALR-HSP的供氧能力强于CALR。

1-ALR-HSP;2-CALR图5 不同培养时间下,在ALR-HSP和CALR中培养的高山被孢霉的单位菌体中脂肪酸组成变化Fig.5 Fatty acid composition changes of M. alpine plantscultivated in ALR-HSP and CALR under differentculture time

表3 两反应器对高山被孢霉中脂肪酸组成的影响Table 3 Effects of two reactors on fatty acid composition by M. alpine
2.4 氧气消耗速率(OUR)和体积传质系数(kLa)
从图6中可以看出,2个反应器中菌体的OUR均在短时间内迅速上升,并在36 h左右开始达到最大,之后由于反应器供氧有限和氮源匮乏等问题,使得菌体生长受限,致使OUR迅速下降。通风发酵过程中的DO同时取决于传氧速率(oxygen transfer rate,OTR)和OUR。其中OUR取决于细胞密度、代谢活力和营养条件,OTR取决于设备性能、操作条件和发酵液性质。在恒定通风条件下,可近似地认为OTR主要与设备性能有关。又在稳态条件下,有OTR=OUR。因此在稳态条件下,微生物的OUR越高,其对应反应器的性能越好。在48~96 h期间DO值无明显变化,此时发酵过程达到稳定状态,有OTR=OUR。从图6-a中可得出有筛板反应器中菌体的OUR比无筛板反应器中的菌体的OUR高21%~36%。
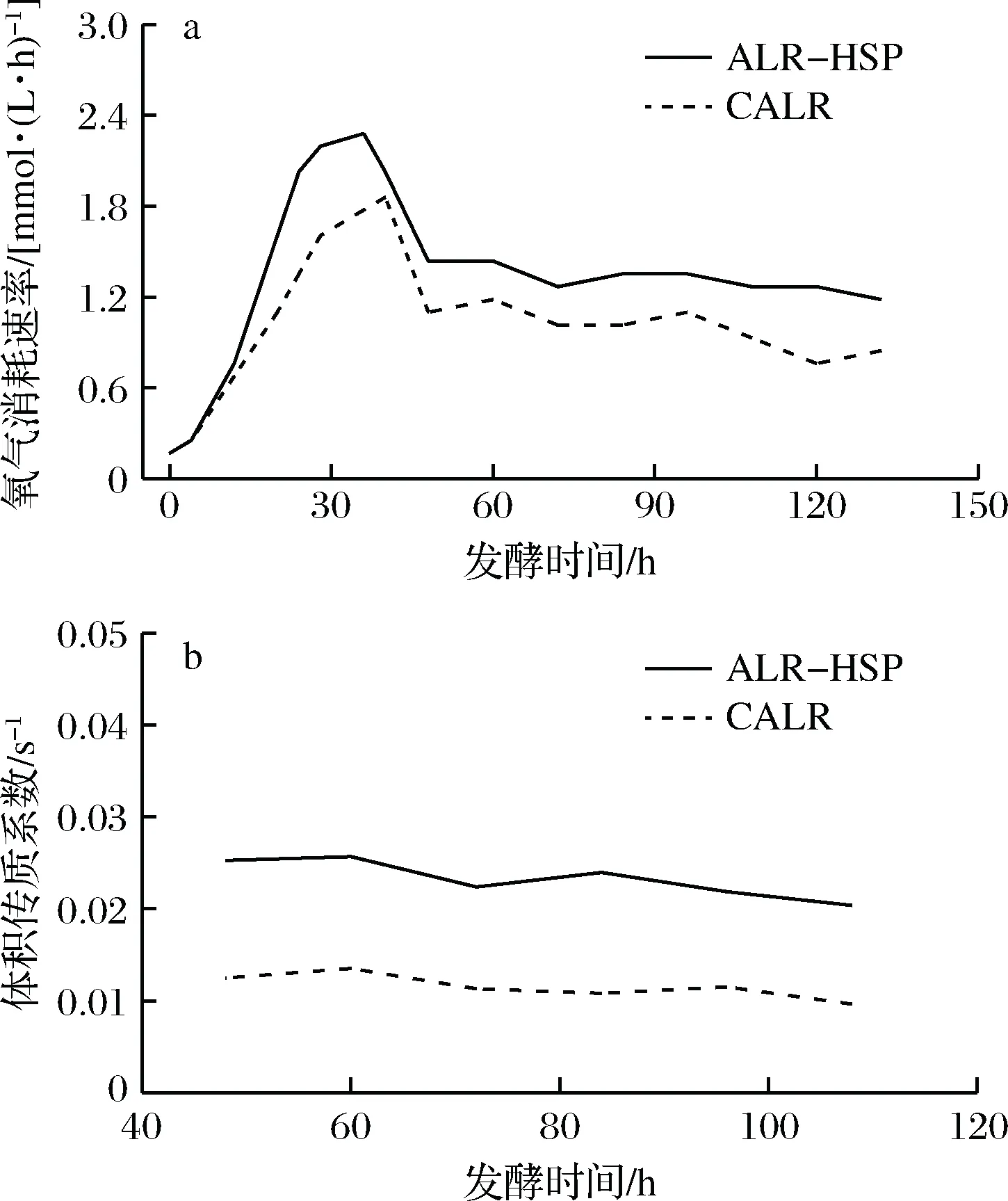
a-高山被孢霉培养过程中氧气消耗速率变化;b-高山被孢霉培养过程中体积传质系数变化图6 使用ALR-HSP 和 CALR培养高山被孢霉过程中的氧气消耗速率和体积传质系数Fig.6 The comparison of OUR and kLa forM. alpine in the ALR-HSP and CALR
图6-b为培养体系在稳态条件下,根据公式(2)计算出来的对应发酵时间的体积传质系数(kLa)。在通气量为10 L/min条件下,ALR-HSP和CALR的kLa值分别为(0.023 2±0.002 1) s-1和(0.011 5±0.001 3) s-1。ALR-HSP的kLa比CALR提高1倍多,这进一步说明了ALR-HSP的优势。在进行通风发酵实验前,对反应器进行空气-水实验,采用动态驱赶法[22]测得在通气量10 L/min条件下,ALR-HSP和CALR的kLa分别为(0.017 3±0.001 6) s-1和(0.009 1±0.000 6) s-1。与使用尾气分析仪测出来的kLa数值接近,但也存在差异,其原因可能是测定方法和测量介质不同[23]。
气升式反应器的主要能量消耗来源于空压机,通风量越大,能耗越大。在气升式反应器中,牛顿流体常用公式(3)[23]估算kLa,同理也可以使用此公式计算kLa对应的表观气速(Ug),公式(4)[24]可用来计算通气过程的功率消耗。
(3)
(4)
式中:C和a,常数,在纯水条件下C=0.47,a=0.82;Ug,表观气速,m/s;P,空压机功率,kW;pi,空压机进口压力,bar;po,空压机出口压力,bar;Qg,通气流量,m3/s。
根据公式(3)和(4)计算得出,CALR要达到ALR-HSP的kLa需要多消耗1倍的功率,这说明ALR-HSP的能量利用效率更高。
培养过程中现场观察发现,ALR-HSP中液面以上的泡沫数量明显比CALR多。在有筛板条件下,大气泡被分割为更多的小气泡,小气泡的热力学性质稳定,不容易破碎,不断在液面上方累积。这是今后需要研究者解决的问题,为解决气泡过多问题,可以与其他消泡设备同时使用,例如:离心消泡[25]和旋击分离器[26]。另外在灭菌结束后高山被孢霉的丝状菌株会附着在筛孔板上,使清洗变得复杂。但总体上,ALR-HSP相比于CALR,其传质效率和能量利用率上均得到提高,这在提升气升式反应器应用范围方面具有显著的工业价值。
3 结论
螺旋筛板气升式反应器(ALR-HSP)在毕赤酵母和高山被孢霉的培养过程中表现出了比经典气升式反应器(CALR)更加优良的传质特性和节能特性。在高山被孢霉的培养中,在相同功率消耗条件下,ALR-HSP内菌体的最大生长速率和最大比生长速率分别比CALR提高25%和30%,菌体量增长约10%,脂肪酸总量提高约40%;在毕赤酵母培养中,在相同功率消耗条件下,ALR-HSP内的菌体浓度增长20%,最大生长速率和最大比生长速率分别比CALR提高40%和20%。在低通风量条件下,ALR-HSP的传质效率比CALR提高了1倍多。但在培养过程中ALR-HSP也存在一些问题,如在液面上泡沫更多和灭菌后的清洗更为复杂。这些问题有待进一步的研究和解决,以期待这种新型的气升式反应器能够在工业上取得广泛应用。
