披麻草野生居群表型多样性分析研究
2020-08-14尹子丽冯泽辉陈静怡
尹子丽 冯泽辉 陈静怡
摘要:目的 分析披麻草野生居群表型多样性。方法 对12个居群植株叶片长、叶片宽、叶片数、叶基长度、叶基直径等 5 个性状进行测量,并测定居群土壤pH值和肥力指标。所得数据采用Excel 和 SPSS(19.0)软件进行描述性、单因素方差和 LSD 多重比较处理分析。结果 披麻草变异系数最大的为叶基直径,最小的为叶基长度;叶片长、叶片宽、叶片长/叶片宽、叶数、叶基长度、叶基直径各项性状在各居群间均有极显著性差异(P<0.01)。全盐量、有效磷及速效钾含量在居群间并无一定的规律可循,地上部分表型特征不能作为全盐量、有效磷和速效钾的直观性指标。结论 披麻草居群间的环境差异及个体间的小环境差异对披麻草生长过程中的形态变化产生很大的影响。
关键词:披麻草;野生居群;表型多样性
【Abstract】Objective:To study the wild population phenotypic diversity of Veratrum taliense Loes. Methods:Five characters such as leaf length,leaf width,leaf number,leaf base length and leaf base diameter of 12 population plants were measured,and the pH value and fertility index of the population soil were determined. The data obtained were analyzed by using Excel and SPSS(19.0)software for description,one-way variance and LSD multiple comparisons. Results:The largest coefficient of variation of Veratrum taliense Loe was leaf basal diameter,and the smallest was leaf basal length,and the leaf length,leaf width,leaf length-width ratio,leaf number,leaf basal length and leaf basal diameter had extremely significant differences(P<0.01)among the populations. There is no certain rule for the total salt content,available phosphorus and available potassium content among the populations,and the aboveground phenotypic characteristics cannot be used as visual indicators of the total salt content,available phosphorus and available potassium. Conclusion:The environmental differences among the populations and the small environmental differences among individuals of Veratrum taliense Loe have a great influence on the morphological changes during the growth of Veratrum taliense Loe.
【Key words】of Veratrum taliense Loe,wild population,phenotypic diversity
披麻草是国家保密配方制剂“云南白药”的主要组成药材之一,也是国家中药保护品种“一粒止痛丸”的主药,收载于1996年版《云南省中药材标准》[1],具有活血散瘀、消肿止痛、杀虫毒之功效,用于中风痰壅、喉不通畅、跌打损伤、风湿疼痛、创伤止血、疥癣、恶疮等。目前,披麻草尚未收入《中国药典》,全国各地多就地取材,地产地销,未形成有效的保护和统一规范的种植。近年来,由于市场需求量增加,西南地区持续干旱以及部分地区土地使用的变迁,采挖价格持续走高,同时由于人为干预,现有居群中成年繁殖期个体比例较低,种群资源量的稳定性难以维系,致使资源将会或者已经趋于渐危状态。因此,开展披麻草种质资源表型多样性与生境相关性的研究,及时制定该物种的保护策略是非常有必要的。
目前,披麻草的种质资源研究主要集中在生物学特性、生药学鉴定、遗传多样性分析、适生区等研究上,披麻草种质资源相关研究表明:披麻草的花序结构、居群分布特征和种群资源量的不稳定性直接导致其自然繁殖率和资源量的大幅锐减,遗传多样性大量喪失,趋于渐危状态[2-6]。因此,本文以野生居群为实验材料,进一步了解披麻草表型间的差异及多样性的变化规律。表型是指在一定环境下,植物所表现出的性状总和,是基因与环境共同作用的结果[7]。土壤是重要的环境因子,同时也是植物正常生长发育的基础,对其植物的表型和产量影响较大[8]。植物种群的形态结构与当地生态环境有密切关系,研究各个居群分布区内各种环境下的表型变异,是了解其遗传变异的重要线索,也是生物多样性与生物系统学研究的重要内容[9]。本文通过研究12个披麻草野生居群的表型性状的多样性及其与土壤因子的相关性,以期揭示披麻草种质资源的表型多样性及其分布规律,并为披麻草野生种质资源的保护提供相关理论参考依据。
1 材料和方法
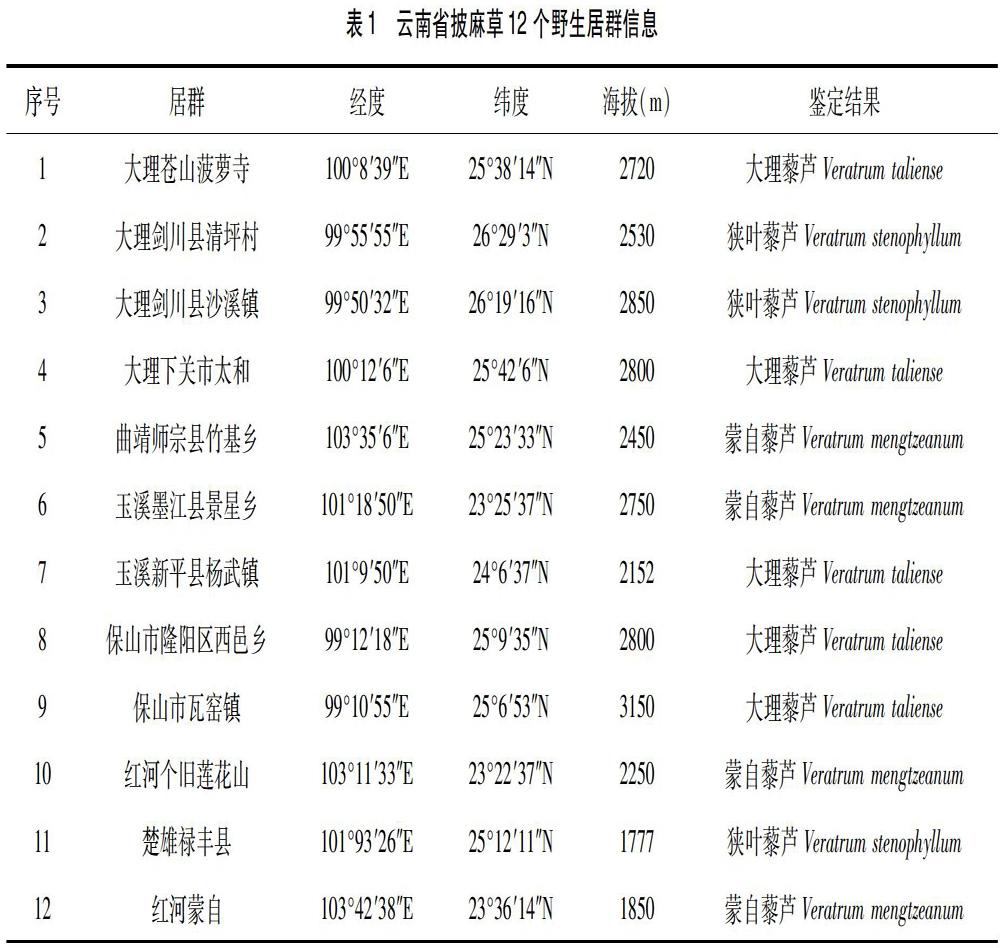
对披麻草资源进行详细的实地调查,选定了云南省 12个野生居群作为研究地,进行群体取样及形态学研究,具体采样居群情况见表1。标本经中国科学院昆明植物所雷立公研究员鉴定。
按照常规统计学方法[10-12]随机选取测量个体,且每个体相距 5 m 以上。每居群 20个个体(部分居群因个体数太少则测了15个),用卷尺、直尺及游标卡尺对其植株叶片长(指离地面1~3cm)、叶片宽(测中间最宽处)、叶片数、叶基长度、叶基直径(测最宽处)等5个性状进行测量,每个性状重复测量3次。在野生居群中选取5个土壤子样(取样深度为0-20cm)作为重复,取完后充分混匀装入自封袋内。土壤样品带回实验室后置于阴凉处自然风干,然后弃去杂物,测量土壤全氮、全磷、全钾、有机质、有效磷和速效钾含量及pH。所得数据采用Excel和SPSS(19.0)软件进行描述性、单因素方差和LSD多重比较处理分析。
2 结果和分析
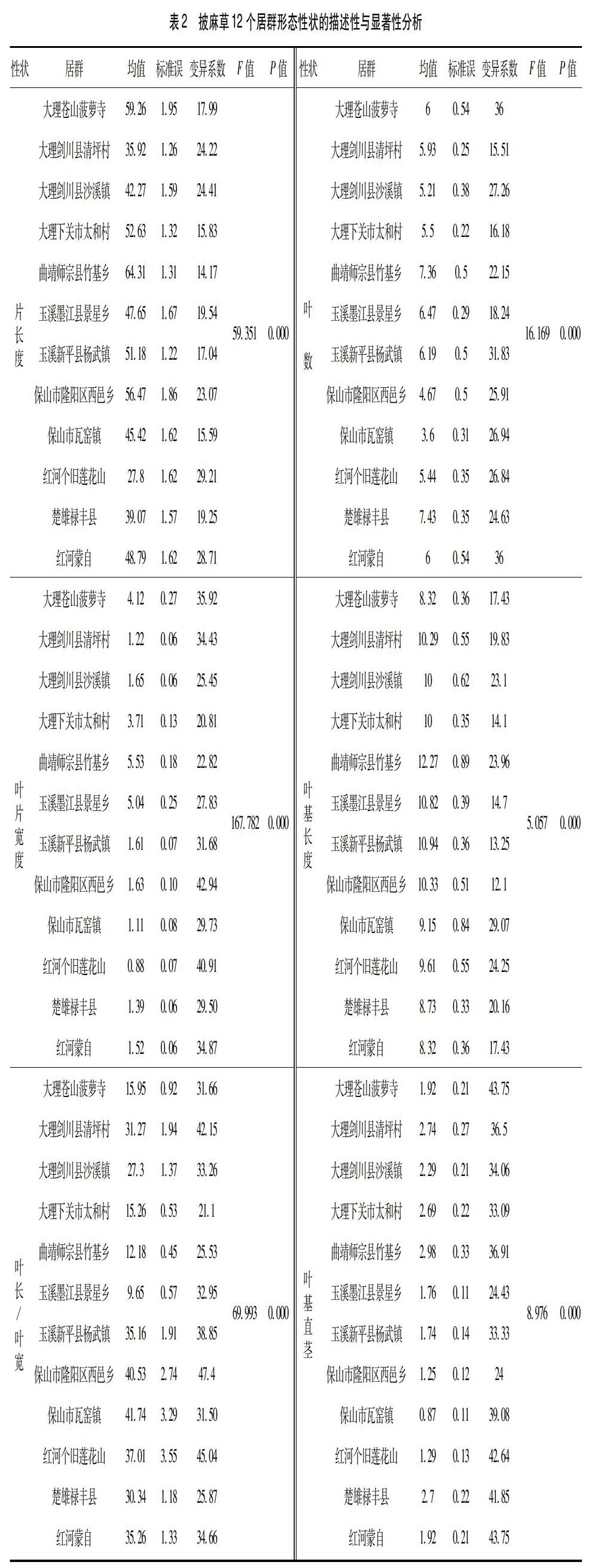
2.1 披麻草野生居群表型多样性 各居群叶片长、叶片宽、叶长/叶宽、叶片数、叶基长度、叶基直径形态特征见表2。以叶片长为例,师宗竹基居群>大理菠萝寺居群>保山西邑居群>大理太和居群>新平杨 武居群>蒙自居群>墨江景星居群>保山瓦窑居群>剑川沙溪居群>楚雄禄丰居群>剑川清坪居群>个旧,分别为(64.31±1.31)cm、(59.26±1.95)cm、(56.47±1.86)cm、(52.63±1.32)cm、(51.18±1.22)cm、(48.79±1.62)cm、(47.65±1.67)cm、(54.42±1.62)cm、(42.27±1.59)cm、(39.07±1.57)cm、(35.92±1.26)cm和(27.8±1.62)cm;变异系数则反映了叶片长的相对变异程度,个旧居群变异系数最大为 29.21,最小的为师宗竹基居群为14.17;单因素方差分析结果显示,披麻草叶片长这一性状在各居群间有极显著性差异(P<0.01).
从表2中可以看出其它5 个性状:叶片宽、叶片长/叶片宽、叶数、叶基长度、叶基直径的各项描述性参数。其中,变异系数最大的为叶基直径;最小的为叶基长度;各形态性状在各居群间均有极显著性差异(P<0.01)。
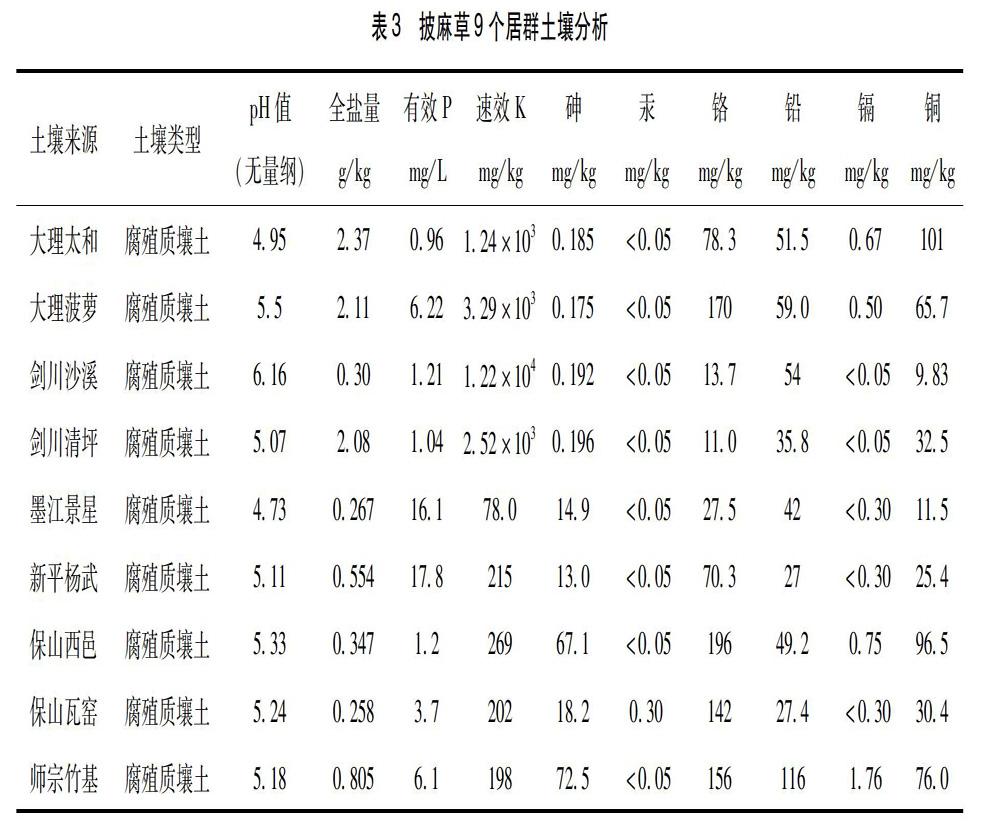
2.2 土壤条件分析 披麻草生长地土壤成分分析结果(表3),从表中可以看出:披麻草生长于呈微酸性的腐殖质壤土中,土壤全盐量值范围为:0.258~2.37 g/kg,保山瓦窑居群最小为0.258 g/kg,大理太和居群的最大为2.37 g/kg;有效磷含量为0.96~17.8 mg/L,大理太和居群含量最小为0.96 mg/L,新平杨武居群含量最大为17.8 mg/L;速效钾含量为78~1.22×104 mg/L,墨江景星居群含量最小为78 mg/L,剑川沙溪居群含量最大为1.22×104 mg/L,速效钾最小值和最大值差异较大。全盐量、有效磷及速效钾含量在9个居群间并无一定的规律可循,因此各居群的植物形态除考虑与生境、气候等相关性因素外,应需考虑土壤与植株各形态相關性的分析研究。
土壤中重金属含量与国家二级标准(土壤环境质量标准GB15618—2008)比较,9个居群砷元素含量除保山西邑居群和师宗竹基居群高于40 ppm(pH<6.5)外,其余居群均低于标准;汞元素含量均低于或等于0.30 ppm(pH<6.5)的标准;铬元素含量均低于250 ppm(pH<6.5)的标准;铅元素含量均低于250 ppm(pH<6.5)的标准;镉元素含量大理太和居群、大理菠萝寺居群、保山西邑居群、师宗竹基居群高于0.30 ppm(pH<6.5),其余居群均低于标准;铜元素含量大理太和居群、大理菠萝寺居群、保山西邑居群、师宗竹基居群高于50 ppm(pH<6.5),其余居群均低于标准。
披麻草生长于微酸性的腐殖质壤土中,土壤中pH值、全盐量值、有效磷值、速效钾值及重金属含量均存在较大的差异。
2.3 生长土壤与植株相关性分析 披麻草药用部位根及根茎的生长状况,难于直观看出,而其每年10月~11月时披麻草地上部分倒苗,于次年春季发芽,其生长状况可反映植株当年的生长状况。因此分析土壤成分与植株特别是地上部分易于观测项目的相关性,希望得出土壤成分与植株形态的相关性,以便于进行栽培管理。土壤成分与植株形态数值(表4)。
用SPSS19.0软件分析全盐量、有效磷和速效钾与植株形态间的相关性。结果如表5显示,各成分与植株形态之间无相关性。因此植株形态特别是地上部分易于观测的项目不能作为全盐量、有效磷和速效钾的直观性指标。
2.4 披麻草野生居群聚类分析 所研究的12个披麻草居群中个旧居群分布于向东的陡坡,大理太和、大理菠萝寺、保山西邑和蒙自居群分布于向西陡坡上,其余7个居群均分布于向西缓坡上。其中剑川清坪居群、师宗竹基居群、保山瓦窑居群分布于林下,师宗居群部分长于石缝中;大理太和居群、大理菠萝寺居群、保山西邑居群、墨江景星居群、个旧居群分布于山坡灌丛;蒙自居群分布于石灰岩山灌丛;其他居群主要分布于在肥沃、疏松、潮湿、排水良好的腐殖质土壤或砂质土壤的山坡草地,且自然植被覆盖度较大。
根据单因素方差和 LSD 多重比较的分析结果,运用 Ward 联接,可以对披麻草大理菠萝寺、剑川清坪、剑川沙溪、大理太和、师宗竹基、墨江景星、新平杨武、保山西邑、保山瓦窑、个旧、楚雄禄丰、蒙自12个居群的形态性状进行聚类分析,结果如图1所示。其中距离较近的居群有:剑川沙溪与墨江景星居群、大理太和与楚雄禄丰居群、新平杨武与保山瓦窑居群、大理菠萝与蒙自居群、剑川清坪与保守西邑居,说明生境较为相似,居群中个体的小环境也很相似,故从形态性状方面来说,两个遗传距离较近的两个居群都分别聚为 1 类;剑川沙溪、墨江景星、师宗竹基、楚雄禄丰和剑川清坪5个居群聚为一支,说明狭叶藜芦和蒙自藜芦分布区的生境差异不是很大;大理菠萝、蒙自、大理太和、保山西邑、个旧5个居群聚为一支,说明大理藜芦和蒙自藜芦部分分布地区的生境很相似。
从披麻草云南12个自然居群的形态变异上来看,居群间的环境差异及个体间的小环境差异对披麻草生长过程中的表型多样性产生了很大的影响。
3 讨论
长期以来,对披麻草的使用,无论是民间还是制药企业完全靠野生资源,加之自然生长缓慢、市场需求量增加,致使其资源量剧减,资源趋向渐危状态。同一居群的披麻草多呈星散残余分布,片段化明显。本文研究结果表明,在所调查的12个披麻草野生居群中,叶片长、叶片宽、叶片长/叶片宽、叶数、叶基长度、叶基直径在居群内和居群间均存在不同程度的变异,主要表现为居群间变异大而居群内个体间变异小,推测生境片段化后,把披麻草原有自然连续分布的居群分割成彼此隔离的小居群后,使片段化居群个体数量减少,增大了片段化居群个体之间近交和自交的概率,各片段化居群间可能发生一些形态上的分化。
形态学性状向来是物种分类、资源鉴定以及探讨物种进化问题的基础[13]。植物的形态性状是由其内在的基因型与所处的外界环境共同作用的结果,因此来自内在和外在因素的综合作用可能是披麻草群间形态性状发生显著差异的主要原因。披麻草表现出来的形态变异,是长期适应不同环境的结果,形态上的多样性和可塑性是披麻草赖以适应和生存的基础。披麻草野生居群的表型多样性丰富,各表型性状在居群间都存在极显著差异,且叶片长、叶片宽、叶片长/叶片宽、叶数、叶基长度、叶基直径等是构成这种差异的主要表型性状。所选居群的地理位置和所选表型性状具有局限性,因此关于披麻草野生居群表型的多样性及土壤因子披麻草表型的综合影响还需进一步研究。
参考文献:
[1]云南省卫生厅.云南省药品标准[M].昆明:云南大学出版社,1998:67-68.
[2]尹子丽,谢晖,张洁.披麻草生物学特性初步研究[J].云南中医中药杂志,2016,37(4):54-57.
[3]尹子丽,杨仙雨,张洁.云南民间药狭叶藜芦生药学研究[J].云南中医中药杂志,2015,36(3):57-59.
[4]尹子丽,谭文红,杨仙雨,等.披麻草三种基原植物的研究進展[J].云南中医中药杂志,2014,35(3):60-62.
[5]LIAO,W J,Yuan,Y M,ZHANG,D Y.Biogeography and evolution of flower color in Veratrum(Melanthiaceae)through inference of a phylogeny based on multiple DNA markers[J].Plant Systematics and Evolution,2007,267:177-190.
[6]尹子丽,张洁,谢晖.药用植物大理藜芦全国适生区的预测分析研究[J].时珍国医国药,2014,25(11):2762-2763.
[7]蒋亚君,申晴,丁西朋,等.柱花草种质资源的表型性状多样性分析[J].草业科学,2017,34(5):1032-1041.
[8]贾丽华,赵长星,王月福,等.不同质地土壤对花生根系生长、分布和产量的影响[J].植物生态学报,2013,37(7):684-690.
[9]阎爱民,陈文新.苜蓿,草木樨,锦鸡儿根瘤菌的表型多样性分析[J].生物多样性,1999,7(2):112-118.
[10]杜荣蹇.生物学统计[M].北京:高等教育出版社,1999:10-23.
[11]虞泓,郑树松,黄瑞复.云南松居群球花多态性研究[J].生物多样性,1998,5(6):267-271.
[12]虞泓,徐正尧,杨彩云.云南松居群花粉形态多样性[J].云南大学学报(自然科学版),1999,21(2):86-89.
[13]Stebbins GL.Relationships between adaptive radiation,speciation and major evolutionary trends[J].Taxon,1971,20(1):3-16.
