眼斑双锯鱼仔稚鱼发育异速生长
2020-08-07静周胜杰杨其彬马振华
胡 静周胜杰 杨 蕊 陈 旭 杨其彬 马振华
(1.中国水产科学研究院南海水产研究所,热带水产研究开发中心,三亚 572018;2.农业部南海渔业资源开发利用重点实验室,广州 510300;3.三亚热带水产研究院,三亚 572018)
眼斑双锯鱼(Amphiprion ocellaris),俗称公子小丑,隶属于雀鲷科(Pomacentridae)、海葵鱼亚科(Amphiprioninae)、双锯鱼属(Amphiprion),广泛分布于印度—西太平洋地区,从印度洋东部延伸到印度尼西亚和菲律宾群岛,从澳大利亚西北部海岸延伸到日本琉球群岛[1]。20世纪50年代,眼斑双锯鱼的人工规模化繁育即已取得成功[2],近年来随着电影《海底总动员》的播出,以眼斑双锯鱼为代表的小丑鱼家族知名度急剧上升,市场价值翻倍增长,供不应求[3]。为降低成本、提高养殖效率,创造更多的价值,近几十年不少学者开展了小丑鱼的各项研究,内容囊括早期胚胎发育、营养学、病害学及遗传生物学等[4—7]。然而,在鱼类早期发育过程中个体器官受环境因素影响而体现出不同的发育速率,进而影响仔、稚鱼的运动、摄食及存活生长等生理生态学活动,因此,研究仔鱼发育对了解个体早期生活史、指导生产环境调控具有重要意义,目前未见对眼斑双锯鱼发育异速生长的研究。
异速生长又称相对生长,普遍存在于生物界,是动植物适应外界环境而产生的机体各个功能器官发育不同步的现象[8]。异速生长概念最早由Snell[9]提出,后D’arcy[10]、Huxley等[11]分别对其进行了阐述。Huxley等[11]在研究招潮蟹蟹钳生长发育时首次提出了量化异速生长指数的“简单异速方程”,即幂函数y=axb(a为截距,b为异速生长指数),该方程广泛运用于机体异速生长研究,是目前分析研究鱼类早期发育生长模式最常用手段[12]。有关鱼类异速生长的研究,国内外相关报道很多,如云纹石斑鱼(Epinephelus moara)[13]、金眼短鲷(Nannacara anomala)[14]、牙鲆(Paralichthys olivaceus)[15]、红鳍笛鲷(Lutjanus erythropterus)[16]及大鳞副泥鳅(Paramisgurnus dabryanus)[17]等。研究发现,鱼类早期发育各个功能器官的生长发育具有一定规律性[18—20],即个体早期优先发育与生命活动密切相关的器官,体现为正异速生长,后为保证以最小的能量获得最优的生存能力而转为等速或慢速生长[20—23]。因此,通过对眼斑双锯鱼仔、稚鱼异速生长的研究,有助于了解其早期发育阶段对环境的偏好及重要功能器官的发育特性,相关研究对育苗工作具有重要的实践意义。
1 材料与方法
1.1 实验材料
眼斑双锯鱼苗种由中国水产科学研究院南海水产研究所热带水产研究开发中心人工繁育,来自同一对亲鱼同一批受精卵,亲本为野生捕捞后、人工驯养2年个体,受精卵孵化日期为2018年5月16日。以2尾/L密度养殖于300 L水体玻璃钢桶,水质参数: 溶氧量≥6.5 mg/L,盐度为32±2,水温为(28±1)℃,pH 8.0—8.2,亚硝酸盐含量小于0.03 mg/L,氨氮含量小于 0.01 mg/L,自然光照。
1日龄(2.97±0.12) mm开始投喂轮虫(Brachionus rotundiformis),密度保持在10—20个/mL直至5日龄(4.36±0.27) mm,于3日龄(3.75±0.25) mm混合投喂丰年虫,轮虫密度不变,丰年虫密度范围保持在20—30个/mL直至9日龄(5.52±0.22) mm。之后仅投喂丰年虫,密度保持在50—60个/mL直至实验结束。
1.2 研究方法
使用体视显微镜(XTL-165-VT-45X系列连续变倍体视显微镜,凤凰光学股份有限公司)进行观察、拍照,后使用配套软件ToupCam以平行或垂直于鱼体水平轴为基准进行长度测量(精确度0.01 mm)。初孵仔鱼1日龄开始每隔一天取样至29日龄,取样时间为每日上午10:00,每次取样10尾,用25 mg/L浓度的丁香酚 (New Zealand Ltd)麻醉后,在体视显微镜下观察其外部各个器官的形态变化并拍照,剔除畸形苗、保留骨骼健康个体待数据统计。测量指标包括头部部分(吻长、吻至鳃裂前缘长、眼径、眼间距、口宽、头高)、身体部分(头长、体高、躯干长、尾长、尾柄长、尾柄高、体厚)及游泳器官(尾鳍、背鳍、胸鳍、腹鳍和臀鳍)(主要测量参数直观图如图1)。
1.3 数据处理
全长、体质量与日龄的关系采用Brody’s生长方程拟合:y=aebx来表达,x为日龄,y为相应日龄的全长、体质量,a为常数,b为Brody’s系数。鱼类异速生长模型以Huxley等[11]提出的方程y=axb来构建,其中b为异速生长指数,b=1为等速生长,b>1为快速增长(正异速生长);b<1为慢速增长(负异速生长)[24]。仔稚鱼的划分及生长拐点的确定分别按照Stroband等[25]和van Snik等[26]的方法进行,均得出11日龄为临界点,以此将眼斑双锯鱼仔、稚鱼生长曲线方程定为应用SPSS 19.0统计软件对模型进行非线性回归参数拟合,并应用EXCEL 2016进行分段回归模型拟合,以决定系数R2的最大值和残差平方和的最小值为曲线拟合标准,运用SPSS 19.0统计软件,采用单因素方差分析方法(ANOVA)进行显著性检测。
2 结果
2.1 眼斑双锯鱼仔稚鱼全长、体质量与日龄的关系
眼斑双锯鱼仔稚鱼全长、体质量与日龄的关系指数方程分别为y=2.97e0.06x(R2=0.957)、y=0.001e0.16x(R2=0.952),拟合可靠性均较高(图2和图3)。眼斑双锯鱼初孵仔鱼全长为(2.86±0.00) mm,经过30d生长,全长达到(16.31±1.08) mm,平均增长率为0.448 mm/d;眼斑双锯鱼初孵仔鱼体质量为(0.697±0.00) mg,经过30d生长,体质量达到(86.1±5.69) mg,平均增重率为2.85 mg/d。
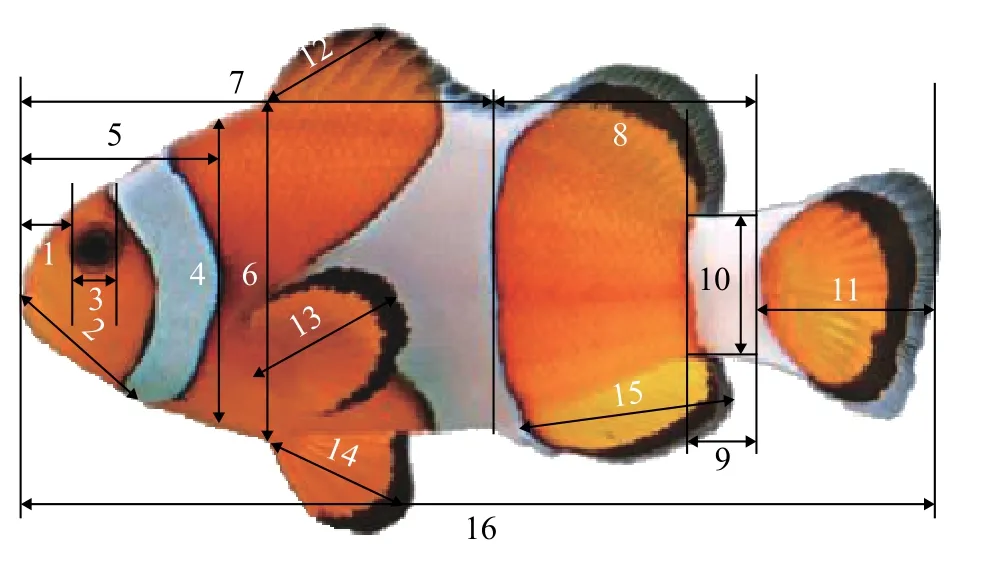
图1 眼斑双锯鱼仔稚鱼形态学量度指标Fig.1 Measurements of morphometrics of A. ocellaris larvae and juveniles

图2 眼斑双锯鱼仔、稚鱼日龄与全长的关系(从1日龄至29日龄)Fig.2 Total length of A. ocellaris larvae and juveniles from 1 to 29 days post-hatching
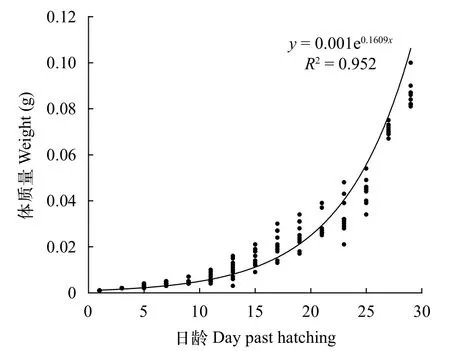
图3 眼斑双锯鱼仔、稚鱼日龄与体质量的关系(从1日龄至29日龄)Fig.3 Body weight of A. ocellaris larvae and juveniles from 1 to 29 days post-hatching
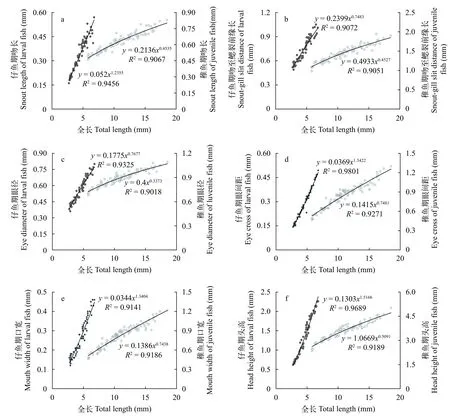
图4 眼斑双锯鱼仔、稚鱼头部各器官异数生长曲线Fig.4 Allometric growth curve of head organs in larval and juvenile A. ocellaris
2.2 眼斑双锯鱼仔、稚鱼发育过程中各器官的异速生长
头部各器官异速生长眼斑双锯鱼的吻长
(图4a)、吻至鳃裂前缘长(图4b)、眼径(图4c)、眼间距(图4d)、口宽(图4e)和头高(图4f)在仔、稚鱼期均表现出了异速生长。异速生长指数b1分别为1.236、0.748、0.768、1.342、1.340和1.517,6个异速生长指数与1均具有显著性差异(P<0.05),即吻长、眼间距、口宽和头高表现为正异速生长,吻至鳃裂前缘长和眼径表现为负异速生长;b2分别为0.454、0.453、0.337、0.748、0.744和0.509,各异数生长指数均具有显著性差异(P<0.05),即头部各器官于稚鱼期均表现为负异速生长。故在11日龄前,眼斑双锯鱼仔鱼的头部器官中吻长、眼间距、口宽和头高均为快速生长,11日龄后,头部各器官均为慢速生长。在11日龄时,眼斑双锯鱼仔鱼全长为(6.12±0.63)mm,此时吻、眼间距、口宽和头高均存在生长拐点(P<0.05)。
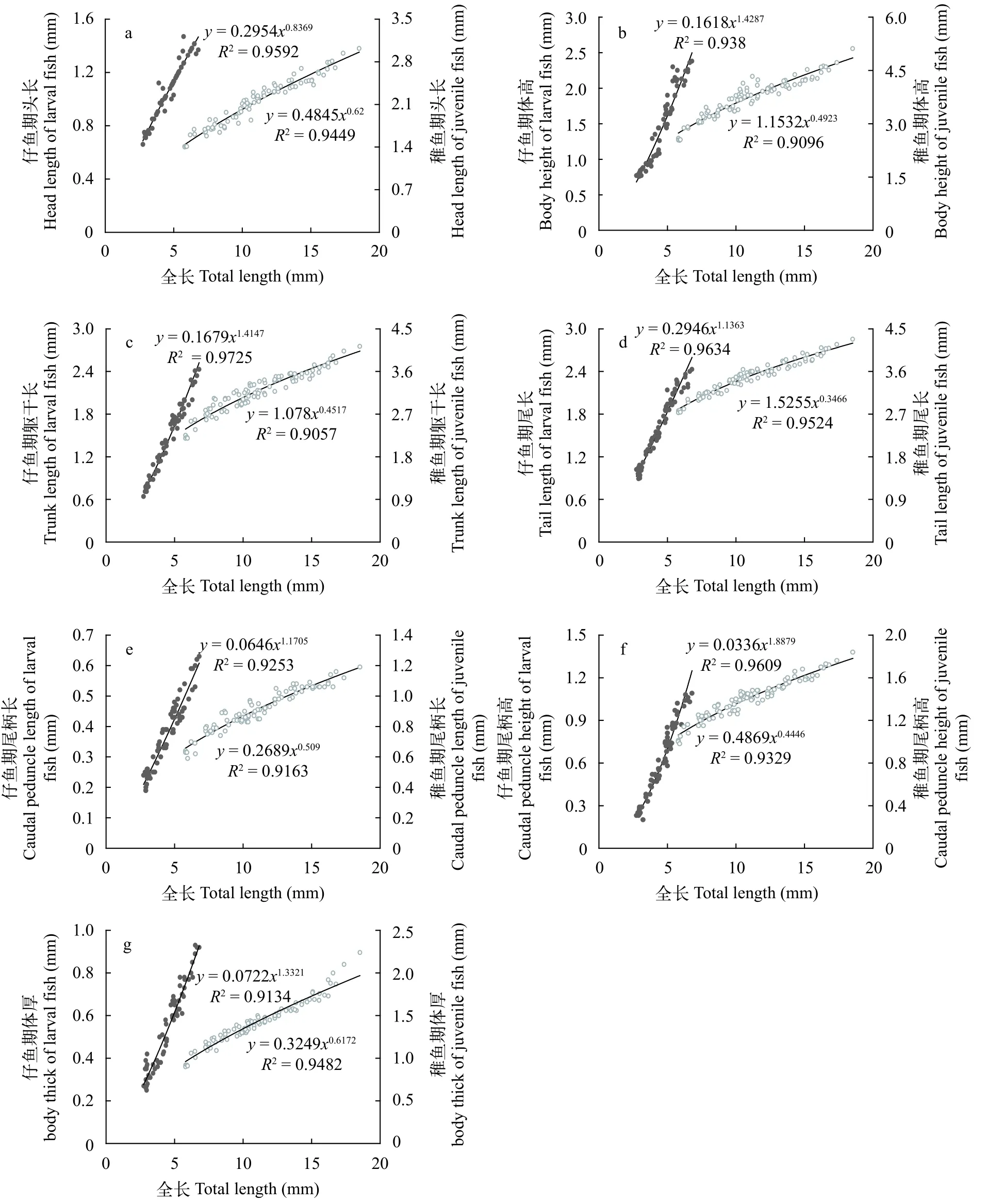
图5 眼斑双锯鱼仔、稚鱼身体部分异数生长曲线Fig.5 Allometric growth curve of body parts in larval and juvenile A. ocellaris
身体部分的异速生长眼斑双锯鱼的头长(图5a)、体高(图5b)、躯干长(图5c)、尾长(图5d)、尾柄长(图5e)、尾柄高(图5f)和体厚(图5g)在仔、稚鱼期均表现出了异速生长。头长异速生长指数b1为0.837(P<0.05),表现为负异速生长,其他部分的异数生长指数分别为1.429、1.415、1.136、1.171、1.888和1.332,与1差异显著(P<0.05),表现为正异速生长。稚鱼期各部分的异速生长指数b2分别为0.620、0.492、0.452、0.347、0.509、0.445和0.617,与1之间存在显著性差异(P<0.05),均表现为负异数生长。这说明在身体部分中,眼斑双锯鱼头长在仔、稚鱼期均为慢速生长,其他仅在仔鱼期表现为快速生长,稚鱼期均为慢速生长。体高、躯干长、尾长、尾柄长、尾柄高和体厚在11日龄时存在生长拐点(P<0.05)。
游泳器官异速生长眼斑双锯鱼的外部游泳尾鳍(图6a)、背鳍(图6b)、胸鳍(图6c)、腹鳍(图6d)和臀鳍(图6e),在仔、稚鱼期均表现出了异速生长。仔鱼期的尾鳍、背鳍、胸鳍、腹鳍和臀鳍的异速生长指数b1分别为1.887、2.466、3.406、2.679和2.408,与1均存在差异显著(P<0.05),均表现为正异速生长。稚鱼期各鳍条异速生长指数b2分别为0.395、0.856、0.822、0.622和0.724,与1差异显著(P<0.05),均表现为负异速生长。眼斑双锯鱼的游泳器官在仔、稚鱼的发育存在显著差异,各鳍条于仔鱼期均快速生长,稚鱼期均慢速增长,且各鳍条在11日龄时均存在生长拐点(P<0.05)。

图6 眼斑双锯鱼仔、稚鱼游泳器官异数生长曲线Fig.6 Allometric growth curve of swimming organs in larval and juvenile A. ocellaris
3 讨论
鱼类发育的早期阶段是育苗成功的关键时期,常因开口饵料、机械冲击、捕食者及各种环境因素而产生较高死亡率[27—29],因此,在人工养殖条件下,探究鱼类的发育模式,以此寻找合适的饲养环境,使两者相协调,对提高存活率、确保育苗成功至关重要[30,31]。鱼类早期发育过程极其重要而复杂,各器官经历剧烈、迅速的形态变化,以适应环境条件变化和摄食、避敌等生理活动需求[32,33],包括捕食、呼吸和运动器官等[19,34]。鱼类的异速生长现象是一种自我保护机制,体现为仔鱼孵化后,对早期存活发挥积极作用的重要功能器官优先发育,表现出快速生长。与本研究结果一致,同时也被大量研究证实[20,35,36]。
3.1 头部器官的发育
水产动物包括鱼类的头部发育与个体视觉、神经及信息处理等的功能行使有关,是摄食、游泳器官发育的基础[37],眼斑双锯鱼短期内头部(头高)快速生长,为其他器官的发育提供了可能性[38],吻、眼间距及口宽仔鱼期的快速生长印证了该结论。仔、稚鱼摄食能力的强弱与口、吻的发育和完善程度密切相关,眼斑双锯鱼仔鱼的卵黄囊于孵化当天即消耗殆尽,故需立即进行开口饵料的投喂,避免死亡率的迅速增高。吻、眼间距和口宽在11日龄时出现生长拐点,即眼斑双锯鱼在开口后短期内以上摄食相关器官需继续发育,以适应不同食物大小、提高摄食能力、降低死亡率[39]。
一般来说,由于视觉与个体摄食、避敌间显著的相关性,视觉器官在大多数鱼类早期发育中应均为正异速生长[16,38,40—42],稚鱼期均为慢速生长,与席丹等[43]对许氏平鲉(Sebastes schlegelii)的研究结果相似,孵化初期即进入慢速生长阶段,这表明眼斑双锯鱼胚胎发育的器官形成期,其眼部发育即已基本完成[44,45],具备完善摄食、避敌功能,胚后的慢速发育为其他器官发育提供了更多能量与空间。不同物种眼部发育快慢的差异,可能与其眼睛发育史差异有关[34]。许氏平鲉为卵胎生生殖,其眼球可能在亲本体内已发育完善,孵化初期既体现为慢速生长,眼斑双锯鱼孵化周期长,胚胎期眼睛即已得到良好发育。眼斑双锯鱼仔、稚鱼的吻、眼间距、口宽和头高在11日龄时均出现拐点,表明了吻、眼间距、口宽和头高的生长已完成了优先发育,进入次要发育阶段。
3.2 身体部分的发育
研究表明,大部分鱼类身体部分的早期模式为头长、尾长呈正异速生长而躯干长呈负异数生长,如西伯利亚鲟(Acipenser baerii)[46]、鲈鲤(Percocypris pingi)[47]、赤眼鳟(Spualiobarbus curriculus)[48]、珍珠龙胆石斑鱼(E. fuscoguttatus♀×E. lanceolatus♂)[49]及卵形鲳鲹(Trachinotus ovatus)[40]等,与此不同的是,本研究中眼斑双锯鱼拐点前头长为负异速生长、躯干长为正异数生长,其身体部分这种异数生长模式与许氏平鲉[43]相似,对比两者生殖方式发现,许氏平鲉为卵胎生、在亲鱼体内停留时间较长,而眼斑双锯鱼孵化周期长(7—10d)、孵化过程亲本具护卵行为,推测在此过程中,两者头部已得到较好发育,而两者头高与头长的异数生长情况相反,这可能与物种间外部形态差异有关[50]。多数鱼类躯干部的负异速生长显示了对动力学和低能耗的适应,如鮸(Miichthys miiuy)[51]、西伯利亚鲟[46]及赤眼鳟[48]等,该生长特征有效的减少了头部和尾部距离,而减少了摄食运动负担、促进了身体平衡。在该研究中,眼斑双锯鱼躯干长、体高和体厚均为快速生长,与加利福尼亚牙鲆(P. californicus)[52]结果相似。这和消化系统的发育及卵黄囊消失时间相关,消化道占据体腔绝大多数空间,使得立体结构纵、横向拉升,确保0日龄卵黄囊即消失的眼斑双锯鱼个体具备足够的消化外源性食物的能力[16]。眼斑双锯鱼尾长、尾柄长和尾柄高于仔鱼期均处于快速生长状态,一方面,尾部的快速发育为重要游泳器官尾鳍的发育提供了空间,为捕食动力提供了组织基础[46],另一方面,使得眼斑双锯鱼的后部体型婀娜、摇曳生姿,奠定了其观赏价值高、经济效益显著的基调[50]。
3.3 游泳器官的发育
眼斑双锯鱼仔、稚鱼尾鳍、背鳍、胸鳍、腹鳍和臀鳍均于11日龄出现生长拐点,且11日龄前均处于快速生长,11日龄后为慢速生长,与多数鱼类游泳器官发育模式相似,如红鳍笛鲷、鲈鲤及赤眼鳟[16,47,48]等,拐点前各鳍条生长迅速,而拐点后显著减慢,这表明眼斑双锯鱼的各外部游泳器官于稚鱼期均已完成了快速生长,能够满足眼斑双锯鱼的快速游泳能力,之后的慢速生长为机体其他部位的发育提供了更多能量[38]。与多数鱼类孵化后游泳能力弱不同的是,在实验观察中发现,眼斑双锯鱼孵化后活动较强,受触动刺激后游散迅速,聚光效果明显,这可能与其发达的鳍条尤其是尾鳍优势发育有关,是适应环境的结果[53]。孵化时,眼斑双锯鱼个体卵黄囊即已消耗殆尽,发达的外部游泳器官为外源营养的摄取提供了足够前进动力,对适应外界环境起着积极作用。通过眼斑双锯鱼主要游泳器官(背鳍、胸鳍、腹鳍、臀鳍和尾鳍)的全面分析发现,眼斑双锯鱼孵化初期既具备较强游泳能力,为提升摄食、避敌能力短期内继续快速发育,11日龄游泳器官已经基本完善,运动能力大幅度增加,而美洲鲥(Alosa sapidissima)、许氏平鲉、鲈鲤及珍珠龙胆石斑鱼[34,43,47,49]等分别需13—44d、12—46d、24—4d5及14—23d才进入慢速发育阶段,从发育时间来看,眼斑双锯鱼游泳器官具备显著发育优势。这充分证明了游泳器官在仔鱼早期生长发育中的重要地位,游泳能力的显著提升与对外源营养的需求呈正相关,对自身捕食能力大有裨益。
在外部游泳器官中,尾鳍在鱼类早期发育中最为重要,该结论在多种鲷科鱼类[54—57]的研究中得以体现,在本研究中亦得到相似结果,尾鳍较其他鳍条优先发育,各鳍条初始长度分别为: (0.45±0.03、0.02±0.00、(0.98±0.01)×10-2、0.02±0.00和0.03±0.00) mm,平均增长速度分别为: (6.857、0.964、0.865、0.893、1.143)×10-2mm/d。
本研究对眼斑双锯鱼仔、稚鱼的异速生长特征进行了较为详细的叙述,亦初步探讨了异速生长对其体型的影响,研究表明,眼斑双锯鱼头部、躯干及游泳器官均呈现出异速生长,并且根据其生理、繁殖特性及各器官功能差异,既存在与其他鱼类相似的生长模式,又存在自身独特模式,从而根据自身情况选择合适的生长发育模式,以提高自身主动摄食、躲避敌害等能力。有关眼斑双锯鱼异速生长模式的研究,不仅有助于人为营造合适生长环境、提供适宜饵料,最大限度提高存活率、摄食率,还能在形态上进行人为筛选,提供优质鱼苗,为眼斑双锯鱼高效养殖提供理论依据。
