STAT5与肿瘤关系的研究进展
2020-08-04赵玉张俊伶冯国兴樊赛军
赵玉,张俊伶,冯国兴,樊赛军
信号转导和转录激活因子(signal transducersand activators of transcription,STAT)家族有7 个成员,包括STAT1~4、STAT5A、STAT5B 和STAT6。其 中,STAT5A 和STAT5B 是STAT5 的两个高度同源的亚型,在人体各个组织器官中广泛表达[1]。除了参与调控细胞的增殖、迁移和凋亡,STAT5 在尿囊素、脂肪酸、肌酐等多种物质的代谢以及胚胎发育、免疫调节、造血过程中也发挥重要作用[2-4]。STAT5信号异常与肺癌、甲状腺癌、乳腺癌等多种常见癌症的发生发展密切相关。有研究表明,前列腺癌组织中活化STAT5 水平显著高于正常组织[5],提示STAT5 具有促进肿瘤生长的作用;而在乳腺癌晚期,STAT5活化水平升高则肿瘤侵袭能力减弱[6],提示STAT5 亦具有肿瘤抑制的作用。靶向抑制STAT5可以提高肿瘤放疗、化疗及免疫治疗效果,这为肿瘤治疗提供了新策略。目前有关STAT5在不同肿瘤中的作用差异及其靶向治疗策略均未明确。鉴于此,本文对STAT5在几种常见人类肿瘤中的作用及相关机制进行总结,并对STAT5靶向治疗策略进行探讨,为STAT5相关肿瘤的临床治疗提供依据。
1 STAT5概述
STAT5 包括两种由人染色体17q21.2 上串联的两个基因编码的亚型,即STAT5A 和STAT5B,两者同源性高达96%[7]。STAT5 蛋白具有6 个功能性结构域:N 端相互作用结构域、卷曲螺旋结构域、DNA结合结构域、连接区、Src 同源2 结构域(Src homology 2 domain,SH2)和转录激活结构域。STAT5 的活化主要通过其自身酪氨酸的磷酸化完成,正常生理状态下,细胞内STAT5磷酸化的活化状态与去磷酸化的失活状态保持动态平衡,当这种平衡被打破时,会导致多种人类恶性肿瘤的发生发展[7-8]。
2 STAT5在不同类型肿瘤中的作用
STAT5 在肿瘤中具有双重作用。一方面,STAT5 既可通过直接调控与细胞增殖、凋亡相关的靶基因表达,影响肿瘤发生发展;也可通过诱导线粒体功能紊乱,增加细胞内活性氧(reactive oxygen species,ROS)水平,诱导DNA损伤,从而促进肿瘤进展。另一方面,STAT5 在特定微环境下可通过抑制肿瘤转移而发挥抑癌作用。
2.1 前列腺癌 95%的激素难治性前列腺癌及61%的远处转移前列腺癌与STAT5 持续活化相关[9-10]。研究表明,前列腺癌中STAT5 的活化与非受体酪氨酸蛋白激酶Janus 激酶2(Janus kinase 2,JAK2)介导的催乳素(prolactin,PRL)及生长激素(growth hormone,GH)、白细胞介素(interleukin,IL)-6 信号异常相关[11-13]。抑制STAT5 活化可下调前列腺癌细胞中抗凋亡蛋白B 淋巴细胞瘤2(B-cell lymphoma-2,Bcl-XL)和细胞周期素D1(cyclin-D1)的表达水平,进而诱导前列腺癌细胞凋亡,从而抑制肿瘤生长[14]。有研究显示,采用siRNA干扰STAT5B可以显著降低前列腺癌细胞系DU145 的存活率,而干扰STAT5A对前列腺癌细胞系DU145的存活率无显著影响;但在体内实验中,STAT5A/B 敲降组小鼠的肿瘤体积明显小于STAT5B 单独敲降组,提示STAT5A、STAT5B 均可促进前列腺癌细胞增殖,STAT5B 对前列腺癌细胞生长的促进作用是否强于STAT5A 有待进一步研究验证[15]。另有研究发现,STAT5A/B高表达的前列腺癌容易发生转移[16]。
2.2 乳腺癌 在乳腺癌发生发展的不同时期,STAT5发挥从促癌到抑癌的不同作用。在乳腺癌早期,STAT5 促进乳腺肿瘤的生长及恶性转化。在乳腺癌中,GH[17]和PRL[18]是参与STAT5过度活化过程的重要因子。就PRL-STAT5信号轴而言,有研究证实,带状疱疹透明带样结构域蛋白1(CUB and zona pellucida-like domain-containing protein 1,CUZD1)是影响PRL介导小鼠乳腺上皮细胞STAT5活化的关键因子,在乳腺癌细胞系MCF7 细胞中过表达的CUZD1 会促进STAT5 活化,从而诱导细胞恶性增殖[19]。在乳腺癌晚期,活化STAT5 能够有效抑制病灶扩散,延缓肿瘤的侵袭和转移进程[6,14,20]。Lee等[21]研究表明,活化STAT5 可以通过促进自然杀伤细胞(nature killer cell,NK)成熟,增强NK 细胞的抗肿瘤能力,抑制乳腺癌细胞MDA-MB-231的增殖。
2.3 黑色素瘤 黑色素瘤是一种高发病率、高病死率的皮肤恶性肿瘤,62%的黑素瘤患者中活化STAT5 水平较正常组织升高[22]。有研究表明,STAT5参与黑色素瘤细胞的增殖、侵袭,影响患者化疗、免疫治疗效果[23]。酪氨酸蛋白激酶Src 和JAK1介导人恶性黑色素瘤中STAT5的活化[24]。抑制黑色素瘤细胞系A375细胞中STAT5的活化,可显著下调抗凋亡蛋白Bcl-xL 的表达,从而促进黑色素瘤细胞凋亡[25]。Wellbrock 等[26]研究发现,STAT5在干扰素α(interferon α,IFNα)耐受的黑色素瘤细胞中高表达,抑制STAT5表达可恢复黑色素瘤细胞对IFNα治疗的敏感性。Ohanna等[27]研究发现,烟酰胺磷酸核糖基转移酶(nicotinamide phosphoribosyltransferase,NAMPT)可促进黑色素瘤细胞增殖、转移和侵袭,使黑色素瘤产生免疫治疗抗性,而抑制STAT5 活化可以降低NAMPT 的表达,进而抑制黑素瘤细胞增殖。以上研究提示STAT5 可以作为黑色素瘤治疗的靶标。
2.4 血液系统恶性肿瘤 正常情况下STAT5 可调节造血细胞的增殖和分化,而过度活化的STAT5 会导致骨髓细胞的恶性转化。研究发现,慢性髓系白血病(chronic myeloid leukemia,CML)、红系白血病及急性淋巴细胞白血病(acute lymphoblastic leukemia,ALL)等患者体内存在持续性活化的STAT5[28-30]。
具有高度酪氨酸激酶活性的BCR-ABL 融合基因是一种抗细胞凋亡基因,其突变会导致CML的发生。研究表明,STAT5 表达上调会增加CML 细胞DNA 双链断裂率,进而增加BCR-ABL 突变率;ROS亦可增加BCR-ABL 突变率[28,31],并且ROS 与STAT5间存在正反馈调节效应。ROS能够诱导酪氨酸磷酸酶失活,提高STAT5的活化水平,而活化的STAT5会抑制抗氧化酶的合成,增强烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)氧化酶的转录,促进细胞分泌更多的ROS,进一步增加BCR-ABL 突变率,进而加速白血病进程[32]。另有研究表明,活化的STAT5 可通过乳腺癌易感基因1(BRCA1)下调人红系白血病细胞TF-1细胞中抑癌基因p53 的表达水平,从而发挥促癌作用[33]。Katerndahl 等[34]通过染色质免疫共沉淀和二代测序技术发现,原本在正常B细胞分化过程中,受核因子κB(nuclear factor kappa-B,NF-κB)相关肿瘤抑制通路调节的基因,与STAT5结合后,会促进B细胞祖细胞的增殖,从而引发B细胞急性淋巴白血病;且STAT5表达水平越高,患者预后越差。
2.5 免疫系统相关肿瘤 免疫系统相关肿瘤的发生与STAT5 的功能性缺失有关。STAT5 是NK 细胞发挥免疫功能的主要调节因子,是其从发挥免疫监视作用到促肿瘤作用的转换分子,见图1。正常情况下,STAT5 可促进NK 细胞的成熟,使其发挥抗肿瘤作用。STAT5缺陷的NK细胞分化异常,不能分化为成熟NK 细胞。未成熟NK 细胞血管内皮生长因子(vascular endothelial growth factor,VEGF)-A 分泌水平升高,VEGF-A 可通过增强血管生成能力促进肿瘤生长[35]。已有研究在非小细胞肺癌(non-small cell lung cancer,NSCLC)、乳腺肿瘤和结肠肿瘤患者中检测到高VEGF-A 分泌水平的非成熟NK 细胞[36-37],在这种情况下使用STAT5 抑制剂进行肿瘤治疗,不仅会干扰NK 细胞的免疫功能,还可能诱导VEGF-A 表达,促进肿瘤进展。Zhang 等[38]研究表明,STAT5的功能性缺失会干扰淋巴细胞的分化,导致严重的免疫缺陷,使肿瘤发生风险大大增加。
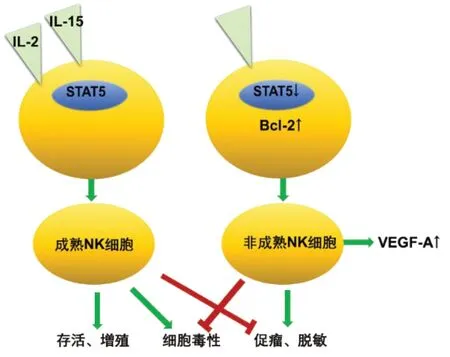
Fig.1 STAT5 is a regulator in the maturation of NK cells图1 STAT5是NK细胞成熟的调节因子
2.6 肝细胞癌(hepatocellular carcinoma,HCC) 在HCC的发生发展中,STAT5既有抑癌作用,也有促癌作用。正常情况下,STAT5 可激活肝细胞中细胞周期蛋白依赖性激酶抑制剂(cyclin dependent kinase inhibitor,CDKN)1a 和CDKN2b 的表达,对细胞增殖产生负调控作用[39]。STAT5缺失会使肝细胞增殖能力增强,进而引发脂肪变性、肝纤维化,促进肝癌进程[40-41]。另有研究表明,环状RNA circScd1 通过JAK2/STAT5 通路促进脂肪肝的发生[42]。胰岛素样生长因子1(insulin-like growth factor 1,IGF-1)可通过STAT5B 诱导上皮-间充质转化(epithelialmesenchymal transition,EMT),增强HCC 细胞的侵袭性[43]。
2.7 NSCLC 多项研究显示STAT5 在NSCLC 中高表达,可能具有促进NSCLC 发生发展的作用。Sánchez-Ceja 等[44]对92 例NSCLC 标本进行研究发现,肺癌的组织学类型与细胞核中STAT5 和抗凋亡蛋白Bcl-XL的表达相关,提示STAT5可能通过抑制细胞凋亡促进NSCLC 的进展。 Pastuszak-Lewandoska 等[45]研 究 发 现,STAT5A、STAT5B 在NSCLC 癌组织中的表达水平较癌旁组织分别增高69%和79%,提示STAT5A、STAT5B 在NSCLC 中均具有促癌作用,STAT5B 的促癌作用可能强于STAT5A。Zhou 等[46]研究发现,NSCLC 患者的晚期病理分期、淋巴结转移和总生存期皆与细胞因子信号转导抑制因子2(suppressor of cytokine signaling 2,SOCS2)表达下调有关,而SOCS2 可与IGF-1 受体竞争性结合STAT5,导致STAT5失活,NSCLC细胞的迁移和侵袭能力受到抑制,进一步提示STAT5 在NSCLC 中的具有促癌作用。然而Yang 等[47]研究发现,STAT5 表达上调可延长患者的总生存期。以上研究提示STAT5 在NSCLC 中可能具有促癌与抑癌的双重作用,其相关机制有待进一步研究。
2.8 其他肿瘤 Liang 等[48]研究表明,STAT5B 可通过调控抗凋亡蛋白B 淋巴细胞瘤-2(B-cell lymphoma-2,Bcl-2)、细胞周期蛋白依赖性激酶抑制剂家族蛋白p21、周期蛋白依赖性激酶p27、酪氨酸激酶局部黏着斑激酶(focal adhesion kinase,FAK)和VEGF,参与人类多形性胶质母细胞瘤(glioblastoma,GBM)的生长、侵袭和迁移,进而促进GBM进展。在结肠癌(colorectal cancer,CRC)中,STAT5 可通过上调ROS 水平,诱导人CRC 细胞G1期细胞周期阻滞,抑制肿瘤细胞增殖,促进CRC 细胞凋亡[49-50]。Miao等[51]研究发现,鼻咽癌(nasopharyngeal cancer,NPC)组织中组蛋白乙酰转移酶1(histone acetyltransferase 1,HAT1)高表达;而在NPC 细胞系HNE2 细胞过表达HAT1 后,STAT5 活性和抗凋亡基因Bcl2L12 表达水平显著增加,细胞凋亡比例降低;进一步分析发现HAT1 直接参与STAT5 与Bcl2L12 启动子的结合,STAT5 可通过HAT1-STAT5-Bcl2L12 促进NPC 进展。Zeng 等[52]通过基因组富集分析发现,IL2-STAT5 信号通路参与COL5A2 促进膀胱癌(bladder cancer,BC)细胞增殖。另有研究表明,STAT5 通过调控上皮型钙黏蛋白、波形蛋白的表达,诱导EMT,促进甲状腺癌的侵袭、转移;JAK2-STAT5有促进骨髓纤维化的作用[53]。人T淋巴细胞病毒-I(human T cell lymphotropic virus I,HTLV-I)编码的p12(I)蛋白可通过增强STAT5 转录活性,引发成人T 细胞白血病和热带痉挛性截瘫[54]。人类免疫缺陷病毒(human immunodeficiency virus,HIV)感染导致的免疫缺陷也与STAT5表达下调相关[55-56]。
3 STAT5相关肿瘤治疗策略
3.1 STAT5靶向治疗 STAT5相关肿瘤的靶向治疗策略主要是直接或间接抑制STAT5 活性。理论上,直接靶向STAT5可以通过干扰STAT5的表达、活化,及干扰STAT5 与靶基因的结合实现。AC-4-130 是一个靶向STAT5 SH2 结构域的小分子抑制剂,通过直接与STAT5结合,干扰STAT5的活化,影响STAT5与靶基因的结合,抑制人急性髓系白血病(AML)细胞增殖[30]。在乳腺癌细胞中,采用短发卡RNA(short hairpin RNA,shRNA)直接下调STAT5B 表达,可以抑制乳腺癌细胞的侵袭能力[57]。采用匹莫齐特选择性抑制STAT5活化,可以抑制CUZD1驱动的乳腺癌细胞增殖,对乳腺癌起到治疗作用[19]。
间接抑制STAT5 活性的方法包括:通过靶向STAT5 相关激酶,抑制STAT5 信号传导;通过增强STAT5 负反馈调节通路活性,抑制STAT5 活性。去乙酰化酶抑制剂(如曲古抑菌素A、辛二酰苯胺异羟肟酸和丁酸钠)可特异性阻断STAT5信号通路,下调STAT5靶基因C-Myc的表达[58]。SOCS2重组蛋白通过与生长激素受体相互作用,下调GH-STAT5 信号下游靶基因,抑制生长激素依赖性肿瘤细胞增殖[59]。采用丝氨酸、苏氨酸激酶抑制剂AZD1208或Bcl2家族拮抗剂Sabutoclax抑制STAT5,可改善对酪氨酸激酶抑制剂有抗性的费氏染色体阳性急性淋巴细胞白血病的病情[60]。酪氨酸激酶抑制剂奎扎替尼(quizartinib)可通过直接抑制FMS 样酪氨酸激酶3(FMS-like tyrosine kinase,FLT3),抑制JAK-STAT5信号通路的活性,进而导致细胞凋亡,对AML 产生治疗作用[61]。需要注意的是,间接抑制STAT5 会产生一些非特异性效应,如增加机体病毒感染的风险[62]。
3.2 STAT5 与放疗 在与STAT5 过度激活相关的肿瘤中,抑制STAT5活化可增强肿瘤的放疗敏感性。放疗主要通过诱导肿瘤细胞DNA 双链断裂(DNA double strand breaks,DSBs)杀伤肿瘤细胞,但受照肿瘤细胞可能通过损伤修复而存活。若能降低肿瘤细胞DNA修复能力,则可增加肿瘤放射敏感性。放疗是前列腺癌的常用治疗方法,但单独放疗往往效果不佳。Maranto等[63]研究发现,STAT5可以诱导前列腺癌中Rad51 的表达,促进同源重组(homologous recombination,HR)DNA 修复,抑制STAT5 活化可降低HR DNA 的修复能力,增加前列腺癌的放疗敏感性,并且不会诱导邻近组织的辐射敏感性,可作为前列腺癌的放疗辅助疗法。Huguet 等[64]研究发现,酪氨酸激酶抑制剂伊马替尼可通过抑制STAT5 活化,协同辐照抑制CML 细胞K562 增殖。Stegeman 等[65]研究发现,抑制STAT5 活化可增强头颈癌细胞的放疗敏感性。Chen等[66]研究显示,在接受放疗的宫颈癌患者中,STAT5 高表达组肿瘤转移能力较低表达组减弱,生存期更长,提示STAT5有助于提高宫颈癌患者的放疗疗效。
4 总结与展望
STAT5 具有多功能性,STAT5 异常活化与前列腺癌、肺癌、乳腺癌等多种肿瘤的发生发展密切相关。STAT5的促肿瘤作用主要与STAT5过度活化有关,STAT5 上游细胞信号传导异常或STAT5 调控网络中的其他元件发生突变会造成STAT5 的持续激活,导致受STAT5 转录调控的,与细胞存活、增殖相关的基因功能异常,从而促进肿瘤细胞增殖、侵袭和转移。STAT5的肿瘤抑制作用主要体现在免疫系统恶性肿瘤中。STAT5可以通过调节免疫细胞的免疫应答,影响机体的免疫功能,STAT5缺陷的免疫细胞会对肿瘤的发生发展产生促进作用。靶向STAT5可增强前列腺癌、头颈癌等多种肿瘤的放疗敏感性,为肿瘤治疗提供了新的策略。但STAT5A 和STAT5B在不同肿瘤中的作用差异尚未完全明确,另外抑制STAT5 活性的化合物多处于活性筛选的早期阶段,靶向STAT5的治疗方法及有临床应用前景的高选择性、低毒性的新型STAT5抑制剂尚需进一步探索。
