微小RNA及其介导的竞争性内源RNA调控网络在意大利蜜蜂工蜂中肠发育过程中的潜在作用
2020-08-04杜宇范小雪蒋海宾王杰范元婵祝智威周丁丁万洁琦卢家轩熊翠玲郑燕珍陈大福郭睿
杜宇,范小雪,蒋海宾,王杰,范元婵,祝智威,周丁丁,万洁琦,卢家轩,熊翠玲,郑燕珍,陈大福,郭睿
(福建农林大学动物科学学院(蜂学学院),福州 350002)
0 引言
【研究意义】意大利蜜蜂(Apis mellifera ligustica,简称意蜂)是具有重要生态、经济和科研价值的社会性昆虫,广泛应用于我国和其他养蜂国家的养蜂生产[1]。成年蜜蜂的肠道分为前肠、中肠和后肠,其中中肠是食物消化、营养吸收以及免疫防御的主要部位,其内壁富含几丁质的围食膜具有分子筛功能,可保护中肠抵御病原侵染以及促进营养成分吸收[2-3]。微小RNA(microRNA,miRNA)是一类高度保守的长度约为18—25 nt的内源性单链非编码RNA(non-coding RNA,ncRNA)[4],主要通过特异性结合靶mRNA的3′ UTR,导致靶mRNA的翻译抑制或降解,从而在转录后水平行使基因表达调控功能[5]。较多的研究结果表明miRNA广泛参与昆虫生长发育和免疫防御等生物学过程[6-7]。目前,蜜蜂肠道发育的分子机理尚不明确,ncRNA在蜜蜂肠道发育过程中的作用研究还很有限。对意蜂工蜂中肠发育过程的miRNA差异表达谱、调控网络及其差异表达miRNA(differentially expressed miRNA,DEmiRNA)的潜在作用进行全面分析和探讨,可在miRNA组学层面进一步揭示意蜂工蜂中肠发育的分子机理。【前人研究进展】1993年,LEE等[8]首次报道lin-4在秀丽隐杆线虫(Caenorhabditis elegans)体内具有调节生长发育的作用。近十年来,随着高通量测序技术的飞速发展,越来越多的miRNA在动物、植物和微生物中被鉴定出来[9-11]。但相比于果蝇(Drosophila melanogaster)[12]等模式生物,蜜蜂的miRNA研究相对滞后,涉及miRNA参与调控蜜蜂肠道发育的研究尤为缺乏。LIU等[13]研究发现与神经发育相关的miR-31a、ame-miR-210和ame-miR-278等9个miRNA在意蜂哺育蜂与采集蜂头部的差异表达,并推测它们可能影响蜜蜂的劳动分工过程;SHI等[14]研究发现ame-let-7、ame-miR-13b和ame-miR-279在意蜂工蜂脑部的表达具有显著的时空特异性,且长度为22 nt的miRNA在蜂王与工蜂体内的表达量存在显著差异,作者推测这些miRNA参与调控与蜜蜂级型分化相关的信号通路;LOURENÇO等[15]研究发现,意蜂工蜂被革兰氏阳性菌藤黄微球菌(Micrococcus luteus)和革兰氏阴性菌粘质沙雷氏菌(Serratia marcescens)感染后miR-137等38个miRNA差异表达,并与Toll、Imd、JNK和Jak-STAT通路相关的25个mRNA之间存在潜在的调控关系,影响抗菌肽合成和黑化作用激活等免疫防御过程。2011年,SALMENA等[16]首次提出竞争性内源RNA(competing endogenous RNA,ceRNA)假说,认为含有miRNA应答元件(miRNA response element,MRE)的RNA,如长链非编码RNA(long non-coding RNA,lncRNA)、环状RNA(circular RNA,circRNA)和假基因转录本等,可作为ceRNA竞争性结合miRNA,从而间接影响mRNA的表达。此后,该假说已被越来越多的研究结果[17-18]所证实,WANG等[18]研究发现自噬细胞自噬促进因子lncRNA-APF可直接结合miR-188-3p抑制其活性,从而间接调节ATG7的表达以调控自噬程序和自噬细胞死亡。前期研究中,笔者所在课题组运用二代测序技术及生物信息学方法系统解析了意蜂幼虫肠道发育过程的DEmiRNA表达谱及调控网络[19],揭示了miR-342-y、ame-miR-6052、miR-iab-4-x、miR-281-x、novel-m0031-3p等DEmiRNA可能在幼虫肠道发育中发挥重要的调控作用;此外,还相继解析了意蜂工蜂中肠发育过程的差异表达lncRNA(differentially expressed lncRNA,DElncRNA)表达谱、调控网络及潜在作用[20],差异表达circRNA(differentially expressed circRNA,DEcircRNA)表达谱、调控网络及潜在功能[21],DEmRNA表达谱及ceRNA网络[22],在组学层面深入细致探讨了mRNA和ncRNA介导的中肠发育机理,并筛选出若干功能研究的候选分子。【本研究切入点】根据ceRNA机制,miRNA作为联系mRNA与lncRNA和circRNA等ncRNA的桥梁,在调控网络中具有核心地位。笔者所在课题组前期已在mRNA组学、lncRNA组学和circRNA组学层面对意蜂工蜂中肠发育机理进行了探究,本研究在此基础上进一步对中肠发育过程miRNA差异表达谱、DEmiRNA介导的ceRNA调控网络,以及DEmiRNA的潜在功能进行深入分析,从而将mRNA和ncRNA联系起来,展示意蜂工蜂中肠发育过程的ceRNA调控网络全貌。【拟解决的关键问题】利用small RNA-seq(sRNA-seq)技术、生物信息学方法和分子生物学手段对意蜂工蜂中肠发育过程DEmiRNA介导的ceRNA调控网络及潜在作用进行分析和验证,进而与前期在DEmRNA、DElncRNA和DEcircRNA组学层面的研究结果进行比较分析和探讨,以期在组学水平揭示DEmiRNA及复杂调控网络介导的中肠发育机理,为在分子水平阐明意蜂工蜂中肠的发育机理打下基础。
1 材料与方法
试验于2017年9月至2019年10月在福建农林大学动物科学学院(蜂学学院)蜜蜂保护实验室完成。
1.1 供试生物材料
供试意蜂工蜂取自福建农林大学动物科学学院(蜂学学院)教学蜂场。
1.2 中肠样品制备与Illumina测序
按照笔者实验室前期已建立的方法[20-22]进行意蜂的人工饲养。(1)从群势较强且显微镜检无东方蜜蜂微孢子虫的3个蜂群中提取老熟封盖子脾至实验室恒温培养箱(34±0.5)℃;(2)每30 min将刚出房的工蜂(记为0 d)放入四周打孔且已消毒的干净塑料盒(35只/盒),每个塑料盒上方插入一支装有50%(w/v)无菌糖水的饲喂器;每日检查工蜂存活情况,及时清理死亡工蜂;(3)待工蜂出房7 d和10 d时,在超净台用干净镊子拉取工蜂中肠,放入灭菌后的RNA-Free的EP管,经液氮速冻后迅速转移至-80℃超低温冰箱保存备用,每只中肠的取样时间严格控制在15 s以内。进行3次生物学重复,每个生物学重复包含3只中肠样品。7 d工蜂中肠样品(Am7)的3个生物学重复:Am7-1、Am7-2和Am7-3;10 d工蜂中肠样品(Am10)的3个生物学重复:Am10-1、Am10-2和Am10-3。文库构建方法如下:用Trizol法从上述样本中提取total RNA,琼脂糖凝胶电泳切胶选择18—30 nt的片段。然后连接3′接头,连接产物以15%变性PAGE胶电泳分离,切胶选择36—44 nt目的条带。回收切胶产物,连接5′接头,然后对连接了两侧接头的小RNA样本进行反转录PCR。反转录产物以3.5%琼脂糖凝胶电泳分离,切胶选择140—160 bp区域条带,胶回收产物即为终文库。建好的文库委托广州基迪奥生物技术有限公司进行测序,测序平台为Illumina MiSeq。测序数据已上传NCBI SRA数据库,BioProject号:PRJNA408312。
1.3 测序数据质控、参考基因组比对及miRNA的差异表达分析
对于下机的原始读段(raw reads),按照前期已建立的方法[19,23]步骤进行质量控制:(1)过滤掉质量值<20的碱基数超过1个的reads;(2)过滤除掉含有未知碱基(N)的reads;(3)过滤3′或5′接头的reads,并去除长度<18 bp的reads;(4)过滤包含poly A的reads。过滤得到的clean reads用于后续分析。
利用Bowit软件将非注释序列标签(tags)与西方蜜蜂基因组(assembly Amel 4.5)比对,得到相关tags对应的位置信息,即mapped tags。使用miRDeep2软件将mapped tags与miRBase数据库中已知的miRNA前体序列进行比对,鉴定已知miRNA的表达。利用每百万标签序列(tags per million,TPM)算法公式(TPM=T×106/N,T表示miRNA的tags,N表示总miRNA的tags)对表达量进行归一化处理。使用R软件计算意蜂工蜂中肠样品不同生物学重复之间的Pearson相关性系数。显著性DEmiRNA的筛选标准为|log2fold change|≥1且P≤0.05。
1.4 DEmiRNA的靶mRNA预测及分析
通过TargetFinder软件对DEmiRNA进行靶向预测。利用Blast软件将上述靶mRNA序列分别映射GO(Gene Ontology)和KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库。结合本课题组前期研究结果[20-22],得到DEmiRNA与DEmRNA、DElncRNA和DEcircRNA的靶向结合关系,并根据DEmiRNA与DEmRNA的靶向结合关系筛选自由能≤-20 kcal·mol-1的靶mRNA,进行KEGG数据库注释,再通过比对前期在差异基因方面的研究结果[22]找到相同的代谢通路,利用Cytoscape软件对DEmiRNA靶向的DElncRNA、DEcircRNA以及通路中的DEmRNA进行调控网络的构建及可视化。
1.5 DEmiRNA的茎环反转录实时荧光定量PCR(Stemloop RT-qPCR)验证
参考前期已建立的方法[19,23-24],通过Stem-loop RT-qPCR检测4个随机挑选的DEmiRNA(miR-210-z、miR-342-y、miR-7975-y和miR-155-x)在Am7和Am10中的表达情况,以验证数据的可靠性。通过DNAMAN软件设计特异性引物和通用反向引物,委托上海生工生物工程有限公司合成引物,引物信息详见表1。以snRNA U6作为内参,使用M5 microRNA抽提试剂盒(Mei5bio公司,中国)分别提取Am7和Am10的miRNA作为模板,利用Stem-loop引物进行反转录得到相应的cDNA后,以cDNA作为模板进行qPCR。qPCR反应按照SYBR Green Dye试剂盒(Vazyme公司,中国)操作说明书进行。反应体系20 μL包含SYBR Green Dye 10 μL,cDNA模板1.3 μL,正、反向引物各1 μL,DEPC水6.7 μL。qRT-PCR反应在ABI QuantStudio 3荧光定量PCR仪(ABI,美国)上进行,程序设置为95℃预变性5 min;95℃变性10 s,60℃退火延伸30 s,共40个循环。每个反应进行3次生物学重复和技术重复。利用2-ΔΔCt法计算miRNA相对表达量。利用Graph Prism 7软件处理数据和绘图,通过t检验计算组间显著性。
2 结果
2.1 意蜂工蜂中肠测序数据的质控与评估
Am7和Am10的sRNA-seq分别得到13 511 614条和13 448 175条raw reads,严格过滤后分别得到13 119 002条和13 010 751条clean reads,占raw reads的比例均≥96.13%(表2)。此外,Am7和Am10的组内各生物学重复之间的Pearson相关性系数均达到0.9870和0.9988以上(图1)。上述结果说明本研究中样本重复性和数据质量较好,可用于进一步分析。

表1 RT-qPCR引物信息Table 1 Information of primers for RT-qPCR

表2 sRNA-seq数据总览Table 2 Overview of sRNA-seq datasets
2.2 意蜂工蜂中肠发育过程的miRNA差异表达谱
差异表达分析结果显示,Am7 vs Am10比较组共含有112个显著性DEmiRNA,包括38个显著上调miRNA和74个显著下调miRNA。其中,上调倍数最高的是miR-7132-y,其次是miR-2188-x和miR-1388-x(表3);而下调倍数最高的是miR-8503-x,其次是miR-298-x和miR-291-y(表4)。
2.3 意蜂工蜂中肠发育过程中DEmiRNA的靶mRNA预测及分析
利用TargetFinder软件对上述显著上调和显著下调miRNA进行靶向预测,分别预测出7 434和9 559个靶mRNA,并分别涉及47和51个功能条目。其中注释靶mRNA数前15位的条目均为结合(2 082和2 632)、细胞进程(2 074和2 526)、代谢进程(1 767和2 270)、单组织进程(1 588和1 957)、催化活性(1 288和1 720)、细胞(916和1 130)、细胞组件(916和1130)、细胞膜(913和1 098)、细胞膜组件(893和1 076)、细胞器(681和863)、生物学调控(655和775)、生物进程调控(601和729)、定位(565和713)、应激反应(556和679)、信号(494和615)(图2)。括号内数字代表注释在该条目的显著上调(下调)miRNA的靶mRNA数。

表3 意蜂工蜂中肠发育过程中显著上调前10个miRNATable 3 Top 10 significantly up-regulated miRNAs during the developmental process of A.m.ligustica worker’s midgut
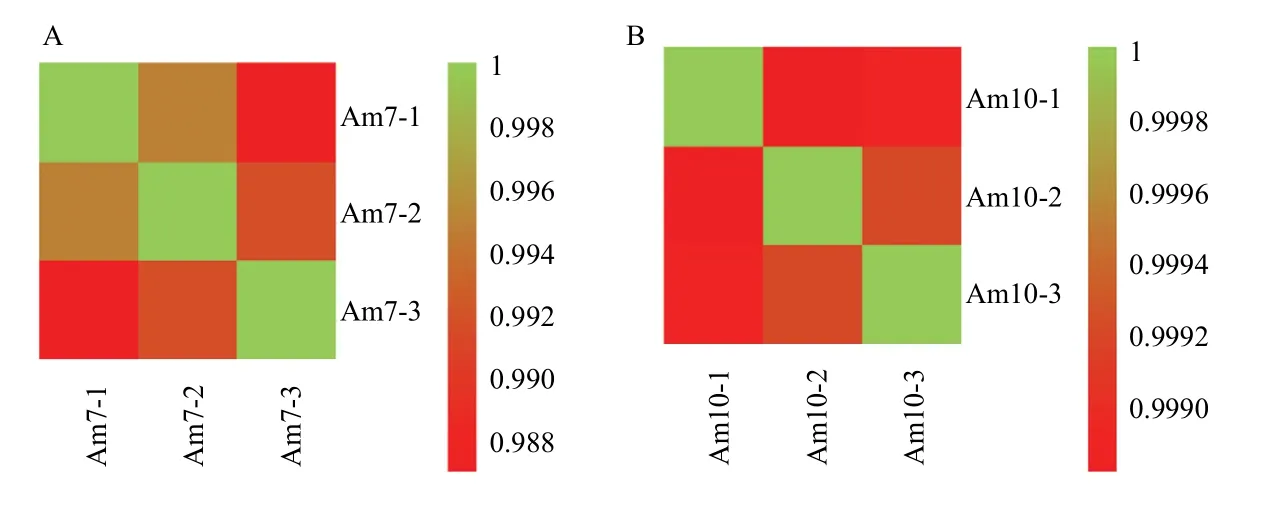
图1 各意蜂工蜂中肠样品不同生物学重复间的Pearson相关性系数Fig.1 Pearson correlation coefficients among different biological replicas within each A.m.ligustica worker’s midgut sample group
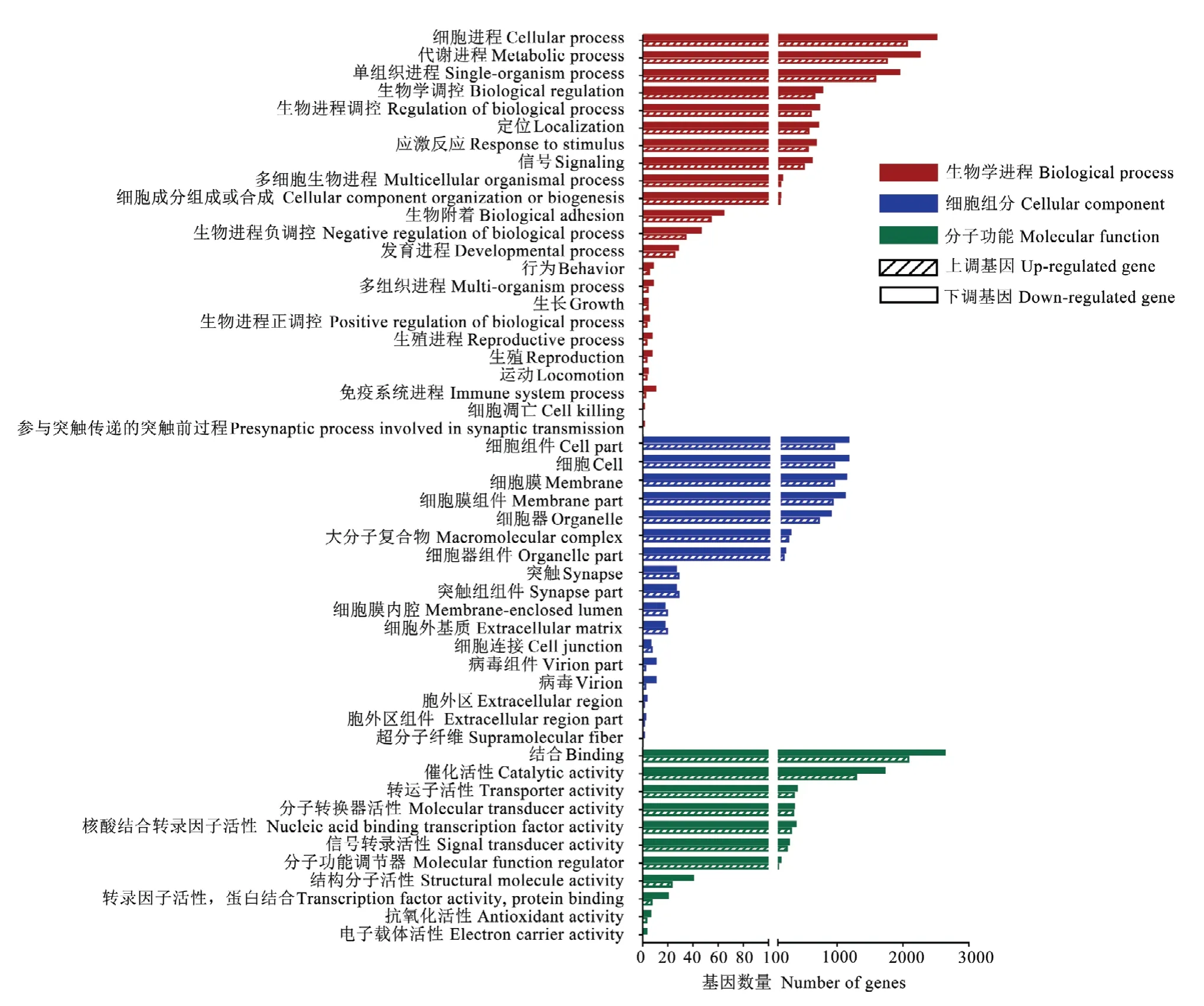
图2 意蜂工蜂中肠发育过程DEmiRNA的GO数据库注释Fig.2 GO database annotation of DEmiRNA target mRNAs involved in the developmental process of A.m.ligustica worker’s midgut

表4 意蜂工蜂中肠发育过程中显著下调前10个miRNATable 4 Top 10 significantly down-regulated miRNAs during the developmental process of A.m.ligustica worker’s midgut
进一步对上述DEmiRNA的靶mRNA进行KEGG数据库注释,结果显示显著上调miRNA的靶mRNA可注释到315条通路,显著下调miRNA的靶mRNA可注释到324条通路。对于显著上调miRNA和显著下调miRNA,分别有118和134个靶mRNA注释到Hippo信号通路,155和190个靶mRNA注释到Wnt信号通路,58和63个靶mRNA注释到FoxO信号通路,30和41个靶mRNA注释到Notch通路;分别有98和109个靶mRNA注释到内吞作用,74和70个靶mRNA注释到泛素介导的蛋白水解,37和32个靶mRNA注释到溶酶体,111和117个靶mRNA注释到黑色素生成等细胞免疫通路;分别有17和22个靶mRNA注释到Jak-STAT信号通路,8和10个靶mRNA注释到NF-κB信号通路,8和18个靶mRNA注释到Toll/Imd信号通路,25和53个靶mRNA注释到MAPK信号通路等体液免疫通路。表5和表6分别展示了显著上调miRNA和显著下调miRNA显著富集的前15位通路。
2.4 意蜂工蜂中肠发育过程中DEmiRNA调控网络的构建及分析
前期研究中,为深入分析意蜂工蜂中肠的发育机理,笔者所在课题组选取13条信号通路(AMPK、PI3K-Akt、Wnt、cAMP、FoxO、Hippo、mTOR、Jak-STAT、Toll-like受体、TGF-beta、Notch、MAPK和NF-κB)及各通路富集DEmRNA构建富集关系网络[22]。在此基础上,根据序列匹配原则,利用软件预测DEmiRNA与前期研究中DElncRNA[20]、DEcircRNA[21]、DEmRNA[22]的靶向结合关系,构建9条通路相关的调控网络,分析结果显示DEmiRNA与DEmRNA、DEcircRNA和DElncRNA之间存在复杂的调控关系,DEmiRNA居于网络的中心位置,而DElncRNA、DEcircRNA和DEmRNA处于网络外周;此外miR-5106-y能够靶向结合3个DElncRNA和2个DEcircRNA(图3)。
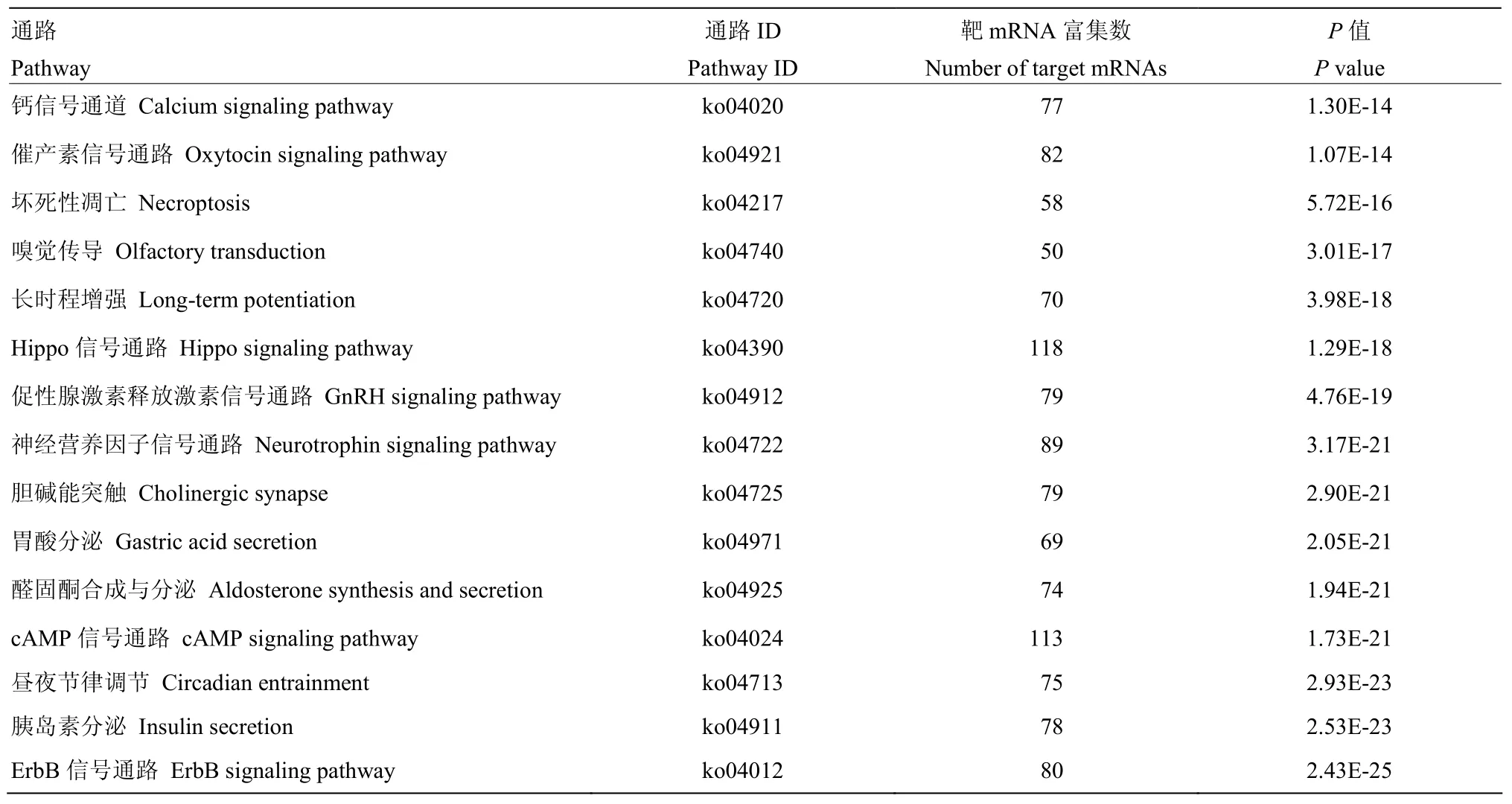
表5 意蜂工蜂中肠上调miRNA的靶mRNA显著富集的前15位通路Table 5 Top 15 pathways significantly enriched by target mRNAs of significantly up-regulated miRNAs in the midgut of A.m.ligustica worker

表6 意大利蜜蜂工蜂中肠下调miRNA的靶mRNA显著富集的前15位通路Table 6 Top 15 pathways significantly enriched by target mRNAs of significantly down-regulated miRNAs in the midgut of A.m.ligustica worker
进一步分析发现,一个DEmiRNA靶向的DEmRNA可注释到多条信号通路,例如miR-291-y靶向的糖原合成酶激酶-3β亚型编码基因(XM_006567354.2)可注释到Wnt、PI3K-Akt和mTOR信号通路,miR-376-y靶向的环盒子亚型编码基因(XM_016910739.1)可注释到Wnt和TGF-beta信号通路,miR-148-y靶向的非特性蛋白LOC411962编码基因(XM_006567604.2)可注释到MAPK和Toll/Imd信号通路,miR-5106-y靶向的丝氨酸/苏氨酸-蛋白磷酸酶-β亚基-ε编码基因(XM_016912389.1)可注释到PI3K-Akt和AMPK信号通路。此外,同一个DEmiRNA靶向的多个DEmRNA富集在同一条信号通路,例如ame-miR-6001-3p靶向的3个DEmRNA(XM_006562526.2、XM_016914568.1和XM_016914811.1)可注释到Hippo信号通路;miR-126-x靶向的2个DEmRNA(XM_006557903.2和XM_006557904.2)均注释到cAMP信号通路(图3)。
miR-342-y在意蜂工蜂中肠发育过程中呈显著性上调表达,与其在意蜂幼虫肠道发育过程的表达趋势[19]相反。进一步对miR-342-y及与其存在靶向结合关系的DElncRNA、DEcircRNA和DEmRNA进行预测和分析,结果显示miR-342-y可靶向结合3个DEcircRNA、4个DElncRNA和327个靶mRNA,上述靶mRNA可注释到松弛素信号通路(15)、趋化因子信号通路(13)、HIF-1信号通路(12)、神经活性配体-受体相互作用(10)、内吞作用(10)、碳水化合物的消化吸收(8)、雌激素信号通路(8)、腹背轴形成(8)和TRP通道的炎性介质调节(8)等162条通路(图4)。
2.5 意蜂工蜂中肠发育过程中DEmiRNA的Stem-loop RT-qPCR验证
利 用Stem-loop RT-qPCR对miR-210-z、miR-342-y、miR-7975-y和miR-155-x进行表达量检测,结果显示它们的差异变化趋势与转录组测序中相应的表达量变化趋势一致(图5),证明了本研究中miRNA差异表达及测序数据的真实可靠性。
3 讨论

图3 意蜂工蜂中肠发育过程中9条信号通路相关的DElncRNA/DEcircRNA-DEmiRNA-DEmRNA关系网络Fig.3 Relationship network of DElncRNA/DEcircRNA-DEmiRNA-DEmRNA associated with 9 signaling pathways in the midgut of A.m.ligustica worker

图4 意蜂工蜂中肠的miR-342-y靶向结合的10条信号通路相关DElncRNA、DEcircRNA、DEmRNA关系网络Fig.4 Relationship network of miR-342-y and its target DElncRNAs, target DEcircRNAs, target DEmRNAs associated with 10 signaling pathways in the midgut of A.m.ligustica worker

图5 DEmiRNA的RT-qPCR验证Fig.5 RT-qPCR validation of DEmiRNAs
昆虫肠道不仅是消化食物和吸收营养的主要场所,也是阻挡病原入侵血淋巴及其他器官的重要防线[25]。目前,有关成年蜜蜂肠道的研究主要集中在肠道微生物的结构特征和功能预测方面[26-27],肠道发育机理的相关研究极其有限。以Illumina为代表的二代测序技术近年来发展迅速,在miRNA等ncRNA的高通量挖掘方面表现出突出的优越性[28-30]。前期研究中,笔者所在课题组对意蜂工蜂响应微孢子虫胁迫的应答机制进行了lncRNA、miRNA和mRNA层面的全面解析[28,30-31];并在lncRNA、circRNA和mRNA组学层面探究了意蜂工蜂中肠的发育机理,系统解析了DElncRNA、DEcircRNA和DEmRNA表达谱及三者涉及的调控网络[20-22]。miRNA作为基因表达调控的关键因子,能够作为桥梁将编码RNA与非编码RNA联系起来。为进一步深入探究意蜂工蜂中肠发育的分子机理,本研究结合测序得到的意蜂工蜂中肠sRNA组学数据和前期获得的中肠circRNA、lncRNA和mRNA组学数据,对miRNA的差异表达谱及调控网络进行了深入分析和探讨。蜜蜂是一种完全变态昆虫,历经卵期、幼虫期、蛹期和成虫期4个阶段,其中,1—2日龄幼虫太小,直接剖取肠道技术上无法满足。前期研究中,笔者所在课题组从自然蜂群中移取2日龄意蜂幼虫至48孔培养板,经24 h的环境适应后,于3日龄时处理组幼虫饲喂接种球囊菌孢子,对照组幼虫饲喂不含球囊菌孢子的饲料,然后于4、5和6日龄分别剖取处理组和对照组幼虫肠道用于sRNA-seq;基于高质量的sRNA组学数据全基因组鉴定和分析了意蜂幼虫肠道miRNA及其结构特征[9],并解析了意蜂幼虫肠道发育过程的miRNA差异表达谱、调控网络及潜在功能[19]。蜜蜂预蛹期,内部的组织经历分解和重构,逐渐形成新的组织,此阶段无法获取完整肠道。本研究仅选取意蜂7日龄和10日龄工蜂中肠进行sRNA-seq,因为这两个时间点均处于意蜂成虫期,且本研究的分析和讨论未涉及变态发育。
从Am7 vs Am10比较组中筛选出38个显著上调miRNA和74个显著下调miRNA,可分别靶向7 434和9 559个mRNA,通过与circRNA、lncRNA和mRNA的相互作用参与调节意蜂中肠发育过程中的各类生命活动。
3.1 DEmiRNA参与调控意蜂中肠发育过程的新陈代谢、信号传导和免疫应答
蜜蜂肠道在消化和吸收等正常生命活动中伴随着活跃的物质和能量代谢。本研究发现,显著上调和下调miRNA靶向结合的mRNA均可注释到嘌呤代谢(81和104个靶mRNA)、甘油磷脂代谢(49和52个靶mRNA)、醚脂质代谢(13和22个靶mRNA)等脂代谢相关通路;果糖和甘露糖代谢(23和23个靶mRNA)、磷酸肌醇代谢(43和64个靶mRNA)等碳水化合物代谢相关通路;赖氨酸降解(13和21个靶mRNA)、甘氨酸、丝氨酸和苏氨酸代谢(11和17个靶mRNA)等氨基酸代谢相关通路。此外,显著上调和下调miRNA靶向结合的mRNA还能注释到氮代谢(2和6个靶mRNA)和氧化磷酸化(22和25个靶mRNA)等多条能量代谢相关通路。上述结果表明意蜂工蜂中肠发育过程中可通过DEmiRNA调节部分物质代谢和能量代谢通路及相关基因的表达,以满足中肠生长和发育的物质和能量需求。
昆虫肠道在生长发育过程中受到Hippo、Wnt、FoxO和Notch等信号通路的共同调节[32]。中肠干细胞能够分化为具有吸收功能的肠上皮细胞以及分泌功能的肠内分泌细胞,保证肠组织的自我更新和损伤修复,从而维持正常的形状和功能,而Notch、Hippo和Wnt信号通路在中肠干细胞的稳态平衡、更新分化过程中发挥重要作用[33-34]。Notch信号通路在果蝇胚胎期中肠发育过程中调节成体中肠祖细胞[35],在成年期调控中肠干细胞以维持中肠内环境稳态[36-37]。Wnt信号通路通过控制果蝇成年个体肠道的腔室内干细胞增殖以及调控腔室边界细胞正常发展,从而在内环境平衡和发育过程中发挥重要作用[38]。上皮细胞Hippo信号的缺失会增加Upd家族细胞因子的产生,并且刺激EGFR配体,进而激活Jak-STAT和EGFR信号通路并促进果蝇中肠干细胞的增殖[39]。昆虫蜕皮是生长发育的重要生理过程,而FoxO信号通路中的FoxO通过调控CPA的表达参与调控昆虫的蜕皮行为[40]。本研究中,对于显著上调和显著下调miRNA,分别有118和134个靶mRNA注释到Hippo信号通路,155和190个靶mRNA注释到Wnt信号通路,58和63个靶mRNA注释到FoxO信号通路,30和41个靶mRNA注释到Notch通路。相应的DEmiRNA通过上调或下调部分基因的表达水平对上述发育相关信号通路进行调控,从而影响意蜂工蜂中肠的生长和发育以及细胞分化。
昆虫中肠作为与外源病原体互作的重要场所,可通过分泌活性氧、抗菌肽以及各类蛋白酶协助宿主抵御病原侵染[25]。蜜蜂在与病原长期协同进化过程中,形成了以内吞作用、吞噬作用、黑化作用和蛋白酶促水解为代表的细胞免疫,以及Jak-STAT、NF-κB、Imd/Toll、MAPK和JNK等信号通路和抗菌肽释放为代表的体液免疫[41]。前人研究发现Jak-STAT信号通路作为昆虫先天免疫系统的重要组成部分,在病原侵染的过程中可被显著激活并广泛参与细胞凋亡、免疫调节等过程[42]。Toll/Imd信号通路和NF-κB信号通路共同介导了抗菌肽的合成与释放过程,并协同吞噬和包埋作用参与昆虫的应激免疫反应[43-44]。JNK途径在内的MAPK级联反应的激活,能够促进昆虫免疫的调节过程[45-46]。本研究发现,对于显著上调和下调的miRNA,分别有98和109个靶mRNA注释到内吞作用,74和70个靶mRNA注释到泛素蛋白水解,37和32个靶mRNA注释到溶酶体,111和117个靶mRNA注释到黑色素生成等细胞免疫通路;此外,分别有17和22个靶mRNA注释到Jak-STAT信号通路,8和10个靶mRNA注释到NF-κB信号通路,8和18个靶mRNA注释到Toll/Imd信号通路,25和53个靶mRNA注释到MAPK信号通路等体液免疫通路。上述结果共同表明,意蜂中肠发育过程中的DEmiRNA具有调节免疫应激反应的潜在作用。
3.2 DEmiRNA参与意蜂工蜂中肠发育过程的ceRNA调控网络
LncRNA和circRNA可作为ceRNA,通过MRE竞争性结合miRNA,以减轻miRNA对靶mRNA的抑制或降解作用,间接调控各类生物学过程[17-18]。笔者所在课题组前期研究发现,意蜂工蜂中肠的发育过程中,DEcircRNA和DElncRNA可分别结合多个miRNA,如lncRNA XR_001702484.1可靶向ame-miR-6001-3p、miR-376-y和miR-182-x等10个DEmiRNA[20];circRNA novel_circ_010719可靶向ame-miR-6001-3p、miR-148-y和miR-291-y等43个DEmiRNA[21]。经比较分析发现,本研究有112个DEmiRNA能够被201个DEcircRNA、112个DElncRNA和283个DEmRNA竞争性结合,其中靶向16个DEmRNA的miR-126-x(log2fold change=1.96,P=4.54E-04)可被11个DEcircRNA和9个DElncRNA竞争性结合。上述结果表明,miRNA作为ceRNA调控网络的核心因子,通过介导circRNA和lncRNA对基因表达的调控,在意蜂工蜂中肠发育过程中发挥关键调控作用。
一个mRNA可被多个miRNA靶向结合,并参与对多条信号通路的调控作用。前期研究发现,意蜂工蜂中肠发育过程中的57个DEmRNA涉及cAMP、AMPK、Hippo、Toll-like受体、MAPK、mTOR、Wnt、PI3K-Akt、FoxO、Jak-STAT、TGF-beta、Notch和NF-κB 13条发育和免疫相关信号通路[22]。结合前期研究结果进行比较分析,本研究发现miR-291-y、miR-376-y、miR-148-y、miR-5106-y、ame-miR-6001-3p、miR-182-x、miR-126-x、miR-3223-x 8个DEmiRNA靶向结合的12个DEmRNA主要富集在cAMP、AMPK、Hippo、Toll/Imd、TGF-beta、MAPK、mTOR、Wnt和PI3K-Akt 9条信号通路(图3)。XM_016912389.1编码的丝氨酸/苏氨酸-蛋白磷酸酶-β亚基-ε在细胞增殖、转化、凋亡及代谢等多种细胞事件中发挥重要调控作用[47]。本研究中,miR-5106-y的靶mRNA XM_016912389.1富集在PI3K-Akt和AMPK信号通路,说明miR-5106-y在意蜂工蜂中肠发育过程涉及这两条信号通路的调控。此外,分析结果显示同一DEmiRNA的靶向结合的多个mRNA富集在同一条信号通路,例如ame-miR-6001-3p靶向结合的4个DEmRNA中有3个富集在Hippo信号通路。前期研究发现多达29个DElncRNA[20]和14个DEcircRNA[21]能够靶向结合ame-miR-6001-3p,推测ame-miR-6001-3p在意蜂工蜂中肠发育过程除了能够直接调控细胞的增殖与凋亡、组织生长、器官大小,以及组织稳态维持,还能作为DElncRNA和DEcircRNA参与调控发育的媒介分子,值得进一步深入研究。cAMP信号通路在细胞中发挥信使的作用,细胞内cAMP浓度的改变影响多种细胞内信号转导途径,从而调控基因表达、蛋白活性以及细胞功能,对细胞的代谢、生长、分化和凋亡发生影响[48]。本研究中,miR-126-x靶向结合的2个DEmRNA都富集在cAMP信号通路,推测可能通过细胞内外信息传递调控细胞活动。ZHOU等[49]探究了lncRNAmiRNA-mRNA网络在舌鳞状细胞癌(SCCT)中的作用,与本研究中miR-148-y同家族的hsa-miR-148b-3p和hsa-miR-148a-3p可以靶向结合lncRNA KCNQ1OT1参与诱导SCCT细胞生长,推测本研究中的miR-148-y能够通过ceRNA网络在意蜂发育过程中参与调节中肠细胞的生长和发育。酪蛋白激酶在围食膜形成的过程中发挥作用[50],本研究中,ame-miR-6001-3p等28个DEmiRNA能够靶向XM_006569003.2在内的25个编码酪蛋白激酶相关蛋白的基因,暗示以上DEmiRNA参与调控围食膜的形成。几丁质是昆虫表皮和围食膜的重要组成成分之一,几丁质代谢随着昆虫不同生长发育阶段而变化[51],本研究发现ame-miR-6001-3p、miR-142-x、miR-3964-y等10个miRNA能够靶向结合XM_396925.6等4个能够编码几丁质酶的相关基因,推测此10个DEmiRNA参与肠道几丁质的形成过程。
昆虫肠道的发育过程伴随着复杂而广泛的mRNA与ncRNA的相互作用。前期研究发现miR-8503-x、miR-342-y、miR-4792-x和miR-5106-y等miRNA在意蜂幼虫肠道发育过程中发生显著性差异表达[19];本研究中,上述4个miRNA在意蜂工蜂中肠发育过程也呈现出显著性差异表达,差异变化倍数分别达到-14.08(P=3.09E-04)、-3.18(P=4.11E-06)、-3.31(P=8.04E-03)和-1.82(P=0.038),以上4个miRNA在意蜂的两种虫态均发生不同幅度的差异表达,暗示它们与意蜂不同虫态的肠道发育存在潜在关联。此外,还发现miR-342-y在意蜂幼虫肠道发育过程显著性下调表达[19];本研究中,miR-342-y在意蜂工蜂中肠发育中显著性上调表达,与其在幼虫肠道发育过程表达趋势相反,说明该miRNA在意蜂不同虫态的肠道发育过程扮演着不同角色。进一步分析发现,miR-342-y在ceRNA调控网络中可靶向结合3个circRNA、4个lncRNA和327个mRNA,进一步对上述靶mRNA进行代谢通路注释,结果显示参与对神经活性配体-受体相互作用、吞噬作用以及背腹轴形成等162条通路的调控。推测以miR-342-y为核心的ceRNA网络在意蜂不同虫态的肠道发育过程中发挥重要的调控作用,但在意蜂幼虫肠道和工蜂中肠相反的表达趋势暗示miR-342-y具有多面功能,值得进一步深入研究。
针对本研究筛选出的关键DEmiRNA,下一步将通过人工合成miRNA的类似物和抑制物,对意蜂工蜂和幼虫进行miRNA的过表达或敲减,同时检测对其靶向结合lncRNA、circRNA和mRNA表达水平的影响,进而揭示ceRNA介导意蜂工蜂中肠发育的分子机理。
4 结论
DEmiRNA可能通过参与调控的物质和能量代谢通路、Hippo和Wnt等信号通路以及细胞和体液免疫通路相关基因的表达影响意蜂中肠的生长和发育;miR-182-x、miR-291-y、miR-342-y、ame-miR-6001-3p等关键DEmiRNA介导的ceRNA调控网络可能在意蜂中肠发育过程发挥重要的调控作用。
