广东汕尾凹顶藻类(红藻门)的形态分类学研究*
2020-08-02姜晶晶刘金梅刘美媛黄冰心丁兰平
姜晶晶,刘金梅,马 鑫,刘美媛,黄冰心,丁兰平
(天津师范大学生命科学学院,天津市动植物抗性重点实验室,天津 300387)
0 引言
凹顶藻类 (LaurenciaComplex)隶属于红藻门Rhodophyta、仙菜目Ceramiales、松节藻科Rhodomelaceae,是以Lamouroux建立的凹顶藻属Laurencia(Lamouroux,1831)为基础的分类群。凹顶藻作为一个大类群,已报道391个物种,其中被确认的有209种[1],主要分布在全球的温暖水域中[2-4]。因其富含次生代谢产物,在藻胶、药品、食品和化妆品等领域具有重要的应用前景[5-13]。
凹顶藻类的形态变异大、可塑性强[14-15],受生长环境的影响,许多物种间的形态鉴定特征交叉,缺乏明确的分类学界限,物种的界定难度大[14,16-17]。因此,自凹顶藻属建立以来,该类群的分类系统多次变动,从早期一属四组[18]、二属二亚属八组二类群[19]、三属八组二类群[20]、四属[21-22]、五属[23]、六属[24]、七属[25],到现在八属[26]的分类系统,该类群的准确分类存在一定难度。
我国目前凹顶藻类的分类系统仍基于四属系统,已经报道3属37种,即凹顶藻属Laurencia、软凹藻属Chondrophycus和栅凹藻属Palisada,其中凹顶藻属21种、软凹藻属9种、栅凹藻属7种[27],与国际研究水平还存在较大的差距。另外,就具体的物种鉴定指标而言,我国凹顶藻类的分类研究主要集中在形态学方面[28],因而,对凹顶藻类物种的区分相对困难,如软骨软凹藻C.cartilagineus和波状软凹藻C.undulatus经常引起混淆[14]。因此,对凹顶藻类物种进行详细的分类鉴定仍尤为重要。
广东汕尾市位于南海北部,海域辽阔,海岸线长,近海岛屿多,属南亚热带季风气候区,自然环境适合海藻的生长。为了解该地区凹顶藻的物种多样性情况,对采自该地区的标本进行研究,这对了解中国南海北部的凹顶藻物种多样性、资源保护,以及加快该地的凹顶藻属物种资源开发和利用具有指导意义。
1 材料与方法
1.1 材料
实验所用材料分别于2013年5月和2014年5月采集于广东省汕尾市,经纬度为22°38′N,114°54′E。
标本分为2类:(1)腊叶标本,用于直观观察并保存;(2)液浸标本,用于形态及解剖学特征观测。
1.2 方法
1.2.1 外形特征比较
观察藻体的颜色、质地等。在体视显微镜 (Nikon D5-U3)下进一步观察藻体的基部、各部分的分枝情况、顶端形状、四分孢子囊排列方式以及囊果的形状,进行拍照及数据测量。
1.2.2 内部特征比较
液浸样品清洗后,挑选完整的观测部位,利用冷冻切片技术 (LEICA CM 1860 UV),于生物冷冻切片机内对液浸样品进行切片,切片厚度20—50 μm,切片转置载玻片,利用亚甲基蓝溶液或0.5%的苯胺蓝溶液进行染色,将装片置于正置荧光显微镜 (LEICA DM5000 B)下观察,拍照,并进行特征测量。
1.2.3 解剖特征比较
主要对藻体内部结构和生殖器官等特征进行观察、比较和分析,包括藻体表皮和皮层细胞的形状和大小,栅状与否;四分孢子囊的形状、大小和排列方式;是否存在次生纹孔连结;围轴细胞数目;是否存在樱桃体等。
1.2.4 物种鉴定
根据所得实验结果,综合查阅文献资料进行系统对比分析,并进行物种鉴定。
2 结果与分析
2.1 波状软凹藻 (图1)
Chondrophycusundulatus(Yamada) Garbary et Harper,1998:185-200;Nam,1999:455-468;Ding,2003:237-238;Tseng et al.,2011:69-70.
LaurenciaundulataYamada,1931:243-244;Tseng,1943:206;Saito,1967:59;Tseng et al.,1980:57-84;McDermid,1988:231-247;Zhang et Xia,1988:249-252.
特征描述:藻体黑紫或紫褐色,直立,丛生,软骨质地,基部盘状,高约8 cm (图1a、b);具匍匐的基部分枝,但不丰富;主分枝直立,近基部圆柱状,宽约1.1—1.8 mm,向上逐渐扁平 (图1c),宽3.5—6.0 mm;羽状分枝,有时亚二歧分枝;次生分枝一般产生许多短的多节末小枝。
横切面上,表皮细胞呈栅状排列,内部细胞存在纹孔连接 (图1h、i)。分枝纵切面上,表皮细胞呈栅状排列 (图1g、k、l),长40—60 μm,宽14—23 μm;皮层细胞间不存在纵向的次生纹孔连结;髓细胞壁缺乏透镜增厚;末小枝的端细胞位于端部凹陷的底部;藻体干燥后不粘在纸上。
四分孢子囊末小枝不扁压,截形或瘤状,直径400—800 μm;四分孢子囊与中央轴呈垂直排列,散生在截形或瘤状的末小枝的端部表面 (图1d、e、f),四分孢子囊呈四面体型分裂 (图1m)。

(a—b)藻体;(c)小枝顶部;(d)四分孢子囊(箭头);(e)四分孢子囊株;(f)四分孢子囊(箭头);(g)藻体横切;(h)藻体横切之边缘细胞;(i)横切面之次生纹孔连接(箭头);(j—k)藻体顶部纵切;(l)藻体顶部纵切之边缘细胞;(m)四分孢子(箭头示四面体型分裂)(a—b)Thalli;(c)Ultimate branchlet;(d)Tetrasporangia (arrows);(e)Tetrasporangial branches;(f)Tetrasporangia (arrows);(g) Transverse section of thallus;(h) Palisade epidermal cells in transverse section of thallus;(i) Secondary pit-connections (arrows);(j—k) Longitudinal section of ultimate branchlet;(l) Longitudinal section of ultimate branchlet,superficial cortical cells;(m) Tetraspores (arrows indicate tetrahedral splitting)图1 波状软凹藻Fig.1 Chondrophycus undulatus
本文样品未观察到囊果。但据文献报道囊果梨形,多个集生,侧生于小枝上[27-28]。
习性:生长在潮间带下部的岩石上或石缝中。
模式标本产地:日本神奈川省江岛。
国内分布:广泛分布于东南沿海,如浙江省,福建省,广东省,海南省海南岛。
国外分布:菲律宾,朝鲜半岛,日本,夏威夷群岛。
研究标本编号:20130427007,采于广东省汕尾市,标本存放于天津师范大学生命科学学院藻类研究室。
2.2 凹顶藻 (图2)
LaurenciachinensisTseng,1943:198; Zhang et Xia,1988:249-252; Ding,2003:62-71;Tseng et al.,2011:88-90.
特征描述:藻体紫红色,软骨质地,高约8 cm (图2a);向下产生假根;具匍匐枝,圆柱状,宽约0.2—0.3 mm;具独立主枝,通常在幼期明显,主枝羽状分枝丰富,近基部圆柱状,向上逐渐扁压,宽约1.2 mm;藻体呈金字塔型或伞状花序状 (图2b、c);幼期分枝明显二歧或对生,后续的较幼分枝多少呈三歧或多歧;末小枝简单,亚圆柱状,二歧或三歧,长于顶部凹陷处 (图2p),端部截形。

(a)藻体;(b)藻体分枝及顶部;(c)四分孢子囊株;(d)四分孢子囊小枝;(e)藻体小分枝顶部;(f—g)囊果;(h)藻体横切面;(i)藻体横切之边缘细胞;(j)藻体横切之次生纹孔连接(箭头示次生纹孔连接);(k)藻体横切之围轴细胞及4个围轴细胞(箭头示4个围轴细胞);(l)四分孢子囊(箭头示四面体型分裂);(m—n)囊果横切;(o)囊果横切之边缘细胞;(p)藻体顶部新生小枝(a) Thalli;(b) Branches and ultimate branchlet of thallus;(c) Branch of a tetrasporangial plant (tetrasporophyte);(d) Ultimate branchlet with tetrasporangia;(e) Ultimate branchlet;(f—g) Cystocarps;(h) Transverse section of thallus;(i) Superficial cortical cells of transverse section;(j) Secondary pit-connections (arrows indicate secondary pit-connections);(k) Transverse section of thallus showing axial cell with 4 periaxial cells (arrows indicate 4 periaxial cells);(l) Tetrasporangia (arrows indicate tetrahedral splitting);(m—n) Transverse section of cystocarp;(o) Superficial cortical cells of cystocarp on transverse section;(p) New branchlets born on ultimate branchlet图2 凹顶藻Fig.2 Laurencia chinensis
小枝横切面的表皮细胞亚方形 (图2h、i),长约25—45 μm,宽20—40 μm,似栅状排列,存在次生纹孔连结 (图2j);横切面的每轴节产生4个围轴细胞 (图2k);纵切面的表皮细胞非栅状排列;髓细胞壁存在透镜增厚;末小枝端部的表皮细胞稍突起;表皮细胞间存在纵向的次生纹孔连结。
四分孢子囊散生于末小枝的端部附近,球形 (图2l),直径约120 μm,与轴平行排列(图2c、d);囊果在末小枝的上部对生或端部聚生,壶形,具有短喙或无喙 (图2f、g、m、p)。
习性:生长在潮间带下部的泥沙覆盖的岩石上。
模式标本产地:中国香港。
国内分布:中国东海以南沿海,如福建省晋江、平潭和东山,广东省汕尾,香港白沙滩和大浪湾。
国外分布:韩国,澳大利亚,新西兰。
研究标本编号:20130528007,20140518009,采于广东省汕尾市,标本存放于天津师范大学生命科学学院藻类研究室。
2.3 三列凹顶藻 (图3)
LaurenciatristichaTseng,Chang,Xia et Xia,1982:81; Zhang et Xia,1988:249-252; Ding,2003:149-154; Tseng et al.,2011:114-116.
特征描述:藻体淡绿色或暗紫色,一般丛生,柔软质地,高5—10 cm,具有直立主枝和匍匐假根 (图3b),直立主枝及顶,圆柱状,直径1.4—1.6 mm,不规则1—2次互生分枝;分枝明显呈三列 (图3c);老的藻体上产生大量的附生末小枝,末小枝长棒状 (图3d),顶部凹陷。藻体干燥后完全粘在纸上 (图3a)。
藻体表皮细胞不突出;横切面上表皮细胞正方形 (图3h),长19—22 μm,宽16—22 μm,不呈栅状排列,辐射状伸长 (图3g、h);纵切面上表皮细胞非栅状排列 (图3j、k);髓细胞壁具透镜增厚;老株的表皮细胞成排排列,具有纵向的纹孔连结;分枝每轴节产生4个围轴细胞 (图3i);每个表皮细胞及生毛体细胞各含一个樱桃体。
四分孢子囊小枝棒状 (图3f),简单或偶尔分枝一次,长1.0—2.5 mm,直径0.5—1.0 mm;四分孢子囊卵形或长卵形,长61—106 μm,直径51—76 μm,四面体型分裂,与小枝轴呈平行排列 (图3f);具疣状小枝的雄植株简单或端部分枝;囊果着生于末小枝的上部,壶形,具有短喙或无喙 (图3e)。
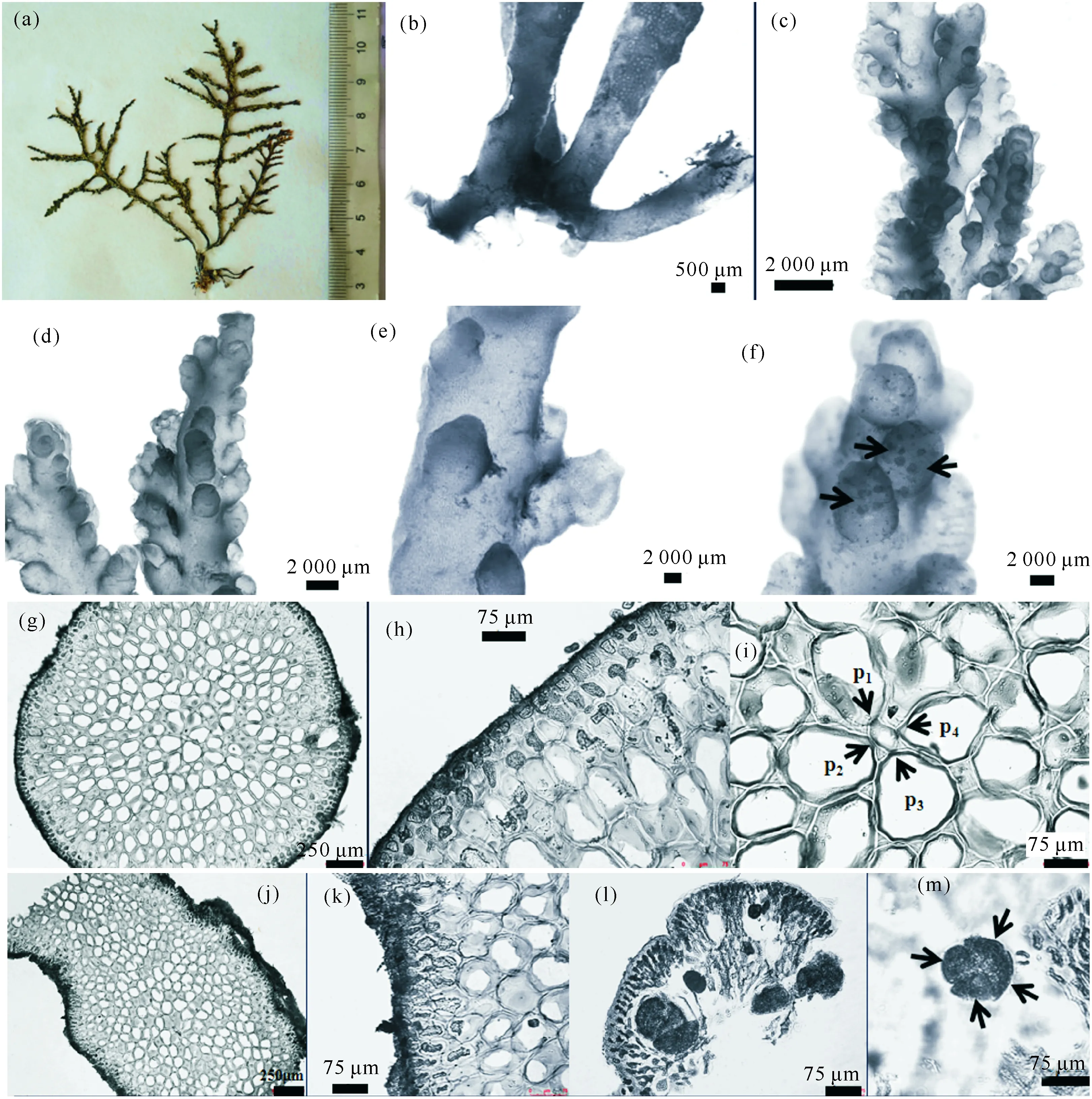
(a)藻体;(b)藻体基部;(c)藻体分枝;(d)末小枝;(e)囊果;(f)四分孢子囊(箭头);(g)藻体横切;(h)藻体横切之表皮细胞;(i)藻体横切之围轴细胞及4个围轴细胞(箭头示 4 个围轴细胞:P1为第1个围轴细胞,P2为第2个围轴细胞,P3为第3个围轴细胞,P4为第4个围轴细胞);(j)藻体纵切面;(k)藻体纵切之表皮细胞;(l)四分孢子囊之纵切面;(m)四分孢子(箭头示四面体型分裂)(a) Thallus;(b) Base part of thallus;(c) Branches;(d) Ultimate branchlet;(e) Cystocarps;(f) Tetrasporangia (arrows);(g) Transverse section of thallus;(h) Superficial cells on transverse section of thallus;(i) Transverse section of thallus showing axial cell with 4 periaxial cells (arrows indicate 4 periaxial cells: P1 is the first axis cell, P2 is the second axis cell, P3 is the thirdt axis cell, P4 is the fourth axis cell);(j) Longitudinal section of thallus;(k) Epidermal cells on Longitudinal section of thallus;(l) Longitudinal section of tetrasporangial branches;(m) Tetraspores (arrows indicate tetrahedral splitting)图3 三列凹顶藻Fig.3 Laurencia tristicha
习性:潮间带岩石上或覆沙的石沼中。
模式标本产地:中国香港。
国内分布:广东省汕尾市,中国香港。
中国特有种类。
研究标本编号:20140518011,采于广东省汕尾市,标本存放于天津师范大学生命科学学院藻类研究室。
2.4 小瘤栅凹藻 (图4)
Palisadaparvipapillata(Tseng) Nam,2007:53-55;Tseng et al.,2011:82-84.
LaurenciaparvipapillataTseng,1943:185-214;Saito,1969:148-160;Cribb,1983:1-173;Nam,1990:41:107-120.
Chondrophycusparvipapillatus(Tseng) Garbary et Harper,1998:195; Nam,1999:455-468;Lipkin et al.,2002:1-90;Ding,2003:220-230;Huisman et al.,2007:264.
特征描述:藻体黑红色至紫红色,高约2.0—4.5 cm,软骨质,多匍匐于珊瑚枝或碎珊瑚上,具盘状固着器 (图4a),固着器上常产生几个独立主枝,主枝匍匐或俯卧,下部约为圆柱状,宽约1 mm,中上部逐渐变成扁压状 (图4b、4c),宽约1.6—2.0 mm;复羽状分歧,具明显的互生或亚对生分枝,分枝可达4级;藻体顶部凹陷;最末小枝棍棒状,顶端截形或圆形,可产生繁殖结构 (图4d、4e、4f);干燥后不易附着在纸上。
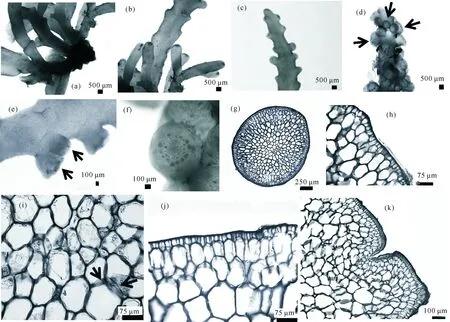
(a)藻体基部;(b)分枝;(c)分枝顶部;(d—f)四分孢子囊枝(箭头示四分孢子囊);(g)藻体横切;(h)藻体横切面的栅状表皮细胞;(i)藻体横切面细胞间的透镜增厚(箭头示透镜增厚);(j)藻体纵切之表皮栅状细胞;(k)藻体顶部纵切面(a) Base part of thallus;(b) Branches;(c) Ultimate branchlet;(d—f) Tetraspoangial branchlets (arrows indicate Tetraspoangial);(g) Transverse section of thallus;(h) Palisade epidermal cells on transverse section of thallus;(i) Transverse section of thallus showing lenticular thickenings (arrows indicate lenticular thickenings);(j) Longitudinal section of palisade epidermal cells;(k) Longitudinal section of ultimate branchlet图4 小瘤栅凹藻Fig.4 Palisada parvipapapillata
表皮细胞表面观为椭圆形或多角形;在横切面上,表皮细胞放射延长,栅状排列 (图4g、h),(20—60) μm×(9—18) μm;在纵切面上,表皮细胞放射延长,栅状排列 (图4j、k),(20—52) μm×(9—17) μm;相邻表皮细胞间存在纵向的次生纹孔连结,有时存在零星的侧向次生纹孔连结;每个轴细胞产生2个围轴细胞;髓细胞壁没有透镜增厚;表皮细胞没有樱桃体存在。
四分孢子囊 (图4d)长100—140 μm、宽40—90 μm,四面体型分裂;环状排列,向心成熟 (图4f),在可育分枝上与轴垂直排列 (图4e)。囊果、精子囊未见。
习性:植株生长在潮间带下部的石灰石上。
模式标本产地:中国香港。
国内分布:广东省,香港,广西壮族自治区,海南省的海南岛和西沙群岛等地。
国外分布:日本,越南,马来西亚。
研究标本编号:20130528018,采于广东省汕尾市,标本存放于天津师范大学生命科学学院藻类研究室。
3 讨论
3.1 物种鉴定中存在的问题
随着分子技术的不断发展,越来越多的藻类学家热衷于采用系统发育分析法来探索物种间的进化关系,这导致凹顶藻类的分类系统变得越来越复杂。凹顶藻类经历了从早期的一属四组 (线型组Filiformes、瘤状组Papillosae、钝型组Obtusae、羽状组Pinnatifidae)[18]和 (软骨组Cartilagineae、福氏组Forterianae、栅状组Palisadae、羽状组Pinnatifidae)[29]、一属二亚属 (凹顶藻亚属与软骨凹顶藻亚属)五组及一个特殊类群 (凹顶藻组Laurencia sensu stricto、福氏组Forsterianae Yamada、羽状组Pinna-tifidae J.Agardh、软骨组Chondrophycus、栅状组Pa-lisadae Yamada、“Spectabilis” group)[30]、一属二亚属六组及一个特殊类群 (第六个组为扁压组Planae Saito et Womersley)[31]、一属二亚属七组及一个特殊类群 (第七个组为节枝组Articulate Zhang et Xia)[32]、一属三亚属八组 (第三亚属为Saitoa;第八组为Pelagosae)[33]、二属(Laurencia和Osmundea)二亚属 (Laurencia和Chondrophycus属于凹顶藻属)[19]、三属 (LaurenciaLamouroux,ChondrophycusTokida et Satio,OsmundeaStackhouse)、四属 (PalisadaNam)[21-22]、五属 (Yuzurua(Nam) Martin-Lescanne)[23]、六属 (LaurenciellaCassano)[34]、八进化枝 (Laurencia,Chondrophycus,Palisada,Laurenciella,Yuzurua,Coronaphycus,Osmundea1,Osmundea2)[4]、七属 (CoronaphycusMetti)[4]到八属 (OhelopapaRousseau,Martin-Lescanne,Payri et Gall)[26]等的分类系统。
早期,凹顶藻类海藻主要分布于中潮带至低潮带的岩礁上,采集困难,且不同的生境下物种的形态也会有差异,这导致了对这类物种进行形态鉴定时出现不同的结果。随着现在分子技术的广泛使用,凹顶藻类海藻的分类系统变得更加复杂,主要有如下原因:(1)进化枝中可能存在大量的同物异名种,如Palisadapapillosa与P.perforata被认为是同物异名[35]、巴西和加那利群岛的L.filiformis、L.majuscula和L.obtusa均为L.dendroidea的同物异名[34]等;(2)基因库中物种信息不全,无法全面比对,导致很多已有种被当成新物种重新命名。
很明显,基于对所选分子标记的分析,可充分明确不同凹顶藻类分类群的亲缘关系。随着分析样本的不断扩大,将能清晰地阐明不同样本或家族的演化过程和亲缘关系,为物种的精确鉴定提供科学依据。然而,就目前的研究现状来说,分子生物学方法显著地扩大了经典形态分类中对物种各级分类系统的划分界线,其本质是扩充了传统生物学“物种”定义。传统生物学中对物种的定义是依据表型特征识别区分生物,而现代遗传学则把物种定义为“物种是一个具有共同基因库、与其他类群有生殖隔离的群体”。以功能性的分子标记为依据的亲缘关系划分,如基于rbcL、COI、18S等功能性序列来进行进化分析,使分类标准过于精细,模糊了当前生物学中坚守的真实“物种”界定标准,可能人为地抬升了遗传学中具有相同或相似遗传功能性特征的“品种”“品系”等种下单位为种级界定标准,这种为物种或更高级分类系统划分制造的新标准,可能导致目前凹顶类种类的增加及分类系统的混乱。
3.2 汕尾凹顶藻类的分类探讨
软凹藻属Chondrophycus由Garbary et Harper于1998年建立,已报道41种及1个亚种,现已有16种得到确认[1]。该属藻体的每个轴有两个围轴细胞,且四分孢子囊与轴垂直排列[28,36]。
波状软凹藻Chondrophycusundulatus隶属于软凹藻属,最初由Yamada于1931年报道,命名为LaurenciaundulataYamada[29,36-40]。然而,Garbary et Harpery (1998)基于支序分类学方法将其转移到软凹藻属中,命名为Chondrophycusundulatus(Yamada) Garbary et Harper。这种处理被后来的学者所接受[28]。波状软凹藻与软凹藻C.cartilagineus在外部形态上比较相似[28],但它们的表皮细胞和囊果存在一定的区别:波状软凹藻的纵切表皮细胞不凸起,且囊果没有突出的喙[28,41];软凹藻的分枝纵切面表皮细胞突起[42],成熟的囊果圆锥形,并有一个突出的孔口[27]。本文样品 (编号:20130427007)虽未观察到囊果,但藻体的表皮细胞不凸起;在分枝纵切面上,表皮细胞栅状排列 (图1h、g、k、l);四分孢子囊末小枝不扁压,截形或瘤状,四分孢子囊散生在截形或瘤状的末小枝端面,与中央轴呈垂直排列 (图1d、e、f);四分孢子囊四面体形分裂 (图1m),这些特征与文献[27-28]中对波状软凹藻的描述一致,因此将其命名为波状软凹藻Chondrophycusundulatus。
凹顶藻属Laurencia于1813年建立,藻体基部多为盘形固着器,少数具有圆柱状的匍匐枝;顶部有一明显凹陷;分枝可呈侧生、轮生、互生、对生和二岐螺旋状等;每个轴细胞产生4个围轴细胞;四分孢子囊与轴平行排列。但随着研究的不断深入,凹顶藻属的物种鉴定由最初依靠藻体颜色和囊果的特征,逐渐转向精细的解剖学特征,如围轴细胞个数、第一个围轴细胞与生毛体位置、精子囊的发育、果胞系母细胞上围轴细胞的数量、受精后的辅助细胞形成时间、四分孢子囊起源和排列方式[19,23,34,43-44]等特征。目前,该属已报道287种及144个亚种,已有138种得到确认[1]。凹顶藻Laurenciachinensis隶属于凹顶藻属,由Tseng[37]于1943年根据香港的样品命名,Tseng认为其藻体在成熟时有显著的扁压主轴和明显的二歧分枝,被认为属于福氏组Forsterianae。福氏组的藻体在幼期主轴为圆柱状,但随着生长,界限逐渐变得模糊,该物种的老枝最终随着时间的增长而变得密集,其轴仅轻微地压缩,并且其主要分枝不规则且紧密地分布,但其藻体明显扁压的特征又符合羽状组的描述,似乎介于Forsterianae与Pinnatifidae之间[37]。丁兰平[28]对其进行研究发现,凹顶藻有稍突出的表皮细胞,与Tseng[37]的描述稍有差别。本文样品 (编号:20130528007,20140518009)藻体也有稍突出的表皮细胞 (图2i、p),且该藻横切清晰可见4个围轴细胞 (图2k),四分孢子囊与轴平行排列 (图2d),符合凹顶藻属的特征,因此,接受其为凹顶藻Laurenciachinensis。
三列凹顶藻Laurenciatristicha隶属于凹顶藻属,由Tseng等[45]于1982年根据香港的样品命名,得到了后续研究者的认同[28,32]。本文样品 (编号:20140518011)的藻体呈浅绿色 (图2a),小枝排列紧密且明显呈三列末小枝和四分孢子囊小枝均为棒状 (图2c、d),四分孢子囊小枝与小枝轴平行排列 (图2f),每轴细胞产生4个围轴细胞 (图2i),囊果壶型 (图2e)。以上这些特征,尤其是小枝呈现明显的三列分岐,与已报道的三列凹顶藻的独有特征一致,因此,接受其为三列凹顶藻Laurenciatristicha。
栅凹藻属Palisada由Nam[22]于2007建立,目前被认为是独立的属,包含29个物种及1个亚种[1]。小瘤栅凹藻Palisadaparvipapillata(Tseng) Nam是由Nam于2007年根据Tseng[37]鉴定为小瘤凹顶藻Laurenciaparvipapillata(产地:香港)的物种修改而来的[22]。其间,这个种的分类地位发生过多次变动,1969年Saito[42]将其归为凹顶藻属软凹藻亚属Chondrophycus;而有学者基于表皮细胞间存在次生纹孔连结[41,46]和四分孢子囊垂直排列[37,41]等特征,认为该物种的亚属位置需重新审议[47-49];1998年Garbary等[20]将其属级地位变动,将软凹藻亚属Chondrophycus提升至软凹藻属,改为小瘤软凹藻Chondrophycusparvipapillatus(Tseng) Garbary et Harper;丁兰平[28]也认可该属级地位的变动。然而,2006年Nam[21]在对凹顶藻类进行系统进化分析时,发现软凹藻属Chondrophycus不是单系类群,并将软凹藻属内的栅状组与软凹藻属分开,列为独立进化枝;随后,2007年Nam[22]将Palisada提升至属级,并将软凹藻属中包括Chondrophycusparvipapillatus(Tseng) Garbary et Harper在内的一些物种移至该属,重新命名为小瘤栅凹藻Palisadaparvipapillata(Tseng) Nam。目前,P.parvipapillata(Tseng) Nam这个命名被国际所接受[1,27,50]。本文样品 (编号:20130528018)的藻体表皮细胞呈栅状排列,末小枝顶部凹陷,有可辨识的中心轴及其两个围轴细胞,这与栅凹藻属的特征符合,因此将其命名为小瘤栅凹藻Palisadaparvipapillata(Tseng) Nam。
4 结论
本文报道了我国广东汕尾凹顶藻类海藻3属4种,即波状软凹藻(Chondrophycusundulatus(Yamada) Garbary et Harper)、凹顶藻 (LaurenciachinensisTseng)、三列凹顶藻 (LaurenciatristichaTseng,Chang,Xia et Xia)和小瘤栅凹藻(Palisadaparvipapillata(Tseng) Nam),并对其现有问题及凹顶藻类海藻的分类系统进行了探讨,以期为凹顶藻类的物种鉴定提供一定的分类学借鉴。
