木霉生物有机肥对白菜和甘蓝产量及土壤微生物区系的影响
2020-07-28刘正洋王若斐乔策策张楠沈宗专李荣沈其荣
刘正洋,王若斐,乔策策,张楠,沈宗专,李荣,沈其荣
(南京农业大学江苏省固体有机废弃物资源化高技术研究重点实验室/江苏省有机固体废弃物协同创新中心/教育部资源节约型肥料工程技术研究中心,江苏 南京210095)
土壤微生物在土壤生产力形成的生物进程中起着关键性作用[1],是土壤生态系统功能持续发挥的重要媒介[2]。在土壤微生态系统中,微生物的活跃程度反映土壤肥力的状况[3]。因此,通过土壤微生物区系调控,提高土壤生产力成为当前研究的热点和难点。研究证实土壤中施入不同的肥料可以改变土壤微生物群落结构[4]。相比于化肥,长期连续施用有机肥和生物有机肥能增加土壤微生物结构和功能多样性[5],提高土壤促生和抑制土传病害的能力[6]。目前,关于生物有机肥的研究主要集中在芽胞杆菌生物有机肥上[7],对于其他微生物有机肥尤其是真菌类生物有机肥对土壤微生物区系调控的研究还不够深入。
木霉是土壤中重要的促生真菌,能通过矿化有机物等多种机制促进作物的生长和发育[8-9],通过产生如抗菌肽类等抗生素类物质抑制病原菌生长,防控植物土传病害[10]。研究表明,木霉通过增加植株中生长素(IAA)的产生量增强植物对盐胁迫的耐受性[11],通过有效控制玉米茎腐病的发生显著提高玉米产量[12]。同时,有研究利用盆栽试验发现,木霉生物有机肥能通过改变土壤理化状况和根际微生物群落组成提高作物产量[13]。然而,田间木霉生物有机肥是如何通过改变土壤理化性质和微生物区系,从而增强作物产量的微生物生态学机制研究仍然不够。
本研究在白菜-甘蓝轮作体系下,通过连续两季生物有机肥的施用,评估用贵州木霉NJAU4742研制的生物有机肥对轮作体系作物产量和土壤理化性状的影响,并进一步通过比较生物有机肥和普通有机肥对土壤微生物区系的影响,探究木霉生物有机肥提高作物产量的土壤微生物生态学机制,以期为设施蔬菜高产栽培提供理论依据。
1 材料与方法
1.1 试验材料
作物分别为白菜‘改良青杂3号’和甘蓝‘H60’。猪粪有机肥由江苏峻德生态农业科技有限公司提供,有机质含量457.1 g·kg-1,全氮含量14.8 g·kg-1,全磷含量25.2 g·kg-1,全钾含量20.1 g·kg-1。生物有机肥则为有机肥中添加5%(质量分数)固体木霉菌种(TrichodermaguizhouenseNJAU4742,功能菌数量≥1.0×109CFU·g-1)。供试菌株为贵州木霉NJAU4742[14],由本实验室保存。
1.2 试验设计
田间试验点位于江苏省南京市蔬菜科学研究所科技园(31°43′N,118°46′E),年平均气温15.4 ℃,平均降雨量为1 106 mm。供试土壤为黄壤土,pH 7.13,有机质含量22.65 g·kg-1,全氮含量4.05 g·kg-1,全磷含量3.58 g·kg-1,全钾含量3.26 g·kg-1,速效磷含量280.3 mg·kg-1,速效钾含量251 mg·kg-1。田间试验于2015年9月至2016年6月,在白菜-甘蓝轮作体系下连续种植2季。设置施有机肥(OF)和木霉生物有机肥(BIO)处理。每个处理3个小区重复,每个小区面积为5.4 m2(2.7 m×2 m),小区间隔0.6 m。每季大田OF和BIO处理按照等养分施肥,全氮(TN)含量为222 kg·hm-2,全磷(P2O5)含量为150 kg·hm-2,全钾(K2O)含量为180 kg·hm-2,有机肥施用量为6 000 kg·hm-2,其余养分用尿素、过磷酸钙和硫酸钾按总养分补齐;肥料分2次施用,种苗前施入总养分的2/3作为基肥(包括有机肥或生物有机肥及相应养分的化肥),剩余的1/3养分于初果期追肥施用。
1.3 土壤样品采集
于每季白菜和甘蓝收获期,在各小区分别采集8棵长势健康的白菜和甘蓝,连根带土挖出,装入无菌自封袋带回实验室,用手轻抖根系,将抖下来的土壤定义为土体土壤。土体土壤一部分自然风干后过筛用于理化性质的测定,另一部分-80 ℃保存待用。
1.4 测定方法
1.4.1 土壤理化测定参照文献[15],分别采用玻璃电极酸度计和电导仪测定土壤pH和EC值;采用醋酸铵浸提-火焰光度计法测定土壤速效钾含量;采用碳酸氢钠浸提-钼蓝比色法测定土壤速效磷含量;采用氯化钙浸提-流动分析仪法测定土壤硝态氮和铵态氮含量。
1.4.2 土壤DNA的提取称取0.25 g土壤样品,根据DNA提取试剂盒(Mo Bio Laboratories,USA)操作说明提取土壤DNA。以核酸定量仪(NanoDrop 2000,USA)测定土壤DNA浓度,稀释至20 ng·μL-1待测序。
1.4.3 PCR反应和MiSeq高通量测序参照Li等[16]的方法,细菌16S rDNA的扩增引物为 520F(5′-AYTGGGYDTAAAGNG-3′)和 802R(5′-TACNVGGGTATCTAATCC-3′),真菌ITS的扩增引物为ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和 ITS2(5′-GCTGCGTTCTTCATCGATGC-3′)。PCR反应体系25 μL:灭菌超纯水9.0 μL,反应缓冲液(5×)5.0 μL,PCR增强添加剂(GC high enhancer)(5×)5.0 μL,dNTP(2.5 mmol·L-1)2.0 μL,模板DNA 2.0 μL,引物F(10 μmol·L-1)1.0 μL,引物R(10 μmol·L-1)1.0 μL,TaKaRa聚合酶(5 U·μL-1)0.25 μL。PCR扩增、文库准备及上机测序程序均委托上海派森诺生物科技股份有限公司完成。
1.4.4 MiSeq测序的序列处理与分析利用Mothur Version 1.25.1进行数据处理,流程为:1)对每个样品的正、反向序列进行拼接,并分配样品名称;2)去除q值大于0.5的序列、长度小于200 bp的序列、重复序列、单序列等劣质序列;3)生成操作分类单元(operational taxonomic unit,OTU),选取的阈值水平为97%;4)去除嵌合体;5)生成OTU表格,得到OTU的代表序列和Read数;6)将16S rDNA基因和ITS序列的每个OTU对应的序列通过RDP数据库比对,确定每个OTU的分类地位,细菌选用数据库为RDP 16S rDNA Gene Database,真菌选用数据库为UNITE Fungal ITS Trainset。
将OTU表格转换成Mothur软件能够识别的Shared格式文件,对样品均一化处理后,计算测序样品各处理的α多样性,比较样品之间的丰富度(Chao指数)和多样性(Shannon指数)差异。利用Mothur软件对所有样品在OTU水平下基于Bray-curtis距离进行主坐标分析(principal coordinates analysis,PCoA)。利用R version 3.5.1软件对测序样品进行方差分解分析(variance partitioning analysis,VPA),研究驱动群落变化的因子。利用冗余分析(redundancy analysis,RDA)探究不同处理、环境因子与微生物群落组成之间的关系。利用线性判别分析效应大小(linear discriminant analysis effect size,LEfSe),分析不同处理属水平下相对丰度具有显著性差异的微生物。利用皮尔逊(Pearson)相关分析找出与产量显著相关的属。
1.5 数据统计与分析
数据统计分析采用Excel 2010 和SPSS 20.0软件。数据间的多重比较选取单因素方差分析(One-way ANOVA),用R version 3.5.1软件进行微生物群落多样性分析,显著性水平设定为P<0.05。
2 结果与分析
2.1 不同施肥处理对白菜-甘蓝轮作体系产量的影响
由表1可知:第1季(白菜季)BIO处理相比OF处理白菜产量虽无显著性差异,但增产4 725 kg·hm-2,增幅达4.25%。第2季(甘蓝季)BIO与OF处理产量差异显著,相比OF处理,BIO处理甘蓝产量增加9 830 kg·hm-2,增幅达8.85%。
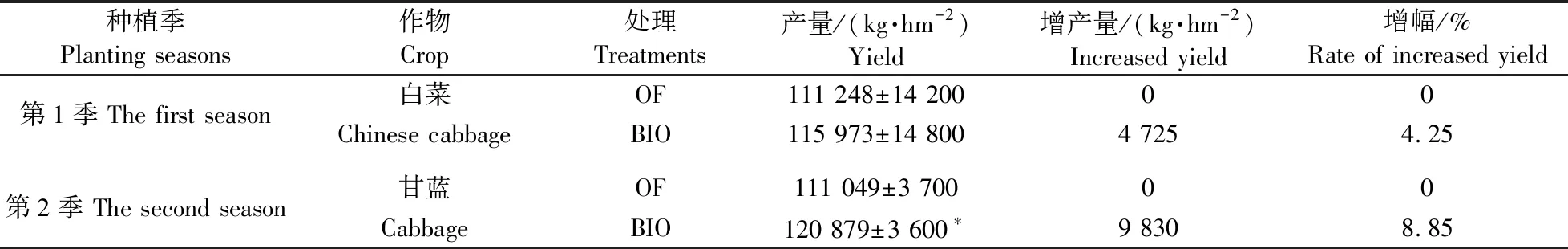
表1 不同施肥处理白菜和甘蓝的产量Table 1 The Chinese cabbage and cabbage yields of different fertilization treatments
2.2 不同施肥处理对白菜-甘蓝轮作体系土壤理化性质的影响
由表2可知:第1季BIO处理相比OF显著提高土壤EC值和速效钾含量,显著降低土壤硝态氮含量。第2季BIO处理相比OF显著增加速效钾和硝态氮含量,显著降低pH值。在2季中,BIO处理相比OF处理提高土壤EC值、速效钾和速效磷含量,降低pH值。

表2 不同施肥处理的土壤理化性质Table 2 Soil properties of different fertilization treatments
2.3 不同施肥处理对土壤微生物群落α多样性的影响
由表3可知:在施肥处理方面,除第2季中细菌群落的Chao指数外,BIO处理微生物群落的Chao指数和Shannon指数均大于OF处理,但各处理间无显著性差异;在种植季方面,BIO处理第2季细菌的Chao指数和Shannon指数均大于第1季,第2季真菌的Chao指数和Shannon指数均小于第1季,OF与BIO处理有相同趋势。

表3 不同施肥处理细菌和真菌的Chao和Shannon指数Table 3 Bacteria and fungi Chao and Shannon indexes of different fertilization treatments
2.4 不同施肥处理对土壤微生物群落β多样性的影响
由图1可知:PCoA1和PCoA2分别解释细菌群落变化的30.02%和18.56%,真菌群落变化的27.09%和17.99%,其中细菌群落第1季与第2季沿PCoA1区分,BIO和OF处理沿PCoA2区分。真菌群落第1季与第2季沿PCoA1区分,BIO和OF处理沿PCoA2区分。
2.5 不同施肥处理土壤微生物群落方差分解分析
方差分解分析(VPA)结果显示,土壤理化性质(速效钾含量、速效磷含量、硝态氮含量和铵态氮含量)、种植季(第1季和第2季)、功能微生物(贵州木霉NJAU4742)及其交互作用分别解释了总变异的32.98%(细菌)和35.61%(真菌)(图2)。解释度最高的因子是功能微生物,分别占细菌和真菌总变异的8.41%和5.21%,而细菌和真菌总变异的7.34%和4.10%归因于土壤理化性质。种植季对细菌和真菌群落组成的影响较小。未解释的细菌和真菌群落分别为67.02%和64.39%。
2.6 不同施肥处理对土壤微生物群落组成的影响
通过线性判别分析的效应大小对不同处理属水平下相对丰度具显著性差异的微生物分析(图3)可知:BIO处理的2季土壤中伯克氏菌属(Burkholderia)和Bhargavaea属的相对丰度均显著高于OF处理(图3-A、C);木霉属(Trichoderma)和红酵母属(Rhodotorula)的相对丰度均显著高于OF处理(图3-B、D)。
2.7 微生物相对丰度与产量的关系
属水平上,产量与微生物的相关性如表4所示。BIO处理土体土壤中伯克氏菌属(Burkholderia)和Bhargavaea属的相对丰度与产量显著正相关(P<0.05);BIO处理土体土壤中木霉属(Trichoderma)和红酵母属(Rhodotorula)的相对丰度与产量显著正相关(P<0.05)。

表4 产量与微生物的相关性Table 4 Correlations between yield and microbial community
2.8 不同施肥处理土壤微生物群落冗余分析
细菌群落冗余分析(图4)结果显示:RDA1和RDA2共解释了81.42%的细菌群落变化;真菌群落的RDA1和RDA2解释的比例为93.62%。RDA1将OF和BIO处理微生物群落明显分开,RDA2将各处理第1季和第2季微生物群落明显分开。BIO处理第1季细菌和真菌群落均与产量、功能微生物、EC、速效钾含量和速效磷含量呈正相关;第2季都与产量、功能微生物、EC、速效钾含量、速效磷含量、铵态氮含量和硝态氮含量呈正相关。BIO处理细菌群落主要与伯克氏菌属(Burkholderia)和Bhargavaea属聚合在一起;BIO处理真菌群落主要与木霉属(Trichoderma)和红酵母属(Rhodotorula)聚合在一起。
3 讨论
连续2季田间试验结果表明,相比于有机肥,生物有机肥有效增加白菜-甘蓝轮作体系产量,推断是由于生物有机肥中含有大量的功能菌——贵州木霉NJAU4742(TrichodermaguizhouenseNJAU4742)[17]。已有研究报道贵州木霉NJAU4742能产生多种促进作物生长的活性物质[18]。Cai等[19]利用盆栽试验发现用贵州木霉NJAU4742研制的生物有机肥能有效促进番茄生长。哈茨木霉能在作物根际定殖并显著增加根系表面积,在养分胁迫的条件下,提高作物对土壤中磷和其他养分的吸收能力,促进作物生长[20]。本研究利用田间试验进一步证实木霉生物有机肥的田间增产效果。同时,还发现木霉生物有机肥能够提高土壤中速效养分的含量,木霉能促进番茄产生大量的根系分泌物,增加根际微生物的种群和活性,释放土壤中缓效氮和磷养分,促进番茄生长[21]。高通量测序已经成为研究微生物群落的主要方法之一[22]。本研究应用MiSeq测序技术对白菜-甘蓝轮作体系不同施肥处理和种植季土壤细菌和真菌群落进行分析,结果表明生物有机肥能提高微生物群落的多样性。Qiu等[23]研究发现施用生物有机肥能提高土壤中细菌和真菌群落的多样性,降低土壤中尖孢镰刀菌的数量,提高黄瓜产量。Shen等[24]研究发现施用生物有机肥能通过提高土壤细菌群落多样性的方式防控香蕉枯萎病并提高产量。因此,推测增产效果与微生物群落多样性的提高有关,这与徐丽霞[25]的研究结果类似。
基于PCoA分析发现,相对于有机肥处理,生物有机肥处理的土壤微生物群落组成差异显著。已有研究表明施用不同种类的有机肥能影响土壤微生物群落的组成[26]。Zhao等[27]发现施肥方式是改变土壤细菌群落结构的主要因子;Zhang等[28]也报道生物有机肥显著改变黄瓜根际土壤真菌群落组成,与本研究相一致。本研究通过方差分解分析发现在白菜-甘蓝轮作体系中,功能微生物是改变土壤微生物群落结构的主要因子。在两季的种植过程中,伯克氏菌属(Burkholderia)、Bhargavaea属、木霉属(Trichoderma)和红酵母属(Rhodotorula)的相对丰度在生物有机肥处理中显著高于有机肥处理,且与产量显著正相关。其中,伯克氏菌属(Burkholderia)具有溶磷作用,从而释放土壤养分[29],而细菌Bhargavaea属对产量的促进作用可能与其能够抵抗盐分胁迫有关[30]。有研究表明红酵母属(Rhodotorula)真菌能够保护甜菜免受丝核菌的侵害[31]。另外,木霉属微生物具有促生作用[32-33]。因此,木霉生物有机肥除木霉的促生作用外,推测其激发的其他土著功能微生物同样也起到促生作用。因此,连续施用木霉生物有机肥能调节土壤微生物群落结构和组成,提高白菜-甘蓝轮作体系产量。
