沙化草地三种牧草叶片生理适应特性的比较研究
2020-07-28周青平孙万斌陈有军
张 伟,周青平,孙万斌,王 平,陈有军
(1.西南民族大学青藏高原研究院,四川成都 610041;2.甘肃农业大学草业学院,甘肃兰州 730070)
青藏高原素有世界屋脊之称,是我国重要的生态屏障,也是全球海拔最高、气候条件独特的地理单元[1].随着社会的发展,受放牧、气候等综合因素的影响,青藏高原高寒草地沙化状况日趋严重,在沙化生境条件下,植物长时间遭受高温、高寒、高旱、盐碱等逆境胁迫,迫使植物的光合作用、生理酶活性和渗透调节物质积累发生改变,导致光合有效积累物质降低,很容易使植物在短期内丧失繁殖潜能,甚至死亡,最终导致草地多样性的减少[2-4].
叶片是植物形态结构的重要组成部分,从自然界获取能量的组织结构,其形态对环境的响应特征直接影响着植物的物质和能量交换[5-6],叶片对环境变化也有极强的敏感性[7-8],长期以来,植物生理学家和植物生态学研究人员对逆境条件下植物叶片的适应性研究保持着高度的关注[9-10],我国的一些研究者从叶片的生理性状特征等角度对叶片形态变化及其适应特征做了很多研究,发现植物叶片适应性的形成是环境长期作用的结果,保证了植物自身完成生活史周期,也是适应环境的重要生理生态特性[11-16].植物通过调整叶片的光合特征、抗氧化酶保护系统的启动、渗透调节剂的积累等途径来影响自身的生长动态,重新构建代谢平衡[17-18].
受到高温逆境条件的影响,叶片的光合系统、细胞膜系统、叶绿体超微结构、保护酶系统都受到了不同程度的破坏[19-20],沙化逆境下植物的生长状况是对环境因素高温、高寒、高旱、盐碱等胁迫的综合表现[21-22],在这种条件下,以单一的生理指标评估来自不同植物适应逆境的能力会出现较大的误差,因此,通过比较叶片的光合特性、光响应曲线、生理酶活性等变化,可以综合说明其对沙化生境的适应性.
目前,人工修复是沙化草地治理的重要措施之一,近些年来,我们在开展高寒地区沙化草地治理工作中发现,以礼草(Koegneria hirsute)和赖草(Leymussecalinus)是该生境条件下的先锋牧草,对沙化生境有着很强的适应性,垂穗披碱草(Elymusnutans)是高寒草地上的主要建群种和优势种之一,是被常用来恢复高寒沙化草地的主要草种之一[23]. 本试验以沙化草地采用人工种植以礼草、赖草和垂穗披碱草为研究对象,测定叶片光合指标、光合曲线、生理酶活性等,比较分析了叶片的生理特征,旨在进一步比较三种牧草在沙化逆境条件下的适应性,为沙化草地恢复中的引种栽培、遗传改良和新品种选育提供科学依据,也为高原地区生态治理、退化草地恢复牧草的选择提供参考.
1 材料与方法
1.1 试验材料
为保证取样地点的外界生长条件和样本的一致性,首先在试验地选取2017 年人工种植的以礼草、赖草和垂穗披碱草,随机设置12 个1 m ×1 m 的样方,在样方内选择花期的样本定株取样,4 个重复(表1).

表1 三种牧草基本情况Table 1 The basic information of three forages
1.2 试验地概况
试验地位于四川省阿坝藏族羌族自治州红原县瓦切乡(N:31°51′~33°19′,E:101°51′~103°23′),海拔3 210 ~4 857 m,属大陆性高原寒温带季风气候,春秋短促、长冬无夏.年均降雨量791.95 mm,降雨主要集中在5 ~9 月,年均气温为1.1 ℃,最冷月平均气温10.3 ℃,最热月平均气温10.9 ℃,极端最低气温-36 ℃,年均积雪期为76 d,无绝对的无霜期. 日照充分,太阳辐射强,年均日照时间2 158.7 h,太阳辐射年总量为6 194 MJ·m-2.
1.3 试验方法
1.3.1 光合特性
(1)光合指标的测定
开花期选晴天早上10:00 -11:30,使用Li-6400便携式植物光合作用测定仪,测定样本的光合速率(Pn)、蒸腾速率(Tr)、水分利用率(WUE)、气孔导度(Gs)、胞间CO2浓度(Ci).每个种选取4 株大小相近、生长良好的测定样本,测定时选择旗叶测定.4 次重复,取平均值[24].
(2)光响应曲线
完成光合指标的测定后,用Li -6400 便携式光合作用测定系统(Li -Cor,lincoln,USA)测定光合响应曲线.测定时,使用2 cm ×3 cm 的红蓝光源叶室,设置光照强度梯度为 0、70、150、250、400、600、1000、1500、2000 μ mol·m-2·s-1,测定不同光照强度下三种牧草相应的净光合速率.
参照二氧化碳浓度设定为400 μ mol·mol-1,沙化草地的地表温度较高,试验控制温度选为30℃. 每种牧草取3 株作为重复,选取旗叶测定,取其平均值.光响应曲线的相关参数利用Walker 所提出的相关模型及净光合速率与光合有效辐射拟合的二次方程进行求导计算,其模型函数表达式为(1):

公式(1)中,A 为净光合速率(μ mol·m-2·s-1);Q 为表观量子效率(mol·mol-1);Amax 为最大光合速率(μ mol·m-2·s-1);Rd 为暗呼吸速率(μ mol·m-2·s-1);Par 为光合有效辐射(μ mol·m-2·s-1);K 为曲角. 对模型中各个相关参数选用SPSS统计分析软件中的非线性回归方法估计,并对光响应曲线的初始部分Par 进行线性回归,光补偿点(LCP,μ mol·m-2·s-1)即为它与X 轴的交点;光饱和点(LSP,μ mol·m-2·s-1)的计算则利用 A 与 PAR 拟合的二次方程求导得到[25-27].
1.3.2 生理指标的测定
在划定的样方中,选取长势均一的三种牧草旗叶,采集的样品放入封口的塑料袋内,并将其储藏在黑暗的保温箱内(有冰袋,内部温度<4℃),样品设3个重复;将其带回实验室并快速进行相关指标测定,其它生理指标的样品采用用锡纸包装,并将样品装于纱布袋中储存在液氮罐中,带回实验室储存于-80℃超低温冰箱中待测[28].测定内容包括:丙二醛(Malondialdehyde,MDA)、可溶性蛋白(Soluble protein,SP)、超氧化物歧化酶活性(Superoxide Dismutase)、过氧化氢酶(Catalase Test,CAT)、抗坏血酸过氧化物酶(Aseorbateperoxidase,APX)、过氧化物酶活性(Peroxidase,POD),叶片的上述生理指标分别采用试剂盒测定[29].
1.4 叶片抗性综合评价方法
采用Fuzzy 数学中隶属函数法对供试材料的6 个生理生化指标进行适应性综合评判[30-31],计算公式如下:
可溶性蛋白、超氧化物歧化酶活性、过氧化氢酶、抗坏血酸过氧化物酶、过氧化物酶活性与适应性呈正相关,用公式(2)计算:

MDA 与适应性呈负相关,用公式(3)计算:
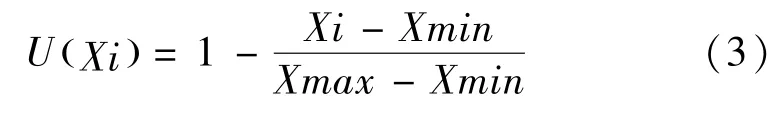
上述(2)和(3)式中:Xi 为指标测定值. Xmax 为参试材料某一指标的最大值,Xmin 为参试材料某一指标的最小值.用供试材料的各项测定指标的隶属度平均值来作为供试材料对沙化生境条件下的综合评判标准.
1.5 数据处理
本试验中的数据用Spss 22.0(Statistical Product and Service Solutions)统计软件进行统计分析,采用Origin2017 绘图.
2 结果与分析
2.1 光合特性分析
由表2 可知,以礼草光合速率Pn 值为31.55 μ mol·m-2·s-1,显著高于赖草和垂穗披碱草(P<0.05);垂穗披碱草蒸腾速率值最高,其次是以礼草、赖草,三者差异显著(P<0.05).赖草叶片的水分利用率为3.74 μ mol·mol-1,显著高于以礼草和垂穗披碱草,且三者差异显著(P<0.05).三种牧草的气孔导度值介于0.55~0.63m mol·m-2·s-1之间,无显著性差异.赖草的胞间CO2浓度显著高于以礼草和垂穗披碱草,差异显著(P<0.05),达到了 248.95 μmol·mol-1,其次是以礼草,为226.65 μmol·mol-1,垂穗披碱草最低,两者之间差异不显著(P>0.05).

表2 三种牧草的光合指标比较Table 2 Comparison of photosynthetic indexes among three forages
2.2 光响应曲线分析
设置外源光照强度(Par)梯度为 0、70、150、250、400、600、1 000、1 500、2 000 μmol·m-2·s-1条件下,测定最大光合速率(Pn);对Par、和Pn 进行非直角双曲线模型了模拟,模拟结果看出,三个方程的决定系数均在0.85 以上,结果能较好的反映3 种植物叶片光合对光照的响应过程.
随着Par 值由小到大的梯度变化,3 种牧草的净光合速率呈抛物线形变化,净光合速率随着光合有效辐射的增大均表现出先增大后减小的趋势,峰值大小不同. 以礼草的峰值较大,出现在光合有效辐射为1 560.38 μmol·m-2·s-1的时候,垂穗披碱草最小,出现在1 664.25 μmol·m-2·s-1,1 854.93 μmol·m-2·s-1 的时候. 当 Par 值 > 900 μmol·m-2·s-1时,三种牧草的净光合速率最大值分别为:以礼草>赖草>垂穗披碱草(图1).
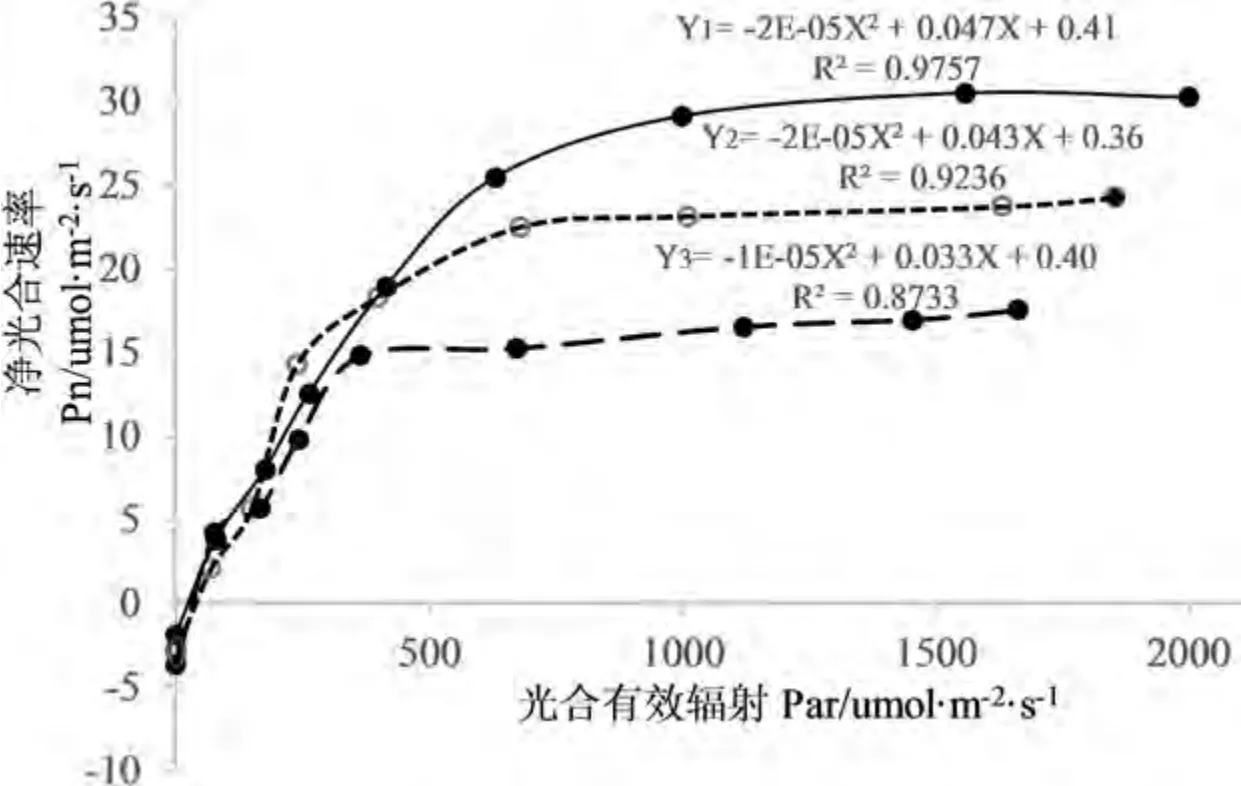
图1 三种牧草的光响应曲线Fig.1 Light response curve of three forages
比较光响应参数可以看出(表3),以礼草的最大光合速率显著高于赖草和披碱草,达到了31.85 μmol·m-2·s-1;其次是赖草,垂穗披碱草最低,仅为19.17 μmol·m-2·s-1,三者之间差异显著(P<0.05).光合曲角和表观量子效率三者之间无显著差异(P>0.05).以礼草的暗呼吸显著低于赖草和垂穗披碱草(P<0.05),仅为 1.25 μmol·m-2·s-1;赖草和垂穗披碱草之间,差异不显著(P>0.05). 以礼草的光饱和点最高,为 1 307.56 μmol·m-2·s-1,其次是赖草为934.88 μmol·m-2·s-1,垂穗披碱草最低,仅有582.48 μmol·m-2·s-1,三者之间差异显著(P<0.05). 赖草和垂穗披碱草的光照补偿点分别为33.39 μmol·m-2·s-1、39.94 μmol·m-2·s-1,无显著差异,但是两者数值显著高于以礼草(P>0.05).
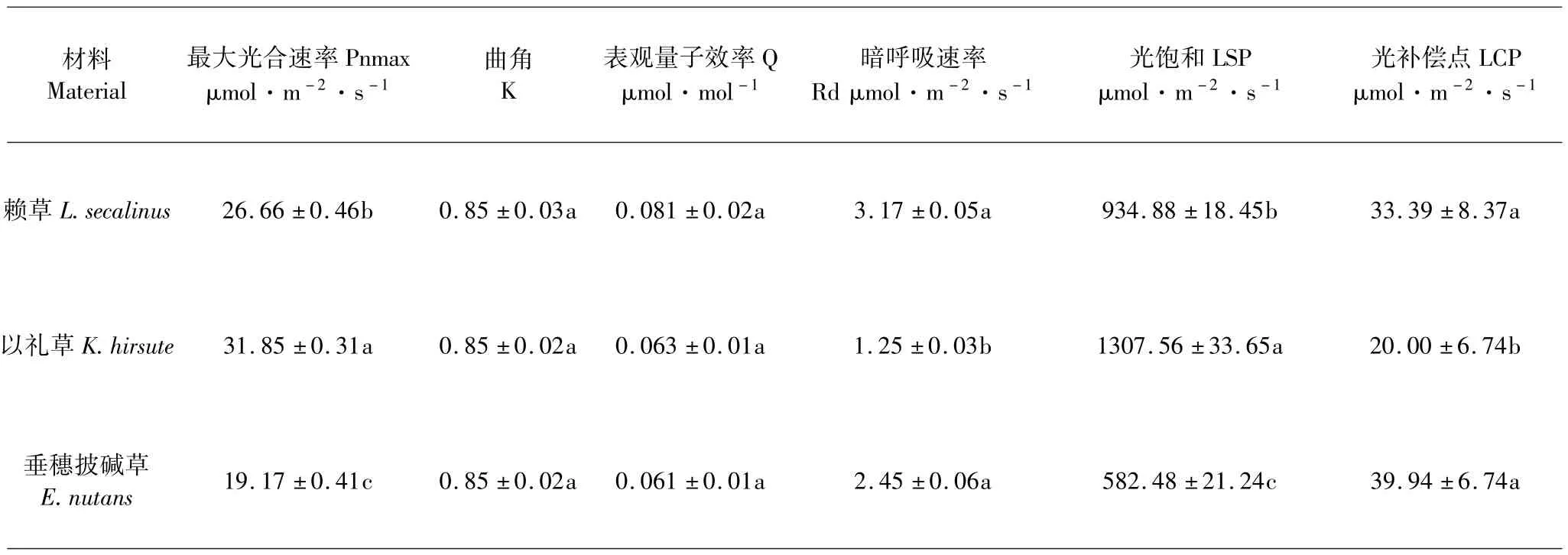
表3 三种牧草光响应参数比较Table 3 Comparison of the light response model parameters of three forages
2.3 抗逆生理指标分析
2.3.1 生理指标测定
以礼草的MDA 含量显著低于赖草和垂穗披碱草(P<0.01),为 21.0 m mol·g-1,赖草和垂穗披碱草之间无显著差异(P>0.05). 赖草、以礼草的可溶性蛋白含量分别为 52.6 mg·g-1、58.3 mg·g-1,均显著高于垂穗披碱草. 赖草的SOD 值为42.3 μ·g-1,显著高于以礼草和垂穗披碱草(P<0.05),垂穗披碱草的含量最低(P<0.05).三种牧草的CAT 活性无显著差异,保持在0.4 ~0.6 μ·g-1之间.以礼草的APX值为4.7 μ·g-1,显著高于赖草和垂穗披碱草(P<0.05).比较POD 的含量发现,赖草的值为307.3 μ·(min·mg)-1 显著高于以礼草和垂穗披碱草(P<0.05). 以礼草和垂穗披碱草之间无显著差异(P>0.05)(图2).
2.3.2 叶片抗性综合评价
用隶属函数法,比较两个沙化草地先锋牧草的抗性强弱.三种牧草测定指标参数的隶属函数值由上述公式(1)和(2)可以求出,将各项指标的隶属函数值相加,求平均值,得出抗性综合评价值.供试材料的综合评价值越大,抗性越强,反之则越弱.综合评判分析发现,在沙化生境条件下,三者抗性综合表现顺序依次为:以礼草>赖草>垂穗披碱草(表4).

图2 三种牧草叶片的生理指标比较Fig.2 Comparison of physiological indexes of three forage leaves

表4 三种牧草叶片抗性综合评判Table 4 Evaluation of leaf resistance between three forage species
2.4 光合指标和酶活性相关性分析
分析了三种牧草叶片最大光合速率、光合补偿点、光合饱和点与抗性相关的酶活性、渗透调节物质之间的相关性.表5 可以看出,植物的最大光合速率与 MDA、CAT 含量呈负相关,差异不显著(P>0.05);与 SOD、蛋白含量、CAT、APX、POD 呈正相关,其中与SOD 差异极显著(P<0. 01). 光的饱和点和 MDA、SOD、可溶性蛋白含量、APX、POD 的相关性呈正相关,其中与 SOD 差异极显著(P<0.01);与 CAT 呈负相关,差异不显著(P>0.05). 光的补偿点与MDA、SOD、APX、POD 呈负相关,其中与 SOD 差异极显著(P<0.01),;与蛋白含量、CAT 含量呈正相关,其中,CAT 差异显著(P<0.05).

表5 叶片光合指标和生理酶活性的相关性比较Table 5 Correlation between leaf photosynthetic index and physiological enzyme activity
3 讨论
3.1 三种牧草的光合指标比较
沙化草地具有高寒、干旱、高温等特殊的交叉胁迫影响着植物的正常生长,沙化地植物通过叶片的光合酶系统及生理调节适应这种特殊的生境[32]. 先锋牧草的以礼草净光合速率值最高,净光合产物积累能力强,王云贺等人研究结论也证明了野生植物净光合速率高于栽培种[33]. 赖草叶片的水分利用率、胞间CO2浓度、气孔导度值表现最大,在该生境下具有更好的CO2和水分的捕捉能力,保持了植株高大、单株鲜重大等特性,李莹等人的研究验证赖草的这一特性[34].作为高原地区常见的栽培种,垂穗披碱草叶片的蒸腾速率最大,叶片更多的通过蒸腾作用去适应这种环境,Burgess 等人的研究结论也表明,受到环境的变化影响,植物通过改变叶片的内外蒸气压,加快蒸腾速率,造成的水势梯度来吸收和运输水分,促进植物的生长发育[35].
光合光响应曲线可以反映植物在生长环境条件下对光的利用方式和对环境的响应[36].以礼草的Pn、Pnmax、LSP 显著高于赖草和垂穗披碱草,Rd、LCD 显著低于赖草和垂穗披碱草,有学者研究发现,紫叶小檗(Berberisthunbergii)的Pnmax 值最大,其叶片在光照有限的条件下进行光合作用能力最强,促进了有机物质的积累[37-38]. 研究小麦(Triticumaestivum)、大豆(Glycine max)、玉米(Zea mays)和千穗谷(Amaranthushypochondriacus)发现,Rd 值低有利于光合产物的积累[39],植物利用弱光的能力可以用光补偿点(LCP)进行反映,其数值越低表明植物利用弱光的能力越强;植物利用强光的能力可以用光合饱和点(LSP)来反映,其值越大表明利用强光进行光合作用能力强[40-41],进一步证明了以礼草对光环境的适应范围较广,对强光和弱光均有很好的利用能力,保持叶片自身的最大光合能力来适应生境条件,赖草高大,叶片较高的LSP 和Pnmax 是其适应这种生境的生理特性.三种牧草的光合特性差异大,这可能是植物不同适应环境的方式不同[41].
3.2 三种牧草的生理活性指标比较
以礼草和赖草的叶片MDA 含量较垂穗披碱草低,徐萍等研究干旱胁迫对银沙槐(Ammodendronargenteum)幼苗发现,植物受到轻度胁迫时,自身叶片细胞膜稳定性增强,随着胁迫的程度增加,细胞膜发生膜脂过氧化,MDA 含增加,导致膜的稳定性降低,膜透性增大[42],以礼草和赖草的叶片细胞膜稳定性较强.匍匐翦股颖(Agrostisstolonifera)在干旱胁迫条件下,通过提高自身叶片细胞的渗透调节物质的含量来适应胁迫,高含量的可溶性蛋白等渗透调节物质使植物细胞的渗透势维持在较低的水平下,进而可以抵御水分胁迫对植物带来的伤害[43],以礼草和赖草的叶片细胞可溶性蛋白含量高,其叶片通过提高渗透调节物质的含量使其自身细胞维持在一个较低的渗透势,减少逆境造成的伤害[44].
植物受到逆境胁迫信号后,通过调节SOD、CAT、POD 和APX 等抗氧化酶维持自身活性氧代谢的平衡,减缓细胞内活性氧自由基积累伤害[45].SOD 催化超氧阴离子自由基的歧化反应,CAT、POD 和 APX 主要清除H2O2[46].SOD、POD、CAT 等酶活性与植物抗逆性呈正相关或负相关存在一定争议[47].
本研究中通过隶属函数综合比较抗逆性的结果来看,作为先锋植物,以礼草叶片在沙化生境中叶片细胞膜稳定性、渗透势水平和抗氧化酶维持自身活性氧代谢的平衡等综合能力最强,这与在花椒(Zanthoxylumbungeanum)[48]、野生大豆[49]、葎草(Humulusscandens)[50]和小麦[51]中的结论类似,证实了SOD、POD、CAT 的活性增强有助提高植株的抗逆性.
3.3 三种牧草的光合指标和酶活性相关性比较
对牧草叶片的光合指标和酶活性相关性比较发现:光的饱和点和叶片的SOD 酶活性呈极显著正相关,光的补偿点与SOD 酶活性差异极显著负相关,与CAT 酶活性差异显著正相关.这与西瓜(Citrulluslanatus)幼苗[52]的研究结果一致,与烟草(Nicotianatabacum)的研究结论不一致,主要原因可能是植物生长环境和植物自身遗传差异导致了不同生境条件叶片光合路径的变化和叶片生理调节的差异[53].
4 结论
以礼草和赖草的叶片具有较强的光合特性,以礼草叶片生理指标综合适应性最强,具有较大的LSP 、Pnmax 和较小的LCP、Rd,对光的适应范围广.三种牧草叶片的SOD 活性显著影响叶片的LSP 和LCP,证实了先锋牧草叶片生理特性对其在沙化生境适应性方面的重要作用.本文仅对沙化草地三种牧草叶片的生理酶活性和光合特性的强弱进行了测定和比较,为下一步开展沙化草地生境条件下叶片的光合特性和相关酶活性生理应答机制方面的研究奠定了基础.今后利用分子手段探究和挖掘以礼草叶片光适应范围广的性状,可以为沙化草地治理培育抗逆牧草新品种发挥重要作用.
