有机废弃物降解过程酶作用及其调控机制的研究进展
2020-07-27张勇,陈骥,张锋
张 勇,陈 骥,张 锋
(1.甘肃农业大学草业学院,甘肃 兰州 730070;2.安徽大学资源与环境工程学院,安徽 合肥 230601;3.Aarhus University Centre for Circular Bioeconomy, Department of Agroecology, Aarhus University, 8830 Tjele, Denmark;4.西北工业大学生态与环境保护研究中心,陕西 西安 710072)
随着社会经济的不断发展,人类生产生活过程中产生的有机废弃物(作物残体、畜禽粪便、生活垃圾、活性污泥等)急剧增加[1-2]。目前,我国总有机废物排放量已超过40亿t,由于处置不当或不彻底(焚烧、填埋、丢弃等),不仅浪费资源和能源,还造成环境污染和温室效应等恶劣生态环境问题[3-4]。例如,作物残体大量焚烧后排放的气体污染物会引起空气污染,增加公众健康风险,加剧气候变化[4];畜禽粪便施用后残留的重金属和抗生素等污染物进入土壤-植物-水体系统,通过食物链富集、系统耦合而引发生物毒性反应[5]。诸多生态环境问题日渐凸显,阻碍了我国生态文明建设和乡村振兴战略进程。因此,有机废弃物资源化利用迫在眉睫。
其实我们理性看一下这个回应,我国没有要求瑞典方道歉,因为外交交涉并不是强制他国给予当事人法律“后果”(比如给个处分、开除公职、罚款判刑之类),这是不尊重他国的主权的行为,我国自然不齿做这种事情。
有机废弃物蕴含着丰富的有机质(木质素、纤维素、蛋白质、脂肪等),生物可降解性强[3]。本着减量化、资源化、无害化原则,生物降解是实现有机废弃物资源化利用的有效途径之一[6-7]。微生物和酶主导的生物降解通常分为变质(biodeterioration)、矿化(mineralization/biofragmentation)和腐殖化(humification/assimilation)3个过程[7]。变质、矿化和腐殖化协同作用,相互影响。变质过程:微生物作用与非生物作用共同改变有机废弃物表面的理化性质,削弱其结构稳定性[7];矿化过程:酶催化大分子有机质解聚为低聚物、二聚物和单体,同时生成盐类、CO2、NH3、H2O等产物[7];腐殖化过程:酶催化矿化过程的小分子中间产物再合成,形成腐殖质(主要成分为胡敏酸、富里酸和胡敏素等分子)和酶-腐殖质复合物,同时微生物代谢产生能量和代谢产物[7-9]。酶是一系列生物化学反应的催化剂,驱动着生物降解过程的矿化和腐殖化[8,10-11],在有机废弃物降解过程中发挥着关键作用,有效控制酶作用对实现有机废弃物资源化利用具有重要意义。
研究显示,酶作用受到酶种类和酶活性的影响[12]379-484。一方面,酶具有特异性,每种酶只作用于特定结构的底物、一类化合物或化学键;另一方面,酶具有高效性,能降低反应活化能而加快反应速率,且酶活性越高其催化能力就越强。然而,尚未有综述系统阐明如何调控生物降解过程的酶作用。深入研究有机废弃物降解过程的酶作用及其调控机制,不仅能全面认识生物降解的途径与分子机制,还能推进酶降解技术的开发和应用。该文综合分析国内外有机废弃物降解方面的研究成果,以期:(1)探讨酶对有机质中碳、氮、磷、硫等元素的降解转化过程,以及对重金属、抗生素毒性的削减作用;(2)总结生物降解过程中酶作用的调控机制;(3)展望有机废弃物降解酶研究的挑战和机遇。
1 有机废弃物降解过程的酶作用
生物酶主要来源于微生物、动植物活体及其残骸,是一类生物活性物质(蛋白质或RNA)[13],具有高度催化能力及温度和pH敏感性[12]379-483。它驱动着生物化学过程的物质转化和能量流动[14-15],其活性能表征有机废弃物生物降解的速度和进程。光谱分析、荧光标记、同位素示踪、酶试剂盒和色谱分析等[16]方法被广泛应用于酶活性测定,能满足大部分实验需求;近几年,生物芯片、单分子荧光成像、纳米粒等技术也被用于酶测定,进一步加强了对酶机制和应用的研究[17-19]。有机废弃物降解酶以水解酶和氧化还原酶为主,大部分来源于细菌和真菌,也有一小部分来源于动植物残体(表1[20-44])。有机废弃物降解矿化和腐殖化过程的潜在分子机制见图1,其酶作用可从催化降解有机质、促进腐殖质合成和削减污染物毒性3个方面探讨。
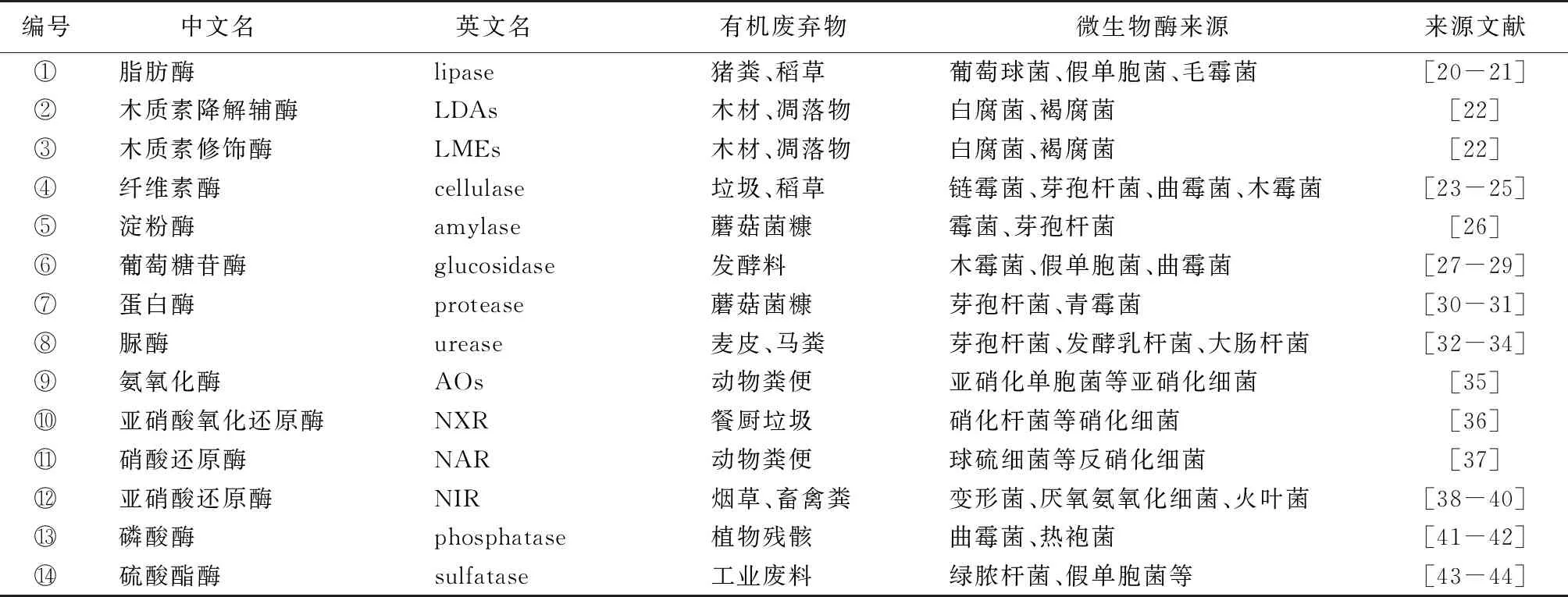
表1 参与有机废弃物降解的酶
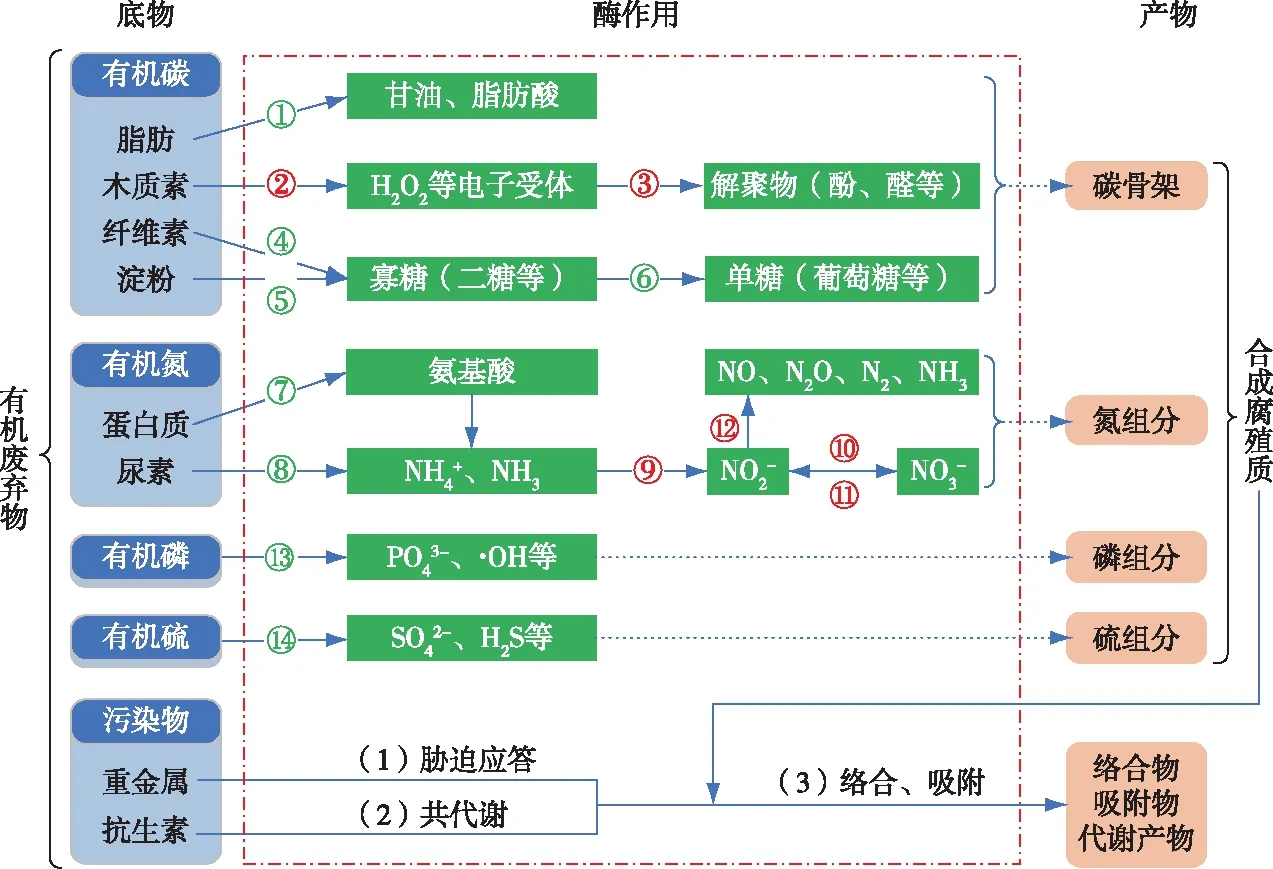
数字编号代表酶种类,详见表1;红、绿色编号分别表示氧化还原酶和水解酶。
1.1 催化降解有机质
蛋白酶分为内肽酶和端肽酶,内肽酶从中间切断大分子蛋白质的多肽链形成相对分子质量较小的朊和胨,端肽酶从多肽的游离羧基末端或游离氨基末端逐一将肽链打断生成氨基酸。氨基酸分解代谢生成α-酮酸和氨,氨代谢形成铵盐和尿素[50]。氮素的转化分为氨化、硝化和反硝化[38-39,51]3个过程。氨化过程指脲酶水解尿素生成氨、二氧化碳。硝化过程(NH3/NH4+→ NO2-→ NO3-)指氨氧化酶(氨单加氧酶和羟胺氧化还原酶)将氨氧化为亚硝酸盐,亚硝酸氧化还原酶进一步将亚硝酸盐氧化为硝酸盐。反硝化过程(NO3-→ NO2-→ NO/N2O/N2/NH3)指硝酸还原酶将硝酸盐还原为亚硝酸盐,亚硝酸还原酶进一步将亚硝酸盐还原为一氧化氮、笑气、氮气和氨气等物质。近期研究报道cytochrome c nitrite reductase (ccNIR)、octahaem nitrite reductase (ONR)和octahaem tetrathionate reductase (OTR)也能催化亚硝酸盐转化为氨气[39],但是尚不清楚有机废弃物降解过程是否存在ccNIR、ONR和OTR,以及ONR和OTR能否作用于微生物的呼吸、亚硝酸盐或羟胺的解毒。
电力企业与人民群众的生活和工作息息相关,人力资源管理工作直接影响企业经济效益和稳定运行。近年,我国电力体制不断改革,很多供电公司开始应用大数据,精准查窃电。前段时间,某国网电力公司与电商公司合作,联合发布“能+”大数据公共服务平台,加快推进电力大数据商业化运营及新型供电服务模式。将大数据应用到电力企业人力资源管理工作中,能够对海量数据信息进行科学存储和分析,使人力资源规划、招聘、培养、绩效考核等一系列工作更加科学、有效。
V表征着生物降解过程中酶作用的强弱(酶活性),它受两个方面因素的影响。一方面,酶具有特异性,每种酶只作用于特定结构的底物、一类化合物或化学键。产酶微生物的筛选[25]、接种[76]及特定酶的提取[77-78]、添加[6,79]可以调整生物酶的种类和浓度。另一方面,S(有机废弃物)饱和可以忽略它的影响,ET和k是关键的控制因子,且k受理化因子约束。控制酶浓度、T、pH等条件[79-82]或添加表面活性剂[83-84]能够提高酶活性。此外,生物酶作为蛋白质易于分解,一旦从活体分离,其酶活性会迅速降低,并对外界环境因素异常敏感。酶的固定化技术克服了这一困难,固定化酶具有可重复利用、稳定性高、易从产物中分离等优势[85]。因而有机废弃物降解过程的酶作用主要有微生物(酶种类和酶浓度)、酶的固定化、理化因子(降解物料的温度、pH和C/N等)和添加剂(激活剂和抑制剂)4个调控途径。
(2)木质素是苯丙烷结构单元组成的复杂有机聚合物,参与其降解过程的酶可分为木质素修饰酶(LMEs)和木质素降解辅酶(LDAs)[22]两大类。LMEs包含酚氧化酶(PO)、木质素过氧化物酶(LiP)、锰过氧化物酶(MnP)、多功能过氧化物酶(VP)等,LDAs包含乙二醛氧化酶(GLOX)、芳醇氧化酶、吡喃糖氧化酶、纤维二糖脱氢酶(CDH)、葡糖氧化酶等。LDAs虽然不能降解木质素,却是木质素降解过程所必需的,其在催化反应中承担传递电子、原子或基团的功能。例如,GLOX能氧化简单的醇或醛生成具有强氧化性的H2O2(电子受体)[22];CDH包含1个黄素和1个血红素基团,能促进MnP的降解作用[45]。基于LDAs作用,木质素的Cα—Cβ键在H2O2、过氧化物酶(LiP、MnP、VP)催化作用下断裂而解聚为芳香族醇、醛、酚类物质,有氧环境中PO还会氧化木质素生成醌类物质[22]。然而,木质素分子结构非常复杂,其降解酶系和催化机制至今尚未完全被认识。
(3)纤维素和淀粉的酶解过程通常分为两步:多糖→寡糖→单糖。第1步,内切和外切葡聚糖酶(属于纤维素酶)作用于纤维素表面,随机攻击β-1,4-糖苷键,截断纤维素大分子形成寡糖(纤维二糖等)和葡萄糖;淀粉酶切割淀粉的α-1,4-糖苷键和α-1,6-糖苷键,水解形成麦芽糖、葡萄糖和糊精[46]。第2步,α-葡萄糖苷酶、β-葡萄糖苷酶分别水解麦芽糖和纤维二糖生成葡萄糖。木质纤维素内存在非常牢固的纤维素-半纤维素-木质素网络结构[47],首先需要木质素酶解聚外层木质素,纤维素酶才能进一步降解内层纤维素。对纤维素酶的探究还在继续,最近BEESON等[48]和TAN等[49]发现纤维素解聚过程存在一种CDH和多糖单加氧酶参与的氧化机制。
1.1.2有机氮
二是知识产权转移转化资本参与度低。一方面知识产权转移转化过程漫长,从研发到工程化、产业化过程,少则几年,多则数十年,投资回报慢,金融投资本身具有趋利性和短视性,因此投资机构不愿花费大量资金在这方面;另一方面政府引导支持范围额度小,社会参与积极性不高,因而资本运作风险大,导致动能不足、效果不佳。大部分高校及科研机构也仅仅是设立知识产权行政性机构,缺乏知识产权转化运营资金投入,这需要国家进行统一筹划,长远布局。
1.1.1有机碳
1.1.3有机磷和有机硫
45例周围神经鞘瘤患者经病理证实确诊,其中良性病变35例,所占比例为77.78%;恶性病变10例,所占比例为22.22%,病变所处部位方面:13例在颈部,所占比例为28.89%;8例在胸部,所占比例为17.78%;10例在腹部,所占比例为22.22%;14例在四肢部位,所占比例为31.11%。
酶的固定化指将游离酶束缚或限制在一定空间内,保留其催化活性,并可回收及重复使用的一类技术。酶的固定化能极大地改善和提升酶的催化活性和稳定性。自20世纪70年代起,酶的固定化技术快速发展,广泛地应用于食品、医疗、能源、环境等领域[90]。传统固定化技术分为吸附、共价结合、包埋和交联4大类;新型固定化技术聚焦固定化载体和固定化方式的创新,如复合新型载体、酶自固定化和多酶共固定化等[85]。这些技术已成功应用到脂肪酶、漆酶、过氧化物酶、葡糖氧化酶、淀粉酶、纤维素酶等游离酶的固定化,其酶活性提高8~54倍,酶回收率高达50%以上[85,91]。部分生物酶活性是与细胞能量代谢相联系的,只有在活体中才能发挥作用,如硝酸还原酶、亚硝酸还原酶等。SACHDEVA等[92]的研究突破了这一限制,该团队利用银纳米颗粒作为载体固定化硝酸还原酶,准确地测定了土壤硝酸盐含量。经过几十年的发展,一些成熟的酶固定化技术已经商业化。然而,当前酶的固定化技术尚缺乏普适性,需要根据酶的特性以及应用需求来选择合适的固定化方法,加强对酶自固定化、多酶共固定化等新型固定化技术的研究是必然趋势。
1.2 促进腐殖质合成
腐殖质合成的机制,目前尚无统一认识。以木质素-蛋白质聚合学说、生物化学合成学说为主流的科学假说认为微生物和酶驱动着有机质分解为酚类、醌类、脂类等化合物,这些化合物再与氨基酸、氨和蛋白质发生聚合反应,形成腐殖质[55-56]。然而,现阶段的实验研究[57-59]仅能表明:腐殖质主要由碳、氢、氧、氮、磷、硫等元素组成,有机碳解聚物构建了腐殖质的基础碳骨架,氮、磷、硫组分为合成腐殖质的辅助原料。
酶作用贯穿有机废弃物生物降解的始终,一方面为腐殖质合成提供充足的原材料(矿化过程的中间产物)和能量,另一方面能催化合成腐殖质。例如,三羧酸循环(TCA)是真菌体内普遍存在的代谢途径之一(糖类、脂类、氨基酸的最终代谢通路和代谢联系枢纽),可以为微生物合成腐殖质提供能量。丙酮酸脱氢酶、丙酮酸羧化酶等酶催化丙酮酸氧化脱羧生成乙酰辅酶A,乙酰辅酶A进入TCA后经过系列酶促反应形成柠檬酸、琥珀酸、苹果酸和三磷酸腺苷等物质。丁杰等[57]对餐厨垃圾堆肥的研究认为TCA过程的产物可能参与腐殖质的合成,如乳酸和丁酸。此外,研究[56,60]报道多酚氧化酶(PPO)能催化醌类化合物与氨基酸、肽、胺等物质缩合生成腐殖质单体分子,如胡敏酸。由于腐殖质合成过程极其复杂,对其合成酶系的组分及机制的探究依旧很困难。
1.3 削减污染物毒性
研究显示,有机污染物(包含抗生素)、金属离子(包含重金属离子)均已在畜禽粪便、生活垃圾等有机废弃物中检出[5,61-62]。有机污染物与金属离子主要通过络合反应引发金属离子-有机物复合污染,且金属有机络合物的形成受到有机污染物官能团组成、金属离子类型、有机物金属离子相对含量等因素的影响[63]。例如,四环素类抗生素含有多个N、O官能团,能与Mn2+、Cu2+等重金属离子发生螯合作用形成有色络合物[5]。值得注意的是,生物酶对有机污染物、金属离子毒性均有削减作用。一方面,酶和微生物联系密切,能通过共代谢作用转化难降解有机污染物[64-65],或是基于胁迫应答机制降低金属离子和自由基的毒性[7,66];另一方面,酶能促进腐殖质合成,生成的腐殖质进一步与有机污染物、金属离子发生络合或吸附作用而改变这些污染物的存在形态[67-68]。生物酶对重金属离子、抗生素、非抗生素有机污染物毒性的削减机制如下:
(1)在重金属离子胁迫下,微生物细胞内会发生一系列代谢变化以达到解毒目的,比如释放抗氧化性酶、激素。例如,王敏[69]报道Cu2+胁迫下微生物体内氧自由基迅速增多,菌体会释放大量抗氧化性酶(超氧化物歧化酶等)和抗氧化剂(胡萝卜素等)以解毒(清除氧自由基)。LUCAS等[7]报道木质纤维素降解过程中H2O2与Fe2+发生芬顿反应产生大量·OH(具有高度活性和非特异性),白腐真菌为了不受·OH伤害,会加速分泌生物酶来解毒(酶能催化降解有机质产生大量小分子有机物,而这些小分子有机物具有良好的·OH亲和性)。此外,腐殖质能钝化重金属离子,以削减重金属毒性。酶作用合成的腐殖质含有羧基、羟基、氨基、硫基、硫酸根、磷酸根等官能团,能与重金属离子发生络合作用[68];同时腐殖质(醌、酚等基团)还能通过电子传递机制发生化学吸附(水溶态)或通过静电力发生物理吸附(固相)[67]。
生物酶大多来源于微生物,因而微生物生长的影响因素(营养条件、环境条件)对酶活性也有一定影响。碳(C)和氮(N)是微生物生长的必需元素,底物C/N大小会影响微生物对养分的吸收量,进而影响微生物活性。当C/N小于20时,氮素相对过剩而释放NH3,氮素利用效率降低;当C/N大于35时,微生物缺乏氮素营养而生长受限,其活性会减弱[100]。不同有机废弃物的C/N存在差异[100],秸秆、木材类C/N较高,粪便、污泥类C/N较低,因而需要调节堆肥物料初始C/N(表2[80,87-88,93-97]),比较适合的初始C/N范围应为20~35。此外,微生物生长对环境条件也存在适应性。研究报道,有机废弃物堆肥过程的最适水分范围为50%~60%[100-101],大多数微生物生长的适宜pH范围为4~9,嗜温微生物和嗜热微生物的最佳生长温度范围分别为35~40和55~65 ℃[12]397-433。WEI等[87]研究认为有机废弃物堆肥过程中酶活性、微生物和环境因子(T、pH和总氮)3者之间存在显著相关性,这也是酶作用受微生物和理化因子影响的体现。堆肥条件的有效控制有利于酶活性的提升和维持。
两组患者均无心源性死亡和再发心肌梗死(0例),治疗组和对照组分别有5例、6例患者行靶血管重建。两组患者MACE比较,差异无统计学意义(P>0.05)。
(3)一般情况,生物酶通过其活性中心将普通有机污染物催化分解成结构简单、毒性较低的小分子产物[65]。若遇到难降解有机污染物(多环芳烃、氯代有机物等),可以通过人工外加生长基质来实现共代谢。共代谢降解作用以关键酶(加氧酶、LiP、MnP和PPO等)和微生物为核心[64],其具体过程包括:a. 生长基质促进微生物生长繁殖;b. 生长基质(或非生长基质,即有机污染物)诱导微生物分泌关键酶;c. 生长基质和非生长基质同时与关键酶的活性中心结合,产生竞争性抑制;d. 生长基质被分解产生能量,非生长基质被降解转化;e. 若非生长基质降解产生的中间产物对关键酶有毒性抑制作用,微生物则会启动自我恢复功能。此外,酶作用合成的腐殖质对有机污染物也有一定的吸附作用,包括共价吸附、“空穴”吸附、氢键作用和疏水性吸附等[74]。
2 酶作用的调控机制
酶通过改变反应路线和稳定中间产物等方式减少反应激活能来提高化学反应速率。酶动力学[18]和热力学[75]公式为
V=(kcatETS)/(KM+S),
垃圾生物处理工艺较为关键的一步是将酶液与热水混合后加入反应器,以促进有机物的分解[6]。有机废弃物堆肥过程中,喷洒微生物菌剂(EM菌剂等)能够增加生物酶分泌量,提高发酵速度[88]。无论是接种高效产酶微生物,还是添加特定酶液,其目的都是通过增加生物酶浓度来加快有机废弃物的降解转化。然而,酶分泌速度和酶提取效率受到限制,不能满足当前的生产需求。构建高效基因工程菌或研发酶生化合成技术,有望实现酶的规模化生产[89]。
(1)
k=Aexp [(lnγ-ΔGTS)/RT]。
(2)
式(1)~(2)中,V、kcat、ET、S和KM分别为反应速率、催化常数、总酶浓度、底物浓度和米氏常数;k、A、R、T、γ和ΔGTS分别为反应常数、指前因子、气体常数、热力学温度、透射系数和过渡态激活能。
(1)脂肪酶攻击酯键,将脂肪逐步水解成甘油和脂肪酸等物质。脂肪酶具有水溶性,其活性位点包含天冬氨酸、组氨酸和丝氨酸(解聚酶的催化三联体)[7]3种氨基酸残基,该催化三联体去质子化后生成的氧基负离子(—O-)攻击脂肪的酯键形成羟基末端(甘油)和酰基-酶复合物,随后水分子攻击酰基-酶键形成羧基末端(脂肪酸)和游离酶。
在进行输出以前,还要设置相关的求解控制参数,包括计算时间控制(设为100 ms)、输出频率、沙漏控制类型、沙漏系数以及缺省设置。由LS-DYNA EXPORTE模块输出K文件,打K文件设置内存空间,以及为提高计算速度而适当进行质量缩放[6]的修改。设置K文件中的参数需要具有一定的理论基础,难度比较大,但是正确的设置会大大提高求解效率,缩短仿真时间,将复杂的仿真模型快速求解。
2.1 酶种类和酶浓度
每种微生物主要分泌一种或少数几种降解酶,每种降解酶只能催化特定结构的底物或化学键。通过宏基因组学技术可以识别编码降解酶的功能基因,并筛选具有特异功能基因的微生物配制复合微生物菌剂。例如,XIE等[86]通过添加具有amoA(AOA,编码氨单加氧酶) 基因的氨氧化古菌,显著加快发酵速度,缩短堆肥时间。针对不同类型底物,可以接种高效产酶微生物或添加有效生物酶来加速降解有机质(表1)。例如,若需要降解纤维素和木质素含量较高的有机废弃物(秸秆等),可以接种芽孢杆菌、白腐菌等菌株或添加纤维素酶、木质素酶等酶液。MAKI等[25]报道从类芽孢杆菌(Paenibacillus)和芽孢杆菌(Bacillus)中分离出的纤维素酶具有高热稳定性、低pH敏感性、高酶活性,从产黄纤维单胞菌(Cellulomonasflavigena)和船蛆杆菌(Terendinibacterturnerae)等菌株分离出的新型多功能纤维素酶能够降解多种底物。WEI等[87]研究发现接种微生物菌剂〔包含链霉菌属(Streptomycessp.)、分支杆菌属(Mycobacteriumsp.)、小单孢菌属(Micromonosporasp.)和糖单孢菌属(Saccharomonosporasp.)〕提高了堆肥过程关键酶(纤维素酶、木聚糖酶、MnP、LiP和PO)活性,促进了纤维素、半纤维素和木质素的降解。冯冲凌等[79]研究了添加酶液(LiP和MnP)对堆肥过程微生物群落代谢能力及木质纤维素降解的影响,发现微生物对中间产物类碳源的代谢能力得到改善,有机质的降解效率显著提高。
第三,由于各路管线众多、管道网络复杂,在施工过程中必须要对管线网络进行统筹考虑,并加强对管线的保护和避让,严格按照规范要求做好规范化施工,以确保工程质量。
2.2 酶的固定化
磷和硫是生物体必需的化学元素,它们在微生物代谢过程中至关重要[52]。通常认为,磷酸酶先催化水解大分子有机磷化合物得到磷酸二酯和磷酸单酯,它们进一步被水解形成PO43-和·OH[53]。硫酸酯酶催化水解脂肪族和芳香族有机硫化合物,使硫酸酯键断裂从而形成无机硫酸盐、醇类、H2S等物质[54]。然而,现阶段对这两大酶系的深入研究相对缺乏,今后还需要进一步识别它们的组分和作用机制。
2.3 温度、酸碱度和碳氮比
温度对酶活性的影响是极其显著的:一方面它会改变酶的稳定性,在5~50 ℃范围内,大部分酶活性随温度上升而增加,>60 ℃条件下有些酶活性开始降低直至完全失活;另一方面,温度会改变酶的动力学和热力学特征,包括速率常数(k)、平衡常数(Keq)、吉布斯自由能变(ΔG)等[12]425-438。例如,k的温度敏感性(Q10)可表示为Q10≈10(ΔGTS-lnγ)/RT2。Top范围大致为30~65 ℃,基本趋势为水解酶Top低于氧化还原酶Top,碳转化酶Top低于氮转化酶Top(表2[80,87-88,93-97])。通过控制堆肥温度,MIYATAKE等[81]研究认为牛粪堆肥的微生物比增长率和酶活性显著提升(54 ℃);张相锋等[98]研究认为秸秆堆肥中有机质降解率大幅增加(60 ℃);常会庆等[99]研究认为污泥堆肥中腐殖质含量提高5.52%(25 ℃)。
蛋白质是α-氨基酸的线性聚合物,其侧链基团带有电荷,因而蛋白质构成的酶具有pH依赖性;电子传递过程中,H+浓度会影响化学键的断裂、酶-底物复合物的形成,进而影响酶催化速率和腐殖质合成效率[12]397-398。酶活性随pH增加的变化趋势呈“钟”型曲线或“S”型曲线,酶作用的适宜pH范围为 4~9[12]398-403。如表2[80,87-88,93-97]所示,pHop范围约为5~9,淀粉酶和蛋白酶pHop较低。姜新有等[82]通过添加过磷酸钙和石灰控制堆肥初始pH(6.42~6.83),不仅减少氮素损失,还提高有机质降解率。此外,缓冲液也能调节催化过程pH,以维持酶活性稳定[12]412-416。

表2 生物降解过程的最大酶活性及其对应理化性质
(2)有机废弃物主要含有四环素类、喹诺酮类、磺胺类、大环内酯类、聚醚类抗生素[70]5大类抗生素。生物酶能够破坏抗生素分子的某些脆弱化学键或阻断抗生素与靶点的结合,从而降低抗生素的毒性作用[71-72]。自1940年首次使用抗生素以来,大量生物酶已被确认可以修饰和降解不同类别抗生素,包括β-内酰胺类、氨基糖苷类、酚类和大环内酯类等抗生素[72]。例如,夏湘勤等[73]报道白腐菌(white-rot fungi)、肉毒链霉菌(Streptomyce)分泌的木质素酶能有效降解诺氟沙星、氧氟沙星和环丙沙星等喹诺酮类抗生素,其毒性可降低70%~90%。
2.4 激活剂和抑制剂
添加剂对酶活性的调控途径包括环境条件、底物和酶系特征[12]379-391。常用的酶激活剂是表面活性剂,其作用机制[102]如下:(1)改变底物结构,增强底物与酶的接触和作用;(2)对生物酶构象产生影响,提高酶的活性和稳定性,改善酶的失活问题;(3)促进生物酶在底物上的有效吸附。如表3[83-84,97,102-111]所示,某些金属离子(Fe2+、Cu2+、K+等)在适宜浓度下,能够活化生物酶功能,因而表现出激活作用。另外,由于堆肥中、后期氮素损失严重,原位保氮的需求促使脲酶抑制剂和硝化抑制剂应运而生,它们能有效减少氮流失[103-104]。与此同时,添加剂对生物酶的调节作用还受到激活剂(或抑制剂)浓度、底物结构特性(疏水性、氢键、官能团、可及度等)、水解/氧化还原条件(水分、pH、搅拌等)、酶系特征(种类和浓度等)的影响[102]。例如,猪粪堆肥过程中低浓度Cu2+(起始浓度为255 mg·kg-1,降解过程为26~117 mg·kg-1)刺激脲酶活性,而高浓度Cu2+(起始浓度为554 mg·kg-1,降解过程为106~518 mg·kg-1)抑制脲酶活性[112];鸡粪和菌糠共堆肥过程中N-正丁基硫代磷酰三胺(NBPT)只作用于脲酶,以达到保氮目的[103]。酶激活剂和抑制剂对酶活性、酶需求量、有机物降解率、产品质量的控制效果显著(表3[83-84,97,102-111])。
常州纺院计算机网络技术专业留学生来源众多,并且毫无汉语基础或者汉语基础薄弱,在职业技能课的学习时就会面临语言难关。因此,英语和汉语相结合的双语教学,在职业技能课的学习中更有必要。

表3 参与有机废弃物降解的酶激活剂和抑制剂
3 研究展望
酶降解高效、环保、可控,是有机废弃物资源化利用的有效解决方案之一。虽然目前对酶降解作用过程及其调控机制的认识尚存在诸多限制和挑战,但是随着分子方法和模型方法的发展与应用,将会逐步揭示微生物-酶-有机废弃物系统的复杂机制,并为进一步高效调控酶作用夯实基础。未来研究中,以下几个方面有待于进一步加强:
(1)有机废弃物酶降解的途径与分子机制。目前人们对有机废弃物主要成分的酶降解过程已有初步了解,但对降解酶系的种类、催化途径和分子机制还不完全清楚,比如木质素降解酶系、有机磷和有机硫转化酶系等。腐殖质的合成机制仍是一个黑匣子,木质素降解酶系催化机制的深入研究有望验证木质素-蛋白质聚合学说的正确性。
(2)微生物-酶-有机废弃物系统的新陈代谢。微生物物种、酶种类、功能基因数量和有机废弃物组分繁多,其形成的代谢网络更是错综复杂。研究微生物-酶-有机废弃物系统的代谢途径与分子机制是系统认识生物降解过程、研发酶激活剂、调控酶作用的基础和关键。同位素示踪、基因测序和网络模型等方法有助于对该新陈代谢机制的识别。
1.2.2 样品近红外光谱扫描及数据预处理 采用Nicolet Antaris II型FT-NIR光谱仪对407个样本进行光谱采集,光谱扫描范围为1 000~2 500 nm,扫描次数64次,分辨率为0.5 nm。3次重复,取平均值作为该样品的最终光谱数据。对烟叶近红外光谱数据进行分析和处理时需对其进行适当的数学操作预处理,最大程度地去除冗余信息,以降低或消除非目标因素对光谱信息的影响[17],从而更利于从复杂的光谱中提取有效信息,在一定程度上提高校正模型的稳健性。在此,通过使用多元散射校正和二阶导数的方法分别对光谱数据进行预处理操作。
(3)激活剂对酶作用的调控。深入研究激活剂对酶系组分之间的协同作用关系,激活剂与有机废弃物组分之间的作用力、动力学和热力学特征,以及激活剂对酶-底物耦合关系的影响,有利于掌握各种激活剂的功能特征和使用方法。
其次,“方便”和“便宜”是影响留学生购买决策的两个重要因素。大多数留学生的可支配收入较低,甚至生活费都是他们父母提供的,所以他们可能更关心产品的价格。因此,在保证自身利益的前提下,网上零售商最好提供最大的优惠价格,以促进他们的网上消费。
(4)酶的固定化技术。固定化技术是改善和提升酶活性和稳定性的关键。一方面,当前固定化技术缺乏普适性,需要进一步深入研究以完善技术;另一方面,需要与企业紧密对接,推动酶的固定化技术在实际生产中的应用。
(5)降解酶的规模化生产与应用。通过构建高效基因工程菌来提高降解酶的表达量或者利用生化合成技术来批量生产酶制剂,在生物降解的基础和应用研究方面都具有重要意义和价值。此外,还需要减少酶需求量和提高酶解效率以降低成本,优化工艺流程和削减污染物毒性以保护环境。
