罗氏沼虾第二步足的相对生长式型及其生态学意义
2020-07-27丁福江
杨 明,丁福江
(1.上海市水产研究所,上海 200433;2.上海市水产技术推广站,上海 200433;3.上海申漕特种水产开发公司,上海 201516)
甲壳动物在其生活史中需要经历多次蜕皮,且不同性别之间及性成熟前后的形态和生长速度存在差异,使得甲壳动物成为研究相对生长的极好材料,一直备受生物学家的关注[1-3]。目前对甲壳纲中虾、蟹等十足目种类的相对生长开展了较多研究[4-6]。但多数集中于雌、雄个体身体大小差异方面,而关于身体的特化器官——螯足的相对生长研究相对匮乏[7-9]。
罗氏沼虾(Macrobrachiumrosenbergii)原产于印度洋-西太平洋的热带、亚热带水域,是我国重要的淡水虾类养殖品种之一。性成熟的罗氏沼虾存在着明显的性二型现象:雄虾不仅在形态规格上显著大于雌虾,而且雄虾依照第二步足长度与身体的比例、第二步足的颜色以及第二步足是否有棘等外部形态特征划分为蓝螯型(blue-clawed males,BC)、橙螯型(orange-clawed males,OC)和小型雄虾(small males,SM)[10-11]。BC、OC和SM雄虾不仅在形态上存在差异,在社群结构中也占据着不同的序列等级[12]。现有的研究指出罗氏沼虾不同形态型雄虾在战斗行为[13]、隐蔽物的竞争[14]、交配和繁殖策略[15-16]等方面存在着差异。第二步足作为罗氏沼虾的附属身体器官,在以上行为中发挥着重要的作用。但目前尚未见对罗氏沼虾第二步足生长式型进行相关报道。因此,本研究基于池塘养殖条件下的罗氏沼虾雌、雄虾第二步足的生长资料,对其第二步足的生长式型和生态学意义进行了研究,为深入开展罗氏沼虾的行为生态学及生产管理提供参考和帮助。
1 材料与方法
1.1 实验材料
实验用罗氏沼虾苗种系购入商业罗氏沼虾苗种繁育场,在室内水泥池进行暂养,期间投喂罗氏沼虾专用配合饲料。养殖水源取自当地河水。养殖实验塘为土池,呈“L”型,面积4 669 m2,配备0.75 kW的叶轮式增氧机2台。
1.2 养殖管理
罗氏沼虾仔虾室内暂养后,转入室外土池开展养殖实验,放苗密度27.85尾/m2。于养殖池中部和四角分别设定饵料台,观察沼虾摄食和生长状况,及时调整投喂量和商品饲料规格。养殖前期逐步添加水至1.2 m,中后期根据水质情况加、换水,每次换水量不超过30%,根据DO测定结果调整增氧机开、关时间,后期延长增氧机开机时间,养殖期间确保DO保持在4 mg/L以上。
1.3 取样与生物学测定
将罗氏沼虾亲虾产卵时间记为养殖的零日龄,待虾苗室内暂养26 d后放入室外土池开展养殖实验,从61日龄开始采样,以后每隔2周采样1次,每次随机取样100尾以上。分雌、雄对罗氏沼虾第二步足长、全长和体重进行测量,用以分析第二步足的生长特性。第二步足测量标准为平直状态下座节基部至指节前端的距离;全长测定标准为虾体平展状态下尾节末端至额角前端的距离;体重为虾体的湿重。鉴于罗氏沼虾为等螯型(isochelous)沼虾,因此仅对其第二步足右足进行测量。养殖前90 d采样使用底拖网,90 d后使用撒网采样,样品带回实验室立即开展生物学测定。长度测量采用数显游标卡尺(精度0.01 mm),体重测定采用感度为0.01 g的电子天平逐尾称重,称重前用纱布吸干虾样体表水分。
1.4 数据分析与处理
采用Keys公式W=aLb拟合第二步足长与体重之间的关系,经过对数转换后化为线性方程lgW=lga+blgL,式中W代表体重,L代表第二步足长。当b=3时,为等速生长;当b<3时,表示负异速生长模式,即体长生长快于体重生长;当b>3时,表示正异速生长模式,即体重生长快于体长生长。对于b值与3之间是否存在显著性差异,则根据下述公式通过t-检验进行分析[17]:
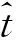
异速生长模型以幂函数方程y=axb计算,以全长为自变量x,依变量y为第二步足长度,a为y轴截距,b为异速生长指数。当b=1时,为等速生长,此时罗氏沼虾第二步足的生长与全长等比例增长;b>1时,为快速生长,此时第二步足的生长要比全长生长快;b<1时为慢速生长,此时第二步足的生长比全长生长慢。异速生长模型中若含有不同生长阶段,则以拐点分开,以决定系数R2最大值与残差平方和最小值作为曲线拟合标准,求得拐点值。采用t-检验方法检验b观测值与1的差异,拐点前后及性别之间的差异同样采用t-检验进行分析。
采用SPSS 21统计软件进行非线性回归参数拟合和显著性检验,P<0.05视为差异显著。
2 结果与分析
2.1 第二步足的足长
试验过程中共计采样1 160尾,其中雌虾604尾,雄虾556尾。经统计分析,雌、雄虾的第二步足长分别为13.69~101.37 mm(56.21±18.66)mm、17.91~178.51mm(71.22±30.19)mm。雌、雄群体的第二步足生长随日龄的变化见图1。可以看出整个养殖周期内,雌、雄群体第二步足呈增长趋势,61~89日龄,雄虾第二步足长度略大于雌虾,两者之间差异不显著,103日龄开始,雄虾第二步足长度均显著大于雌虾,且103日龄后两群体之间差异逐渐增大,雄虾第二步足生长速度快于雌性群体,至养殖末期的187日龄,雄虾第二步足长度是雌虾的1.49倍,呈现明显的性二型现象。
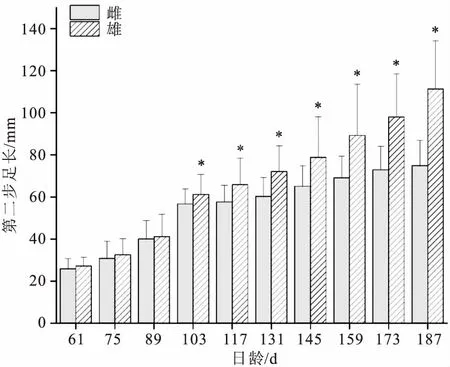
图1 不同日龄罗氏沼虾雌、雄虾第二步足生长统计结果Fig.1 Statistical results of the second cheliped length in female and male M.rosenbergii on different culture days“*”代表雌雄两组间第二步足长存在显著差异(P<0.05)
2.2 第二步足-体重的相对生长
图2为雌、雄群体第二步足-体重的对数回归关系。依图2可知,雌、雄虾第二步足与体重的对数回归呈显著的线性关系,相关系数均在0.94以上,说明第二步足长度生长与体重生长相关程度较高。雄虾的b值小于雌虾b值,表明雄虾的第二步足-体重异速生长水平大于雌虾。
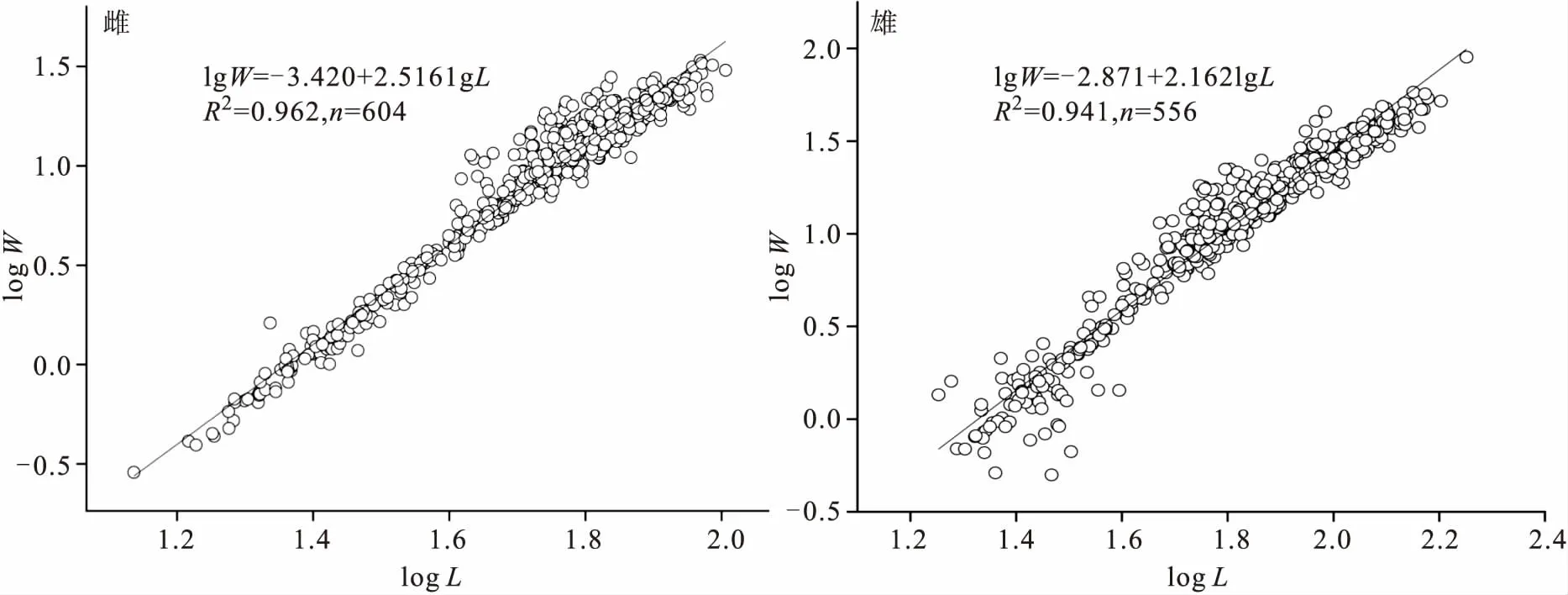
图2 罗氏沼虾雌、雄虾第二步足长与体重的对数回归关系Fig.2 Regression relationship between the second cheliped length and body weight after data log-transformed for female and male M.rosenbergii
对罗氏沼虾雌、雄群体第二步足长-体重的对数拟合关系进行方差分析,检验结果列于表1。由
表1可知,罗氏沼虾雌、雄虾lgW-lgL关系均达到显著水平。为了进一步比较雌、雄群体lgW-lgL的异速生长水平,对回归方程的b值进行t-检验,结果列于表2。从表2可以看出:无论雌虾还是雄虾,其值均大于t标准值(α=0.05),说明第二步足与体重对数拟合方程的b值均显著小于3,雌、雄虾体重的生长为负异速生长式型,即第二步足长度生长快于体重生长。此外,对雌、雄群体的b值通过t-检验进行相互比较,结果显示t=3.58>t0.05(1 156)=1.96,说明雌、雄群体的回归b值不相等,即雄虾的异速生长水平显著大于雌虾。

表1 罗氏沼虾雌、雄虾第二步足长-体重对数回归关系的方差分析Tab.1 Analysis of variance in the relationship between the second cheliped length and body weight after data log-transformed for r female and male M.rosenbergii
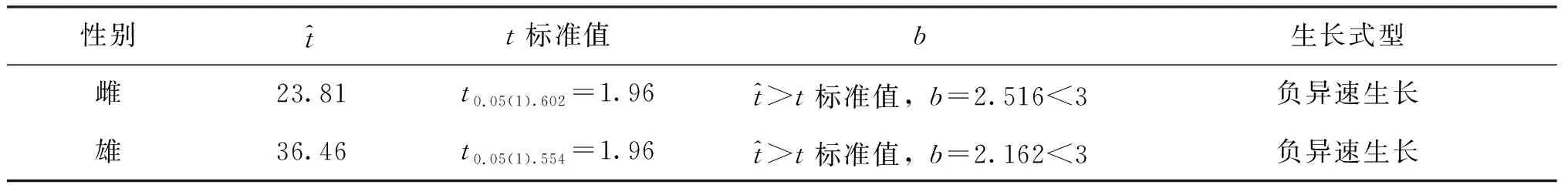
表2 罗氏沼虾第二步足长-体重对数回归关系中b值的t-检验结果Tab.2 The results of t-test of b value in the relationship between the second cheliped length and body weight after data log-transformed for M.rosenbergii
2.3 第二步足-全长的相对生长
由图3可以看出,罗氏沼虾雌、雄群体的第二步足与全长出现了明显的异速生长:雌虾在全长83.10 mm、89日龄出现生长拐点,拐点前第二步足长的异速生长指数b1为1.210,表现为正异速生长(t=14.06>t0.025(137)=1.96,P<0.05);拐点后b2为1.405,表现为正异速生长(t=12.19>t0.025(463)=1.96,P<0.05),且拐点后的异速生长水平显著大于拐点前(t=4.26>t0.05(600)=1.96)。雄虾在全长80.79mm、75日龄出现生长拐点,拐点前第二步足长的异速生长指数b1为1.241,表现为正异速生长(t=12.73>t0.025(103)=1.96,P<0.05);拐点后b2为1.787,表现为正异速生长(t=21.79>t0.025(449)=1.96,P<0.05),且拐点后的异速生长水平显著大于拐点前(t=7.56>t0.05(552)=1.96)。
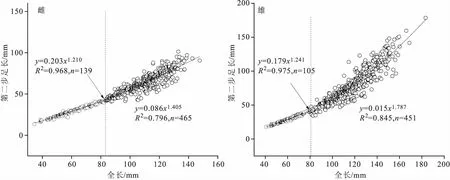
图3 罗氏沼虾雌、雄虾第二步足与全长的异速生长关系Fig.3 Allometric growth of the second cheliped length versus total length of female and male M.rosenbergii
此外,对雌、雄虾拐点前的异速生长指数b1值进行相互比较,显示雌、雄群体拐点前的异速生长水平相等,差异不显著(t=1.27
3 讨论
3.1 罗氏沼虾第二步足的相对生长
步足在种间和种内对有限资源的竞争以及获取和保留隐蔽居所方面起着重要的作用,是甲壳类动物中生长最快的身体器官,是研究相对生长的最佳样本[18]。da Nóbrega等[20]对巴西河口平原的亚马逊沼虾(Macrobrachiumamazonicum)建立了体重-头胸甲长的异速生长方程,结果显示雌、雄群体之间的生长指数b值存在显著差异(雄性>雌性),表明雌虾和雄虾之间可能存在着性二型。本研究结果表明雌、雄群体的体重-第二步足拟合方程的b值均显著小于3,说明体重呈现为显著的负异速生长,即第二步足生长快于体重生长,且雄性的异速生长水平显著高于雌性。
Mantelatto等[19]对自然种群状态下的巴西沼虾(Macrobrachiumbrasiliense)构建了掌节长-头胸甲长及掌节高-头胸甲长的相对生长方程,结果显示掌节长无论是在雌性还是雄性群体种均表现为正异速生长(b>1),而掌节高在雄性群体表现为正异速生长(b>1),在雌性群体中则表现为等速生长(b=1)。本实验中,无论雌虾还是雄虾,其第二步足-全长均呈现出异速生长的特征,且整个阶段的b值均大于1,可知罗氏沼虾雌、雄虾第二步足在拐点前后均为正异速生长,且拐点之后的异速生长水平均比拐点前升高,雄性异速生长水平显著大于雌性。Nagamine等[21]对罗氏沼虾构建了长节-座节的异速生长方程,结果显示在座节12.5 mm处,雄性出现生长拐点。本研究中,雄虾第二步足生长在全长80.79 mm时出现生长拐点,雌虾在全长83.10 mm处出现生长拐点。在61~89日龄范围内,雌、雄虾第二步足长度差异不显著,103日龄开始,雄虾第二步足长度显著大于雌虾,且103日龄后两群体之间差异逐渐增大,雄虾第二步足生长速度快于雌性群体,至养殖末期的187日龄(所采雌、雄虾均已达性成熟),雄虾第二步足长度是雌虾的1.49倍,呈现明显的性二型现象。
3.2 罗氏沼虾第二步足的生态学意义
BC、OC和SM雄虾在社群结构中占据着不同的序列等级[12]:BC雄虾占据社群序列等级的顶端,相对OC和SM均具有强势地位,OC雄虾次之,SM则居于社群序列等级的末端,而且BC和OC雄虾依据身体规格存在着大、中、小三种类型[22],这种情况使得其身体规格和武器大小(第二步足)存在着矛盾的不对称性:BC雄虾尽管体重比OC雄虾少45%和25%,但第二步足长度却比OC雄虾多出10%和20%,则BC雄虾总能在与OC雄虾的战斗中取得胜利;即使BC雄虾的体重比OC少93%,但只要BC与OC的第二步足长度接近,那么BC在与OC的战斗中也有50%的胜率[23]。由此可以看出,BC雄虾获取战斗胜利主要依赖武器(第二步足)大小而不是身体规格。对于一些甲壳动物而言,相较于身体规格,螯足的大小更能体现出优先权的获取能力[24]。对蟹类的行为生态学研究清楚地表明:当身体规格和武器尺寸影响到资源获取时,武器尺寸相对于体型来说,是决定战斗胜负的更重要因素[25]。
对于甲壳动物来讲,缺失螯足会对正常的摄食、生长、繁殖、种内攻击和竞争能力、逃避捕食者等诸多方面产生不利影响[26]。在对罗氏沼虾繁殖行为的研究中,雄性罗氏沼虾通过恐吓(第二步足高扬)或者直接与入侵者发生战斗,均可有效地驱赶竞争对象,并在战斗中取得胜利,从而实现对更多雌虾的圈禁和保护[27]。罗氏沼虾雄性个体的形态差异导致其生殖策略有所不同,即个体较大的BC个体拥有最高的序列等级,对隐蔽场所的占有和异性的交配也占有优势。但以上这些过程都是在身体完整的情况下得以进行的。对于缺少第二步足的雄性罗氏沼虾,势必对其完成交配过程造成一定程度的影响。因此,拥有完整强大的第二步足,可以确保雄性罗氏沼虾获得更高的生殖成功率,从而可将自身的基因传递下去。
