饲喂动物油脂对鸭FMO3基因不同基因型表达水平的影响
2020-07-21张莘王统苗刘本帅江勇白皓张扬王志秀陈国宏常国斌
张莘,王统苗,刘本帅,江勇,白皓,张扬,王志秀,陈国宏,常国斌
(1.扬州大学动物科学与技术学院,江苏 扬州 225009;2.扬州大学农业科技发展研究院,教育部农业与农产品安全国际合作联合实验室,江苏 扬州 225009)
FMO3基因属于黄素单加氧酶家族(FMOs),FMOs是酶的多态性家族,存在于光滑的内质网中,其催化软亲核杂原子物质氧化成它们各自的氧化物[1-3].FMO3是FMOs在成人肝脏中的主要存在形式,对叔胺如三甲胺(TMA)有独特的底物偏好,且机体内TMA氧化为无气味的三甲胺N-氧化物(TMAO)的代谢过程主要由FMO3调节,在肝脏和肾脏中进行[4-7].有研究表明TMAO以及TMAO前体如胆碱会在慢性肾病患者中累积且是尿毒症的毒素[8-9],且TMAO被确定为肾病患者的心血管危险因素[10].FMO3的调节直接影响全身TMAO水平,血小板反应性和体内血栓形成速率,与动脉粥样硬化有关[11].一些动物产品也存在由于来源中含有过量的TMA,因此呈“鱼腥味”“腥味”等[12],表明牛奶中的鱼腥味是由牛FMO3外显子6中的无义突变(R238X)导致该密码子变为终止密码子引起FMO3活性缺失;许多家禽物种中也发现了FMO3突变与蛋黄中TMA水平升高和畜产品的鱼腥味之间的有显著关联,如鸡[13]、鹌鹑[14]、鹅[15].有研究报道TMA与食物的新鲜程度有紧密联系,是产生鱼腥臭味的主要物质[14].TMA、TMAO和胆碱由膳食磷脂酰胆碱产生,磷脂酰胆碱在高脂肪食物中含量丰富[16-17].目前,国内外关于鸭FMO3基因的相关报道较少,而FMO3基因在鸭上的研究对其抗病育种和畜产品鱼腥味有重要意义,同时鸭肉又是我国的重要畜产品之一.目前世界上饲养的肉鸭品种以北京鸭及在其基础上选育的品种(‘樱桃谷北京鸭’)为主.鸭具有生长快、瘦肉率高、净肉率高和饲料转化率高,以及抗病力强等优点.本研究通过饲喂鸭2种动物油脂含量不同的饲料,对鸭的FMO3基因第二、六、七外显子的多态性进行检测,qPCR法检测肝脏、肾脏中FMO3基因的表达,对鸭FMO3基因多态和动物油脂与FMO3基因在肝、肾中的表达进行关联分析研究,旨在为FMO3基因的表达调控提供理论依据.
1 材料与方法
1.1 试验材料
1.1.1 试验动物 79只0日龄的‘樱桃谷鸭’购自益客集团.
1.1.2 试验试剂 无油脂饲料、动物油脂均购自益客集团;Trizol和反转录PCR试剂盒Prime Script TMRT reagent Kit购自天根生化科技有限公司;qPCR试剂盒购自诺维赞公司.
1.2 动物饲养
本试验的鸭在益客集团采用网上平养的方式进行饲养,将79只0日龄鸭平均分为A、B 2组,每组雌雄各半,前2周均使用不含动物油脂的饲料进行育雏,第3周开始,A组继续饲喂不含动物油脂的饲料,B组饲喂含10%的动物油脂的饲料,到上市日龄(42日龄)时屠宰.
1.3 DNA提取
在鸭28日龄时,用苯酚和氯仿法提取基因组DNA,选择完整性好(无拖尾现象),D260/D280介于1.8~2.0之间的DNA样品稀释至50 ng/μL,-20 ℃保存备用.
1.4 基因分型
Wang等[18]的研究表示FMO3的错义突变发生在第二外显子、第六外显子和第七外显子.所以本研究根据NCBI上北京鸭FMO3的基因序列设计引物(表1),引物合成由北京擎科生物技术有限责任公司进行.运用PCR技术扩增FMO3第二、第六和第七外显子,将PCR产物送测序公司进行测序.采用MEGA6软件对不同个体测序片段进行序列比对,并结合MEGA6软件对差异碱基测序图进行比较分析,去除可能存在的误判,确定突变位点基因型,计算基因型频率和基因频率.

表1 鸭FMO3基因错义突变检测引物
1.5 鸭FMO3基因表达量的测定
利用实时定量PCR法检测肝脏、肾脏中FMO3基因的表达.鸭42日龄时采肝脏、肾脏置于RNA保护液中,-80℃保存.AA和AG基因型,每组五雄五雌,GG基因型每组4只,做肝脏、肾脏中的相对定量分析.细胞总RNA的提取采用Trizol法按说明进行,将D260/D280介于1.9~2.0之间的RNA浓度稀释至500 ng/μL,-80 ℃保存备用.反转录PCR采用试剂盒按说明进行,实时定量PCR采用试剂盒按说明进行.引物序列见表2.

表2 鸭FMO3基因及内参GADPH mRNA荧光实时定量PCR引物
针对FMO3基因表达及内标基因GAPDH,每次反应中,每个样本重复3次,最后取平均值,每个样本的相对表达量的值等于目的基因的表达量的值除以GAPDH的表达量均值,肝脏组织目的基因的相对表达量采用2-△△Ct方法计算试验原始数据.个体样本之间的表达差异用IBM SPSS Statistics22统计软件进行多重比较.
2 结果与分析
2.1 鸭FMO3错义突变基因分型
用表1中的引物对鸭基因序列进行扩增,经检测PCR产物好,特异性好,符合测序要求(图1).将PCR产物送测序公司测序,检测第二外显子、第六外显子、第七外显子区域SNP如表3所示.在所发现的9个SNPs中除SNP1、SNP3、SNP8和SNP9为错义突变外,其余均为同义突变.SNP1导致其编码的第12位氨基酸发生改变,命名为V12I错义突变,且该突变位于FMO3蛋白结构的FAD结合区.SNP3导致其编码的第252位氨基酸发生改变,命名为S252N错义突变.SNP8导致其编码的第335位氨基酸发生改变,命名为Y335H错义突变.SNP9导致其编码的第336位氨基酸发生改变,命名为S336P错义突变.且Y335H和S336P连锁.
由于在突变纯合子中一共有4只试验动物出现S252N突变,且在雌性个体中只有一个出现,所以暂未对其进行分析.经过对鸭雄性和雌性群体V12I、Y335H、S336P突变基因型进行卡方适合性检测,发现2个群体处于遗传平衡状态(P>0.05).从表4中我们可以看到,V12I突变位点A突变基因的基因频率在雄性和雌性群体中分别为0.65和0.56,均高于同群体中等位基因G的基因频率,2个群体在V12I突变位点的基因频率并不一致.因为Y335和S336P 2个位点连锁,所以Y335H和S336P突变基因型卡方适应性检测结果相同(表5),Y335H和S336P突变位点C突变基因的基因频率在雄性和雌性群体中分别为0.325和0.24,均低于同群体中等位基因T的基因频率,2个群体在Y335H和S336P突变位点的基因频率不一致.
图1 PCR产物琼脂糖凝胶电泳Figure 1 Agarose gel electrophoresis of PCR products

表3 鸭FMO3基因外显子二、外显子六和外显子SNPs检测

表4 V12T突变G/A基因型频率和基因频率
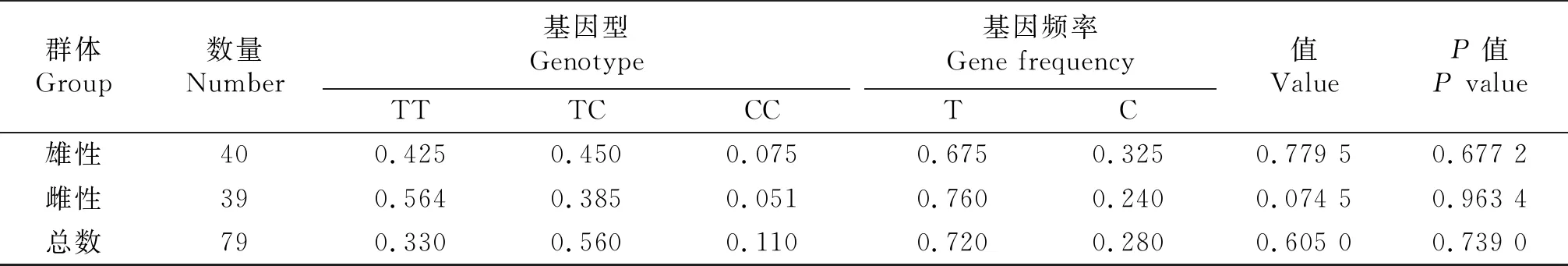
表5 Y335H和S336P突变T/C基因型频率和基因频率
2.2 鸭FMO3 mRNA在肝脏和肾脏中的相对定量分析
本试验以V12I突变位点的基因型,来探究基因型、是否添加10%动物油脂及其交互作用对肝脏、肾脏FMO3mRNA表达水平的影响(表6).基因型间肝脏FMO3mRNA表达量无显著性差异(P=0.181 ) ,AA基因型高于AG和GG基因型.肝脏FMO3mRNA表达水平受动物油脂添加的影响,在添加10%的动物油脂之后,肝脏FMO3mRNA的表达水平较不添加动物油脂的处理组显著降低17%(P<0.05).单因素方差分析结果表明,饲料中添加10%的动物油脂,降低了AA基因型(18%)和AG基因型(2.7%)肝脏FMO3mRNA的表达水平,但差异不显著;显著降低了(28%)GG基因型肝脏FMO3mRNA的表达水平(P<0.05).是否添加10%的动物油脂和基因型对肝脏FMO3mRNA的表达无明显交互作用.肾脏FMO3mRNA的表达没有显著性差异,是否添加10%的动物油脂、基因型以及二者的交互作用对肾脏FMO3mRNA的表达没有显著性影响.
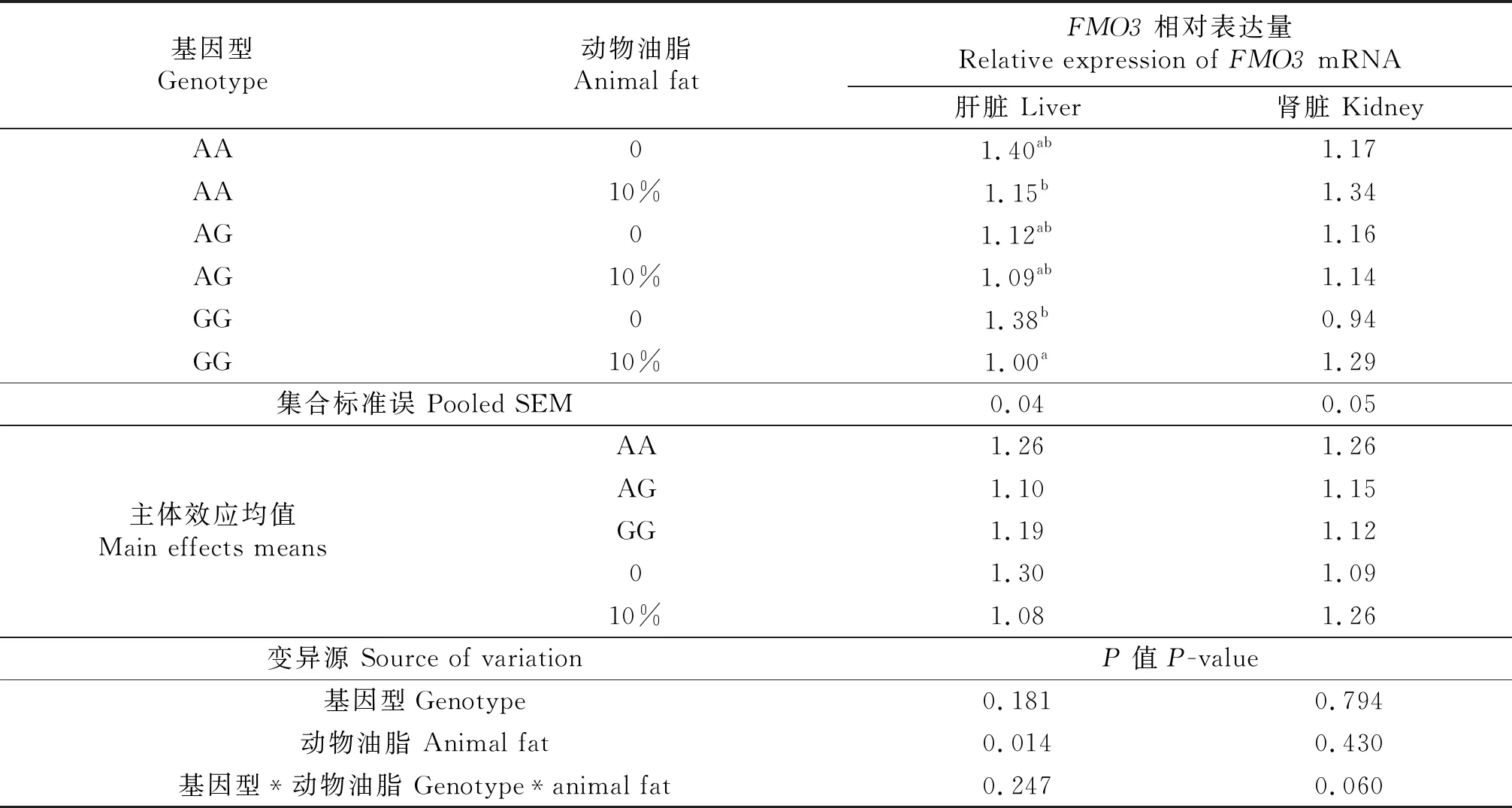
表6 FMO3基因型和动物油脂对鸭肝脏、肾脏FMO3 mRNA表达量的影响
3 讨论
Lawton等[19]推测FMO3蛋白结构包含N终端FAD结合域、NAD(P)H结合区域、一个中央区域和C端接口域.另外,FMO3还包含有FMO特征基序和FATGY基序,其中FATGY基序被认为参与底物识别,是酶与底物的结合区域的关键基序[20].FMO3基因多态性可以影响到很多外源物质的代谢,因FMO3功能降低或丧失会使TMA不能正常代谢排出体外,当过多的三甲胺排出时会使机体产生明显的鱼腥味.这种疾病被称为原发性三甲胺尿(TMAU),这是一种常染色体隐性遗传疾病[1,21].本研究结果表明鸭FMO3基因第二、六、七外显子区域的9个SNPs中有4处为错义突变,这一结果和Wang等[18]一样,.对这些错义突变基因型进行卡方适应性检测,发现2个群体处于遗传平衡状态(P>0.05).在这些错义突变中,SNP1导致其编码的第12位氨基酸发生改变,命名为V12I错义突变,且该突变位于FMO3蛋白结构的FAD结合区,在FATGY基序没发现错义突变,这和Honkatukia等[13]在鸡上的研究结果不同,可能由于鸭的FMO3基因比鸡的更保守.由以上结果可以看出V12I错义突变可能与FMO3基因功能有关.然后,本研究对V12I错义突变个体的肝、肾FMO3基因的表达进行测定分析发现GG突变纯合子个体在饲喂添加10%动物油脂时,肝脏中FMO3基因的表达量显著低于饲喂不含动物油脂的处理组(P<0.05),AA、AG 2种基因型在是否饲喂含10%的动物油脂2个处理组中肝脏FMO3基因的表达量均没有显著性差异(P>0.05).而是否添加10%的动物油脂、基因型以及二者的交互作用对肾脏FMO3基因的表达没有显著性影响(P>0.05),这一结果和王晶等[7]在鸡上对于T239S位点在肝、肾中表达的研究结果相似.
本研究表明,饲料中添加动物油脂可抑制有V12I错义突变的鸭FMO3基因在肝脏中的表达,且对GG基因型的影响显著,饲料中添加动物油脂和基因型与鸭FMO3基因在肝脏中的表达调控有关,可为鸭FMO3基因的表达调控研究以及饲粮配方设计提供理论依据.
