草莓Remorin家族的全基因组鉴定及功能初步研究
2020-07-18温陈金薛程柯美玉
温陈金 薛程 柯美玉
摘 要:研究将FvREM2.1基因异源过表达到拟南芥中,结果表明:过表达FvREM2.1基因能明显抑制拟南芥根生长,引起根的向重性失衡,且在pSUC2:GFP背景下过表达FvREM2.1导致GFP在根部卸载受抑制,表明FvREM2.1通过调控胞间连丝的通透性,参与了草莓根的发育。
关键词:草莓Remorin家族;全基因组;鉴定;功能
草莓果实主要由其膨大的花托和种子构成,具有浓郁的特殊香气,被誉为“水果皇后”。对于植物来说,健康且发达的根系是保障其高产、稳产的重要前提。从育种上提高草莓根系的生命活力,是解决上述问题的有效方法之一。解析草莓根系发育的分子机制,可为草莓的分子辅助育种和基因工程改良提供重要的理论与应用基础。
在植物生长发育过程中,同化物的运输效率影响着植物的生长和果实的发育[1]。阐明胞间连丝通透性的调控机制,可进一步揭示草莓根中的同化物运输机制,为草莓分子辅助育种和基因工程改良提供理论依据。Remorin(REM)蛋白定位在胞质膜上,参与调控植物多个生理通路,而成为一个重要的研究对象[2]。
本研究利用生物信息学软件,对FvREM基因家族进行保守结构域分析、序列特征分析和聚类分析等,并结合转录组数据筛选到了在根部特异高表达的FvREM2.1。拟南芥中过量表达FvREM2.1,表现为明显根的伸长受抑制及向重性缺失。解析草莓FvREM基因家族成员的生物学功能,为进一步阐述草莓根系发育的机制研究提供了重要的基因资源,并且对草莓进行分子辅助育种和基因工程改良等均具有重要意义。
1 材料与方法
1.1 试验材料
植物材料和生长条件:所用的植物材料有二倍体草莓品种,为森林草莓(Fragaria vesca Ruegen红果),及pSCU2:GFP背景株系为野生型的拟南芥(Arabidopsis thaliana)。这些草莓和拟南芥植物种植于人工培养室中,室温为恒温22 ℃,光照强度为8 000 lx,光周期为光照16 h、黑暗8 h。
1.2 草莓REM家族成员进化树构建及其在蛋白质位置的分析
草莓REM基因的鉴定,使用来自拟南芥的REM蛋白序列作为查询条件,在PHYTOZOME V12.0网站上进行BLASTp搜索,通过同源比对法鉴定得到了草莓REM基因家族成员。拟南芥AtREM基因文件从TAIR网站(拟南芥信息资源,http://www.arabidopsis.org)下载。为了探讨草莓与拟南芥之间的进化关系,将拟南芥AtREM(16个)和FvREM(11个)进行多重序列比对,利用MEGA X构建了拟南芥与草莓的系统发育树[3]。
FvREM基因的基因结构通过在线网站:Gene Structure Display Server(GSDS)[4]进行呈现。
使用Motif analysis tool Multiple Em for motif Elicitation(MEME:http://meme-suite.org/tools/meme)(Bailey et al.,2009)对FvREM家族成员进行保守motif检测,默认参数为motif位点分布、顺序分布为每个序列出现0次或1次(zoops)。
基因表达分析:不同草莓品种的RNA-Seq数据[5-6]来自以对数(1og2)转化每个REM基因每千碱基每百万分(RPKM)值,用R绘制表达量热图。
1.3 DNA、RNA的提取及cDNA的合成
森林草莓Ruegen总DNA的提取,采用改良CTAB(Lade et al.,2014)的方法。森林草莓Ruegen总 RNA 提取用(DP441)RNAprep Pure多糖多酚植物总RNA提取试剂盒进行。RNA 样品核酸浓度测定和质量检测用NanoDrop 2000超微量分光光度计(Thermo Fisher Scientific 公司)进行。总RNA 的cDNA反转录试剂盒为TransScript One-Step RT-PCR SuperMix(北京全式金生物技术有限公司)。
1.4 CDS克隆和表达载体构建
在PHYTOZOME V12.1数据库中获得森林草莓FvREM2.1基因序列,设计擴增引物 FvREM2.1-F 和 FvREM2.1-R(表1),进行 PCR 反应。利用PrimerSTAR GXL DNA聚合酶(TaKaRa, Japan)以Ruegen的gDNA为模板,扩增出FvREM2.1基因片段。扩增体系为50 μL:2× PrimeSTAR 酶1 μL;dNTP Mixture(2.5 mM each)4 μL;5× PrimeSTAR GXL Buffer 10 μL;Steril-ized ddH2O 32 μL;引物 FvREM2.1-F 和FvREM2.1-R各1 μL;模板1 μL。反应条件为:98 ℃预变性 5 min;98 ℃变性30 s,58 ℃退火90 s,72 ℃延伸90 s,共32个循环;72 ℃延伸10 min。将回收的片段亚克隆连接到pDONR221中,重组质粒经PCR验证后,挑取测序正确的质粒。利用Gateway技术将FvREM2.1片段与pDONR221重组载体插入pMDC7B中。用扩增引物(表1)鉴定最终载体的阳性克隆,得到FvREM2.1基因目的条带,表明已成功转入,最终诱导过表达载体。
1.5 农杆菌介导转化拟南芥
将克隆好的XVE:FvREM2.1载体通过冻融法转入农杆菌株GV3101,用GRS(庆大霉素、利福平霉素、壮观霉素)抗性筛选出目的菌株,挑取菌点加在10 mL带GRS抗性的YEB液体中,在28 ℃ 200 rpm摇床培养16 h后,用电泳凝胶鉴定菌液中的目的条带。选取正确的菌,加到50 mL带GRS抗性的YEB液体中,在28 ℃ 200 rpm摇床培养16 h后,对拟南芥花序进行浸花法侵染。将获得的 T0代拟南芥种子消毒后播种在含有HYG(潮霉素)抗性的 1/2 MS培养基中培养,初步筛选出绿色抗性苗并移入土中。提取 T1代抗性苗的DNA和RNA,进行PCR和qRT检测(检测引物FvREM2.1-F和FvREM2.1-R,表1)转基因株系。
1.6 拟南芥表型分析
转基因株系与背景株系种植在1/2 MS培养基上和在1/2 MS+5 μM ES(雌二醇)培养基上,观察植物生长状况。用激光共聚焦显微镜观察在ES诱导下的植株根尖pSUC2:GFP的分布情况。激发共聚焦的激发参数和检测参数设置分别为GFP:488 nm,505~550 nm。
2 结果与分析
2.1 草莓REM基因家族的全基因组成员鉴定与进化树分析
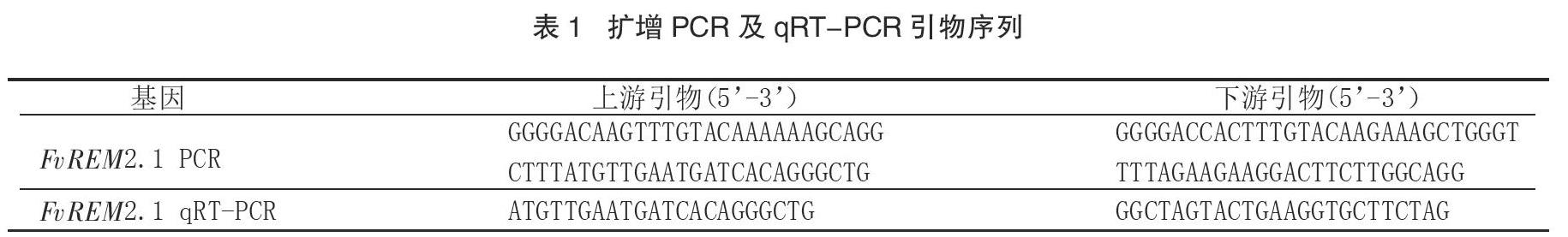
根据16个拟南芥REM 基因(AtREM)与11个草莓 REM 基因(FvREM)的氨基酸序列,利用 MEGA X构建系统发育进化树(图1)。系统发育树显示,拟南芥和草莓的REM成员被分为4组,系统发育学分析探讨了它们之间的进化关系。Group4中成员最多,共有12个成员,其中AtREM和FvREM各有6个;Group3仅有拟南芥成员:1个AtREM3.1和2个AtREM6s成员,不包含草莓基因;FvREM2.1与AtREM4.1,FvREM4.2组成了Group2,该簇的拟南芥成员在功能上已有报道,参与抗病毒防御;Group1主要包括了FvREM1s及AtREM1s全部成员和AtREM3.2。
2.2 基因结构和蛋白保守结构域分析
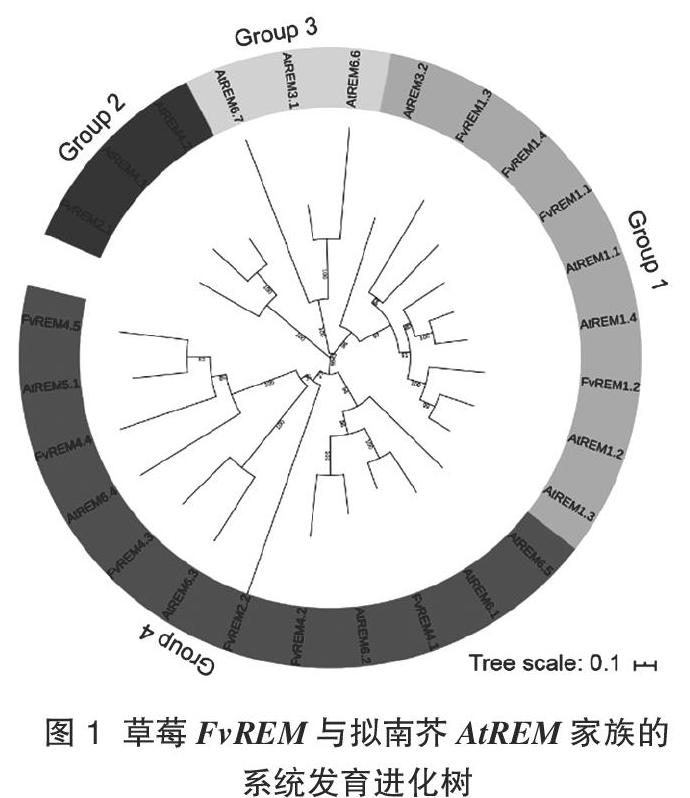
为分析草莓REM的基因结构,根据获得的基因组及CDS序列,绘制成显示外显子-内含子基因结构图(图2)。结果表明,REM家族基因都含有内含子,FvREM2.1只有1个内含子,其他成员最多可达6个内含子。该家族成员的特征是含有一个高度保守的
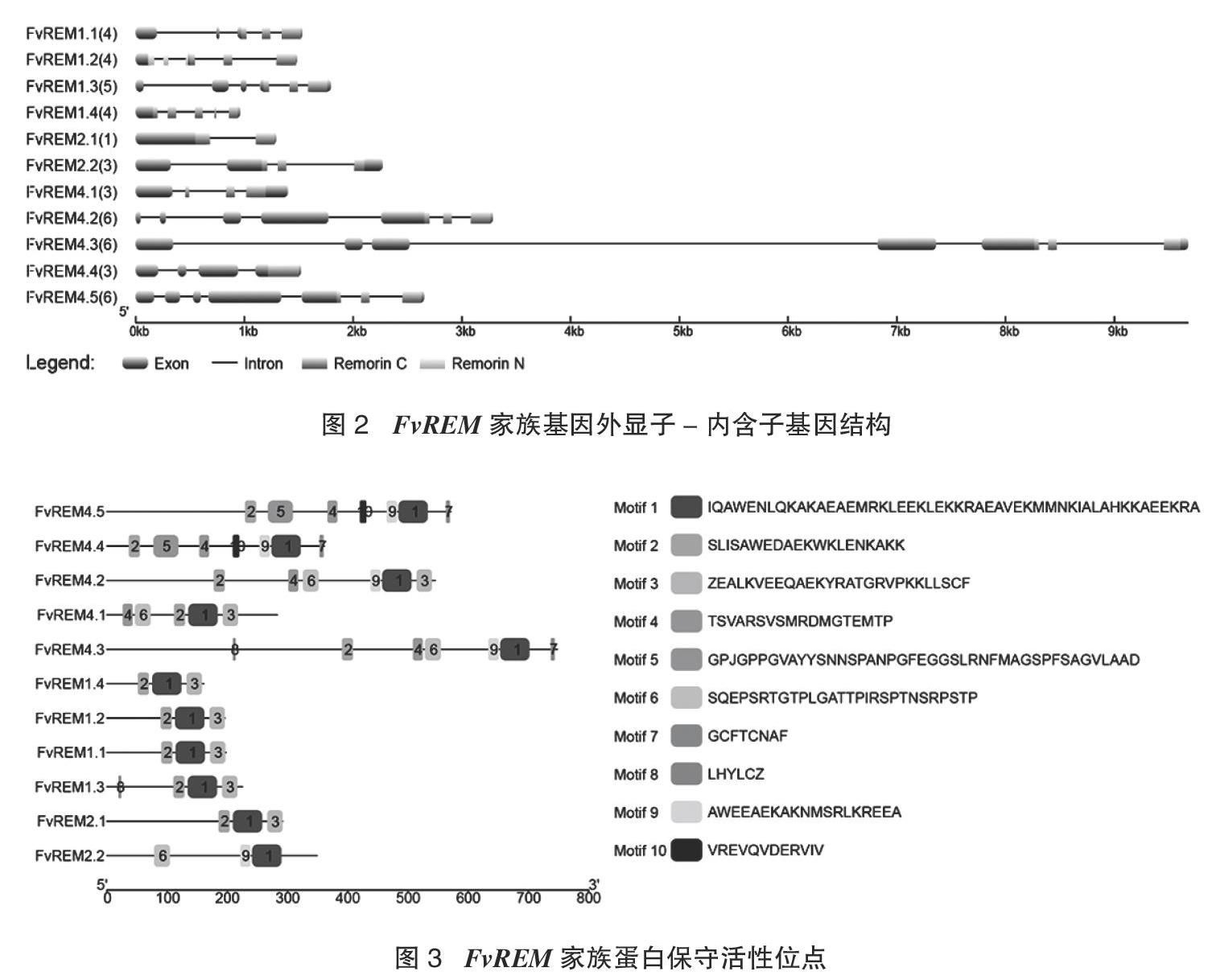
C端区域,该区域包含一个螺旋结构域和一组保守的正电荷和脂肪族氨基酸残基,与质膜(PM)的低聚化和定位相关[7]。相比之下,在不同的FvREM中,N-末端区域是高度可变的或缺失的。其中,仅有FvREM1.2的序列含有N端,其他序列则主要含C端。FvREM基因的開放阅读框长度在965~9 681碱基,编码的多肽链长度在160~747氨基酸。经MEME在线软件的分析,得到10个保守motif,其中,FvREM家族基因都含有保守的motif 1,除了基因FvREM2.2外,其他家族基因都含有motif 2。FvREM1所有的成员,FvREM2.1及FvREM4.1都含有motif 2-motif 1-motif 3紧密排列的结构(图3),推测它们在功能上较相近。
2.3 FvREM不同时空的表达分析
FvREM家族基因在转录水平上有一定的组织特异性和时序性。其中FvREM4.2、FvREM4.3及FvREM4.5在SAM(茎尖分生组织)、FM(花分生组织)、REM(花托分生组织)有相对较高的表达。FvREM4.5在各不同时空中表达水平较稳定。FvREM1.2整体上有相对较高的表达量;相较之下,FvREM1.3整体上表现为较低水平表达量,仅在第二时期瘦果种皮和叶片中较高表达;FvREM1.4则是在特定时期的花药和花柱中有较高表达;FvREM2.1整体表达量不突出,但在根中有特异高表达。由此推测,不同FvREM家族基因可能参与调控不同的生物功能,而FvREM2.1在根中特异高表达,推测其可能在根中发挥着重要功能(图4)。
2.4 过表达FvREM2.1影响拟南芥pSUC2:GFP根系发育
侵染后代经筛选获19个独立株系。在1/2 MS+
5 μM ES培养基上诱导T2代XVE:FvREM2.1株系过表达,T2代植株表现为明显抑制根的生长,与对照比较为显著差异;并伴有向重性失衡(图5)。
在激光共聚焦显微镜下观测根部pSUC2:GFP的表达,其中过表达株系T2-5与T2-6株系诱导后表型最为明显,pSUC2:GFP荧光被限制在中柱中,而对照中柱的荧光会扩散到周围皮层中(图5)。在1/2 MS培养基上生长,无ES诱导的对照与转基因株系无显著差异。
3 讨论
本研究根据已报道的拟南芥REM基因家族信息,从PHYTOZOME上获得FvREM家族基因序列,结合Kang和Darwish等文章中FvREM的转录组数据,在对其进行生物信息学分析后,发现该家族有较高的保守性,但不同的成员在不同的时空分布上表达量存在差异,推测该家族基因含有不同的功能。其中,FvREM2.1在根中特异表达,作为本研究的重点研究对象。
本研究中FvREM2.1在拟南芥中过表达,表现出明显根的伸长受抑制及向重性失衡;过表达FvREM2.1模拟了拟南芥受到生物胁迫从而响应防御系统的过程,使得拟南芥根系荧光信号pSUC2:GFP被限制在中柱中,无法卸载到周围皮层细胞,即胞间连丝的通透性受阻,表明FvREM2.1可能参与防御系统中胞间连丝的调控,从而影响了根系的发育,推测其在植物抗逆与生长发育的平衡中有一定积极作用。
草莓的生产过程中容易受土传病害、土壤养分失调等问题的影响,阻碍根系的发育。同时,根系又是草莓营养生长的“库”器官,地上部分合成的有机物不断地向根运输,促进良好的根系形态建成,对草莓的产量和品质具有重要意义。本研究发现FvREM2.1调控拟南芥根中胞间连丝的通透性,影响了根系发育,揭示了FvREM2.1在草莓根系发育中参与胞间连丝的调控作用,促进了对草莓根系防御系统调控机制的了解,为改善草莓根际微生物域环境提供新的解决思路,对于培育具有良好根系结构的草莓品种具有一定的借鉴意义。
参考文献:
[ 1 ] LEE J Y,FRANK M. Plasmodesmata in phloem:different gateways for different cargoes [J]. Curr Opin Plant Biol,2018, 43(6):119-124.
[ 2 ] RAFFAELE S,BAYER E,MONGRAND S. Upregulation of the plant protein remorin correlates with dehiscence and cell maturation [J]. Plant Signaling & Behavior,2009,4(10): 915-919.
[ 3 ] LIANG P,STRATIL T F,POPP C,et al. Symbiotic root infections in Medicago truncatula require remorin-mediated receptor stabilization in membrane nanodomains[J]. Proc Natl Acad Sci U S A,2018,115(20):5289-5294.
[ 4 ] HU B,JIN J,GUO A-Y,et al. GSDS 2.0:an upgraded gene feature visualization server Bioinformatics[J].Genome analysis, 2015,31(8):1296-1297.
[ 5 ] KANG C,DARWISH O,GERETZ A,et al.Genome-scale transcriptomic insights into early-stage fruit development in woodland strawberry Fragaria vesca[J].Plant Cell,2013, 25(6):1960-1978.
[ 6 ] DARWISH O,SHAHAN R,LIU Z,et al. Re-annotation of the woodland strawberry (Fragaria vesca)genome[J].BMC Genomics,2015,16(29).
[ 7 ] RAFFAELE S,MONGRAND S,GAMAS P,et al.Genome-wide annotation of remorins,a plant-specific protein family: evolutionary and functional perspectives[J].Plant Physiol, 2007,145(3):593-600.
