不同海拔梯度灌丛草甸群落多样性的分布特征
——以五台山亚高山-高山带南坡为例
2020-07-16张晓龙邓清月秦浩史利江苏迎庆张殷波牛俊杰
张晓龙,邓清月,秦浩,史利江,苏迎庆,张殷波,牛俊杰*
1.山西财经大学资源环境学院,山西 太原 030006;2.太原师范学院汾河流域科学发展研究中心,山西 太原 030619;3.山西财经大学统计学院,山西 太原 030006
山地生态系统是具有独特生态-水文过程的生态系统,区域内海拔变化引起的水热条件差异,使得各种生物和非生物因子在海拔梯度上也表现出梯度性变化,这为研究植物群落结构和多样性特征对海拔变化的响应提供了理想的场所(Körner,2007;Tang et al.,2012;Fyllas et al.,2017)。在山区,海拔变化是影响植物群落结构和动态的关键因素,直接或者间接的调控着山地植被的群落结构和多样性组成(Liu,2017;张树斌等,2018)。以往对热带、亚热带和其它山区的研究表明,物种丰富度和多样性指数以“中间高度膨胀”格局较为常见,即中海拔山区植物群落具有最高的物种丰富度(唐志尧等,2004;刘哲等,2015;黄甫昭等,2016)。然而,也有研究表明群落多样性随海拔升高可表现为先降低后升高、单调升高、单调下降、“M”型和恒定不变的分布格局(贺金生等,1997;丛明旸等,2017;邓清月等,2019),实际上,植物群落多样性随海拔变化在不同研究区域或海拔梯度下具有不同分布格局(王志恒等,2004)。因此,开展不同海拔梯度下植物群落多样性分布特征研究,探寻山地植被与海拔之间的关系,有助于认识不同地区山地植被对环境变化的适应,也有助于更加深入地理解山地景观异质性的机制(Zhang et al.,2016;Xu et al.,2017)。
灌丛草甸植被作为高寒山地生态系统的重要组成部分,对维持植物群落、生态系统结构和功能稳定以及水源涵养方面起着至关重要的作用(孟凡栋等,2016;谢宗强等,2017;戴黎聪等,2019)。灌丛草甸在五台山地区垂直分布具有典型性和代表性,在海拔2200 m以上各个植被带内均有分布(崔海亭,1983)。五台山具有典型的亚高山-高山生态系统,是响应全球变化和人为干扰的关键性研究地区,也是华北地区生物多样性保护的重点区域之一(江源等,2009)。五台山地区植被的研究历史较早(张金屯,1986),但有关五台山植物群落的研究主要侧重于亚高山草甸、森林植被、冰缘地貌植被生态学特性以及旅游、放牧等人类干扰活动对植被的影响(刘鸿雁等,2003;江源等,2010;吕秀枝等,2010;Niu et al.,2019)。在当前环境条件下,关于亚高山-高山灌丛草甸生物多样性在连续海拔梯度上的宏观格局,尤其是亚高山-高山灌丛草甸群落 α、β多样性的垂直分异特征的研究相对缺乏,尚需验证和探讨。
本研究以五台山亚高山-高山带南坡沿海拔梯度自然分布的灌丛草甸群落为研究对象,探究灌丛草甸群落 α、β多样性沿海拔梯度的变化规律,分析群落结构和多样性与海拔间的关系,旨在为五台山地区灌丛草甸资源的保护和管理提供科学参考。
1 材料与方法
1.1 研究区概况
五台山(38°30′—39°15′N,112°50′—113°50′E)地处太行山脉北端,是华北地区最高山峰,最高海拔为 3058 m(张金屯,1986)。该区属于典型温带大陆季风山地气候,受地形影响,高海拔山区年均温在-5 ℃左右,无霜期 60—70 d,部分海拔地段四季积冰,年均降水量在1000 mm左右,夏季多地形雨;中低海拔山区年均温为 4—6 ℃,年均降水量在500—650 mm(张金屯,1986,江源等,2010)。五台山海拔高差大,植被分布呈明显的垂直地带性,发育了较为完整的暖温带植被垂直带谱,亚高山-高山带南坡以亚高寒-高寒灌丛草甸和草甸为主,代表植物有金露梅(Potentilla fruticosa)、鬼箭锦鸡儿(Caragana jubata)、高山嵩草(Kobresia pygmaea)、披针苔草(Carex lanceolata)、铁杆蒿(Artemisia sacrorum)等,亚高山-高山带北坡以针阔混交林为主,代表植物有华北落叶松(Larix principis-rupprechtii)、白杄(Picea meyeri)、青杄(Picea wilsonii)、油松(Pinus tabulaeformis)等,山顶发育高寒草甸植被(崔海亭,1983;张金屯,1986)。土壤类型主要有山地褐土、棕壤、亚高山草甸土等。
1.2 野外调查与取样
2019年7—8月沿五台山草地村至北台山顶一线,对五台山南坡亚高山-高山山地进行植物群落调查。在土地利用、坡度和坡向等因子基本一致的生境条件下,大致以 100 m海拔为间隔,从海拔2200—3000 m左右范围内布设调查群落样地,共设置调查样地9个(表1)。五台山样地气候数据为1950—2000年的年均数据(www.worldclim.org/)(表 1)。在灌木-草本结构类型样地,灌木样方大小:5 m×5 m,每个样地3个重复;在每个灌木样方内随机设置3个草本样方,样方大小:1 m×1 m,调查全部物种。在草本结构类型样地,设置3个草本样方,样方大小:1 m×1 m。共布设21个灌木样方,69个草本样方。记录样地基本状况和群落属性,如经纬度、海拔、坡向、土地利用方式以及物种种类、株数、冠幅、高度和盖度等群落指标。

表1 不同海拔样地基本信息Table 1 Information of study plots at different elevations
1.3 数据分析
在群落调查的基础上,计算植物物种重要值,群落优势种由重要值决定(Zhang et al.,2018),公式如下:

依据吴征镒关于中国种子植物科、属的分布类型划分方法,对研究区内植物区系分布进行分析(吴征镒,1991;吴征镒等,2003)。采用单因素方差分析(One-way ANOVA)对不同样地群落盖度、α多样性指数的差异进行检验,多重比较采用LSD法(P<0.05),统计分析在SPSS 18.0中完成。
α多样性指数采用 Patrick丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数表示(余娇娥等,2018;赵峰侠等,2019),公式如下:
Patrick指数:

Shannon-Wiener多样性指数:

Simpson优势度指数:

Pielou均匀度指数:

式中,S为物种数,Pi为种i的相对重要值。
β多样性指数采用Sorensen相似性指数和Cody相异性指数表示(邓清月等,2019),公式如下:
Sorensen指数:

Cody指数:
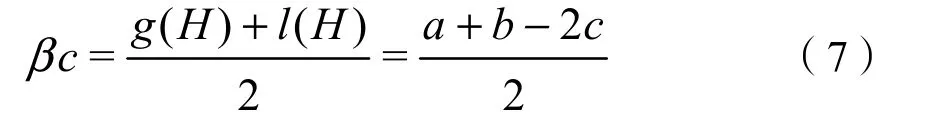
式中,a,b分别为相邻两样地各自的物种数,c为两样地之间的共有物种数。
2 结果与分析
2.1 植物群落组成及其主要特征
经统计,海拔梯度上9个样地共记录植物物种52种,隶属于19科44属。在海拔梯度上,植物科分布区类型以世界广布(78.9%)为主,北温带和南温带间断分布(10.5%)次之;属分布区类型以北温带分布(38.6%)为主(表2)。在海拔梯度上,群落结构相对简单,在2200—2800 m处以灌-草群落结构为主,在2800 m以上出现单一草本群落结构(表 1)。随着海拔的升高,依次为金露梅灌丛草甸群落(2201—2700 m)、鬼箭锦鸡儿灌丛草甸群落(2700—2800 m)、高山嵩草草甸群落(2800—3011 m)。

表2 研究区植物科、属分布区类型Table 2 The Areal-types of family and genera of plants in the studied plots
在海拔梯度上,不同海拔样地群落高度(F=58.73,P<0.001)、群落总盖度(F=11.03,P<0.001)、灌木层盖度(F=306.02,P<0.001)、草本层盖度(F=39.54,P<0.001)存在显著性差异,群落高度变化在11.10—75.8 cm,群落总盖度变化在73.0%—99.0%,灌木层盖度变化在0—86.3%,草本层盖度变化在47.40%—99.10%(表3)。群落高度随着海拔的升高则大致呈先下降后上升又下降的变化趋势,在3011 m处达到最小值,为12.3 cm(表1)。随着海拔的升高,群落总盖度呈先下降后上升的变化趋势,在3011 m处达到最大值,为98%(表1);灌木层盖度随海拔升高与群落总盖度变化趋势相反,呈先上升后下降变化趋势,在 2508 m处达到最大值,为80%,而草本层盖度随海拔升高与群落总盖度趋势相一致,呈先下降后上升的变化趋势,在2508 m处达到最小值,为55%(图1)。该区域植物群落草本层盖度总体上大于灌木层,均值分别为75.78%和41.64%(表3)。
2.2 群落α多样性沿海拔梯度的变化

图1 灌木层和草本层盖度沿海拔梯度的变化Fig.1 Variation in coverage of shrub layer and herb layer along the altitudinal gradient

表3 海拔梯度上群落特征和多样性指数的单因素方差分析Table 3 One-way ANOVA of community characteristics and diversity indices along the altitudinal gradient
在海拔梯度上,不同海拔样地群落Patrick指数(F=8.00,P<0.001)、Shannon-Wiener指数(F=13.94,P<0.001)、Simpson 指数(F=10.85,P<0.001)和Pielou指数(F=5.89,P<0.001)存在显著性差异,其中Patrick指数在6—20,Shannon-Wiener指数在0.88—2.23,Simpson指数在 0.41—0.84,Pielou指数在0.49—0.78(表3)。随着海拔的升高,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均呈显著下降趋势(图2)。
2.3 群落β多样性沿海拔梯度的变化
随着海拔的升高,植物群落间 Cody指数呈先迅速下降后又上升再下降的变化趋势,最小值出现在2903—3011 m之间,为2.5,在2201—2296 m之间和 2800—2903 m之间达到峰值,分别为 12和9,在2296—2800 m之间变化相对平稳(图3)。SI相似性系数与Cody系数随海拔变化相反,表现为先缓慢上升后下降再上升的变化趋势,最小值出现在2201—2296 m之间和2800—2903 m之间;最大值在2903—3011 m之间,为0.78(图3),即在2201—2296 m之间和2800—2903 m之间组分变化较大,物种更替速率较快,而在2903—3011 m之间物种变化相对较小。
3 讨论
在五台山亚高山-高山带南坡,海拔变化对灌丛
草甸植物群落特征的影响非常明显,随着海拔的升高,依次分布有金露梅灌丛草甸群落(2201—2700 m)、鬼箭锦鸡儿灌丛草甸群落(2700—2800 m)、高山嵩草草甸群落(2800—3011 m)。总体来看,重要值很好地反映出亚高山-高山带南坡灌丛草甸群落随海拔变化的变化。在南坡亚高山-高山带,灌木种主要分布在2200—2800 m之间,未在2800 m以上出现,这从侧面反映出灌木层生长和分布可能受海拔变化的制约(张金屯,1986)。在海拔梯度上,灌木层盖度随着海拔的升高呈先上升后下降的变化趋势,波动较大,而群落总盖度和草本层盖度呈相反变化趋势,且波动相对较小(图 1),这可能是由于该地区草本植物盖度占总盖度比例较大,使得群落总盖度主要受草本层的影响(张金屯,1986;江源等,2009)。此外,研究结果表明在2201—2508 m之间灌木层呈上升变化趋势,而草本层盖度呈下降变化趋势,这种变化趋势与贺兰山(苏闯等,2018)、长白山(夏富才等,2012)等研究结果相类似,这主要是灌木层盖度增加,导致了下层水热条件的重新配置,强烈的种间竞争导致草本层的多样性降低,使得草本层盖度有所降低(Wu et al.,2013;余娇娥等,2018)。此外,科的区系成分以世界广布为主,属的区系成分以北温带分布类型为主,这与晋西北饮马池山(邓清月等,2019)、五鹿山(曲波等,2012)、太岳山(王燕等,2017)等温带山区区系成分相类似,体现出该地区温带植被的特点。与以上山区有所不同,属的区系成分出现北极-高山分布类型(表2),反映出五台山亚高山-高山地带的高寒植被属性。

图2 α多样性指数沿海拔梯度的变化Fig.2 Variation of α diversity indices along the altitudinal gradient

图3 β多样性指数沿海拔梯度的变化Fig.3 Variation of β diversity indices along the altitudinal gradient
在山区,物种多样性随海拔升高变化主要表现为随海拔升高先降低后增加、先增加后降低、单调下降、单调升高和保持恒定不变的变化趋势(贺金生等,1997)。本文中五台山亚高山-高山地带南坡物种丰富度格局符合单调下降格局,表现为随海拔升高呈显著线性下降趋势(图 2),这与河北小五台山(白晓航等,2016)、北京东灵山(向春玲等,2009)等研究结果相类似,这种格局与本研究海拔范围和该区域南坡植被特征垂直分布密切相关。在海拔梯度(2200—3000 m)上,植被结构相对简单,灌木层物种数较少,变化不大,群落物种丰富度可能主要受草本植物的影响。在2201—2508 m之间,随着海拔的升高物种丰富度下降,这可能是由于海拔梯度上灌木层盖度逐渐增加,使得下层草本生存生长所必需的光照条件受限,导致其物种多样性逐渐降低(余娇娥等,2018;苏闯等,2018)。已有研究表明,当灌木层盖度超过一定阈值(约 40%),上层荫蔽使得草本植物生长所必需的光照减少,从而降低草本层物种多样性(苏闯等,2018)。在2201 m处,水热条件适宜,且灌木层盖度较低,促使群落物种丰富度达到峰值。随着海拔的升高,在2508—2800 m之间,灌木层盖度有所下降,使得群落物种丰富度有所回升。在2800 m以上,灌木层消失,然而高山地带风速大,日照时间长,土壤容易干旱缺水,加上气候寒冷,水热条件较差,仅有少数耐高寒物种存活(崔海亭,1983),如高山嵩草、珠芽蓼(Polygonum viviparum)等,使得物种分布有限,群落物种多样性较低,从而形成物种丰富度随海拔升高呈显著线性下降的变化趋势。
随着海拔的升高,植物群落 Shannon-Wiener指数、Simpson指数、Pielou指数和Patrick物种丰富度指数变化趋势一致,表现为随海拔升高呈显著线性下降趋势(图 2),这一结果基本上反映了五台山亚高山-高山带南坡不同植物群落在物种组成方面的差异,即随着海拔的升高,植物群落多样性趋于减少,群落内物种分布的均匀程度逐渐降低。然而在小五台山、东灵山等亚高山地区,Pielou指数出现随着海拔的升高呈上升的变化趋势(向春玲等,2009;白晓航等,2016),这与本研究结果不同,可能是与五台山高海拔地区特殊的气候和环境因子密切相关,高寒气候限制草本植物分布,形成以耐高寒草本植物占优势的高寒草甸群落(刘鸿雁等,2003)。野外调查结果也表明,随着海拔的升高,样地物种数目和种类逐渐减少,尤其在2800 m以上高山地区,植物群落以高山嵩草占优势,其它物种分布较少。
β多样性可以反映出不同生境间植物群落间的梯度变化,或是环境梯度中的物种更替速率(张晓龙等,2017;邓清月等,2019)。本研究中 Cody指数在2201—2296 m之间和2800—2903 m之间达到峰值,而SI相似性系数相对较小(见图3),表明以上海拔段之间β多样性最大,即物种更替速率相对较快,植物群落间组成差异性大,反映出2201—2296 m之间和2800—2903 m之间具有明显的过渡性质。该研究结果符合崔海亭(崔海亭1983)、江源(江源等,2009)等得出的在五台山南坡2300 m和2800 m左右为具有重要生态学意义的 2个海拔过渡带的结论,从侧面反映出亚高寒-高寒灌丛主要分布在2200—2800 m之间。野外调查结果也表明,南坡2200 m以下灌木层消失,该海拔段为中山草甸与亚高山灌丛草甸过渡地带;而2800 m以上出现以高山嵩草(重要值为 0.23—0.25)和珠芽蓼(重要值为 0.21—0.22)等为优势种的高山草甸群落,灌木层消失,该海拔段为亚高山-高山灌丛草甸与高山草甸的过渡地带。这些结果有助于揭示五台山亚高山-高山带南坡灌丛草甸群落多样性的变化特征,但结合环境因子监测,多个梯度的长期野外观测更有助于加深对亚高山-高山带灌丛草甸群落多样性变化机制的理解。
4 结论
五台山亚高山-高山带南坡植被盖度、植物群落多样性指数在海拔梯度上的分布格局表明:随着海拔的升高,灌木层和草本层盖度对海拔梯度响应不同,灌木层盖度随海拔升高呈先上升后下降变化趋势,而草本层与灌木层盖度变化趋势相反。在海拔梯度上,重要值很好地反映出亚高山-高山带南坡植物群落随海拔梯度的变化,依次为金露梅灌丛草甸群落(2201—2700 m)、鬼箭锦鸡儿灌丛草甸群落(2700—2800 m)、高山嵩草草甸群落(2800—3011 m)。植物群落α多样性指数随海拔升高呈显著下降的变化趋势;β多样性指数在2200 m和2800 m左右出现明显的波动,以上两个海拔段是重要的生态过渡地带,且2200—2800 m是亚高山-高山灌丛草甸主要的海拔分布范围,该海拔段可能是灌丛草甸植被生态恢复和保护的关键区段。
