硅肥对于提高植物抗性的作用机理研究进展
2020-07-14颜瑾刘晓敏何可佳
颜瑾 刘晓敏 何可佳


摘要 硅元素虽然不是植物的必需营养元素,但越来越多的研究结果表明,硅对于植物抵抗生物与非生物的胁迫具有重要作用。近年来,关于硅元素对植物的功能研究非常活跃。从硅元素能显著提高植物抵抗生物、非生物胁迫能力,硅肥根施和叶施的争论,硅在植物体内的运输及相关基因,硅介导植物产生系统抗性的分子机制等方面对硅元素在植物与病原物互作中的作用机理进行了综述,并提出了今后研究方向,为更合理利用硅肥提供一定的理论基础。
关键词 硅肥;植物抗病反应;硅转运蛋白
Abstract Although silicon is not an essential nutrient for plants,more and more research results show that silicon plays an important role in plantsresistance to biological and abiotic stresses.In recent years,research on the function of silicon on plants has been very active.This paper reviewed the mechanism of silicon in plant pathogen interaction from the aspects of silicon can significantly improve plant resistance to biotic and abiotic stresses,the debate on silicon application in roots and leaves,silicon transport in plants and related genes,and silicon mediated molecular mechanism of plant systemic resistance.The research direction in the future was put forward to provide a theoretical basis for more rational use of silicon fertilizer.
Key words Silicon fertilizer;Plant disease resistance response;Silicon transport protein
硅元素在地球中的含量非常丰富,硅作为地壳中第二大元素,含量仅次于氧,约占地壳的28%[1]。硅元素并不是传统意义的植物必需营养元素,到目前为止,硅元素并没有包含在任何已知植物物质能量代谢途径中,因此关于硅元素对植物生长的作用很长时间内并没有得到重视,一方面可能是由于硅元素在地壳中的含量非常丰富,人们认为无需担心土壤中硅元素不够的问题,另一方面要创造一个不含硅元素的试验环境来研究其对植物生长的作用非常困难。历史上很长一段时间内,人们对于硅元素在植物上的作用并不是很清楚。地球上虽然存在大量的硅,但绝大部分是以晶体形式存在的,不溶于水,并不能被植物直接利用,能被植物利用的硅是能溶于水的硅酸。在自然界的土壤中,硅酸浓度为0.1~0.6 mmol/L[1]。但不同的植物之间,根中硅含量相差非常大,低的只有0.1%,高的可达10%[2]。
由于硅元素长期以来并没有被作为植物的必需元素对待[3],所以一直以来并没有硅肥的概念。最早有科学记载硅元素在植物中含量的是在1804年,De报道了在禾本科植物中,硅含量是其他非单子叶植物的10~20倍[4]。化学家Davy[5]在1819年出版的《基础农业化学》一书中,描述了硅可能对植物抵抗昆虫具有重要作用。英国的Rothamsted试验站对施用硅酸纳对青草和大麦生长和产量的影响进行了研究,发现在缺磷或钾的地里施用硅酸钠可持续提高作物产量,但在磷和钾丰富的地方施用硅酸钠并没有增产效果[6]。20世纪以来,关于硅元素在植物中的作用研究越来越多,研究表明,硅元素能促进植物生长,增加产量,提高植物对生物、非生物胁迫,提高植物光合作用,降低重金属毒性等,但均以现象描述为主。直到2006年日本学者Ma在水稻中找到并克隆了第一个硅转运蛋白基因Lsi1,并证明了Lsi1基因负责将硅酸运输到根部细胞内,将Lsi1基因突变掉后,水稻中的硅含量顯著降低,水稻产量也显著下降,抗病性明显降低[7]。硅转运蛋白基因的发现为研究硅的生理功能提供了直接的分子证据,后来,陆续在其他植物中发现了类似的硅转运蛋白基因,从此,关于硅元素在植物中的功能研究进入分子时代。
在我国最早关于农业的一部著述《齐民要术》中有关于利用秸秆、动物粪便有机肥等循环农业措施来提高农作物产量的描述,这些有机肥中便含有硅元素的重新利用。但第一次科学试验报告是陈恩凤和朱淇在1963年所作出的,报告称施用硅肥后水稻增产10%,且稻米品质更好,同时在小麦、玉米、大豆等作物上也具有明显增产效果[8]。浙江大学梁永超等[6,9]提出,硅介导的抗氧化作用是植物抵抗非生物因素如盐、冻、干旱、重金属胁迫的一个主要作用机理,第一次发现和报道了在黄瓜中硅的吸收和转运过程,并指出在如水稻、向日葵中,硅的运输存在主动运输和被动运输两种途径,取决于土壤中可溶性硅的浓度,高浓度硅时以被动运输为主,低浓度硅时以主动运输为主,但将硅转动至木质部的过程是主动运输,与外界硅元素的浓度无关。
近年来,关于硅元素对植物的功能研究非常活跃,这里只针对硅元素在植物与病原物互作过程中的作用机理作一综述,希望能从中得到一些启发,为更合理利用硅肥提供一定的理论基础。
1 硅元素能显著提高植物抵抗生物、非生物胁迫能力
虽然硅对于大多数植物来说并不是传统意义上的必需营养元素,但越来越多的研究表明,硅在植物提高对生物、非生物胁迫的抗性中具有非常重要的作用,施用硅肥往往与提高植物的抗病性关联在一起,目前,在多种作物-病原物互作体系中均有报道。
在水稻上,硅对稻瘟病[10]、褐斑病[11]均有较好的抗病效果。
不同植物对硅的吸收和利用能力差异很大,一些植物对硅处理没什么反应,人们往往认为硅对于这些植物没有作用,但实际上是这些植物对硅的吸收能力弱,而不是硅在植物体内没有作用。很多研究结果表明,单子叶植物较双子叶植物利用硅元素的能力强。不同植物利用硅元素的能力与硅转运蛋白基因结构相关,在硅积累植物中,硅内转运蛋白基因存在一个特异的结构域NIPs (nodulin 26-like intrinsic proteins),NIPs是屬于MIPs(major intrinsic protein)家族蛋白的一员。在模式植物拟南芥中,缺少这一结构域,这也与之前的研究结果吻合,拟南芥对硅的利用能力非常有限。越来越多的试验结果表明,硅在植物体内的作用具有一定的共性[6],硅进入到植物体内均能提高对生物、非生物胁迫的抗性。
2 硅肥根施和叶施的争论
虽然越来越多的研究表明,硅肥对于农业增产、提高农产品质量具有非常重要的作用,但人们对硅肥的认识、开发、使用依然存在很多误区,其中一个争论的焦点便是硅肥是根施好还是叶施好。由于直接在植物根部施用硅酸比较困难,有人建议在叶部施用硅肥,甚至一些公司开发出了专门针对叶部施用的硅肥产品。关于叶部施用硅肥主要存在两个方面的争论——吸收和作用方式。现有的研究结果表明,植物对硅的吸收主要是通过根部吸收的,然后通过木质部运输到地上部分。目前没有证据表明叶部施用硅肥吸收效率比根部施用高,在大豆-锈病体系研究中,叶部施用硅肥虽然也可提高大豆抗锈病能力,但根部施用硅肥比叶部施用硅肥具有更好的保护效果[12]。
尽管目前所有的试验结果都表明,植物根部吸收硅的效率要比叶部高和具有更好的防病效果,但市场上和研究中依然在极力开发和推广叶用硅肥,这给人们在施用硅肥时带来一定的困惑,随着硅在植物体内的吸收、运输机制的进一步深入研究和解析,有望解决这一争论。这里需要指出的是,叶部施用硅肥可以提高植物的抗病性,特别是叶部病害如白粉病[13],但这类报告很少研究其背后的机理,而后有研究表明,在叶部喷施钾盐溶液,如碳酸钾或磷酸钾溶液同样对白粉病具有防效,甚至防效比硅肥还好。叶部施用硅肥对白粉病的防治原理可能是喷施的酸溶液在植物叶面形成的渗透压影响病原孢子的萌发[14]。Liang等[14]的研究结果也表明,植物并不能吸收叶部施用的硅肥。但必须承认,叶部喷施硅肥对某些病害具有一定的防效,但要清楚这种防效并不是由于硅的直接作用,而是盐溶液所形成的渗透压作用导致的。
3 硅在植物体内的运输及相关基因
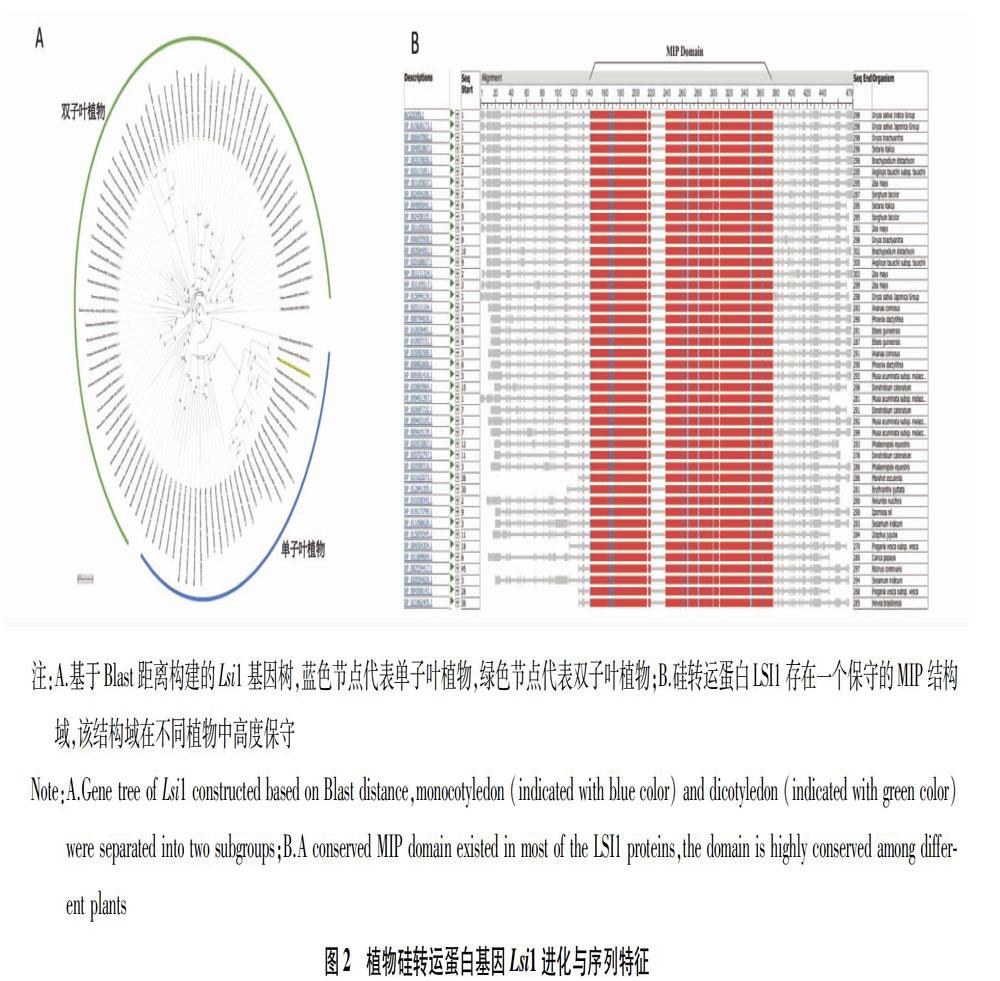
研究结果表明,植物吸收硅元素主要是通过根吸收土壤中的硅酸分子,然后通过木质部运输到地上部分。现在已经清楚硅元素在植物体内转运的分子机制,硅酸在植物体内的转运主要是通过一类硅特异性转运蛋白完成的,植物主要通过Lsi1从土壤中吸收硅酸分子,Lsi1将土壤中的硅酸分子通过主动运输内泵到植物外皮层细胞内,然后由另外一个基因Lsi2负责将吸收的硅酸分子从外皮层细胞泵出到表皮组织胞间,硅酸分子然后扩散进入木质部,随水分运输到地上部分,硅酸分子从木质部出来,还要通过另外一个Lsi6基因的作用将硅酸分子泵出到其他组织(图1)。硅积累植物,如水稻,要从环境中吸取大量的硅元素,Lsi1基因是一个关键基因,将Lsi1基因敲除或突变后,水稻体内硅含量显著降低,对生物、非生物胁迫的抵抗能力也大幅度降低[15]。目前Lsi1基因已在多种植物,包括水稻、玉米、小麦、燕麦、南瓜、黄瓜等中克隆得到,通过基因组数据分析,可以预测得到更多的植物Lsi1基因。研究结果表明,Lsi1基因在不同植物中变化较大,序列上的差异可能是导致不同植物利用硅能力差异的一个重要原因[16]。从Lsi1基因的距离树上可以推测出,Lsi1基因是在单、双子叶植物分化前就已经分化了(图2)。Lsi1基因大多在植物根部表达,但不同植物在根部表达的部位并不一致,水稻Lsi1主要在根的老熟部位表达,很少在根尖部位表达,而燕麦、玉米Lsi1基因在整个根部都有表达[17]。在模式植物拟南芥中并不存在Lsi1的同源物,这也与拟南芥对硅的利用能力非常有限相对应。试验表明,通过将小麦Lsi1基因转入拟南芥植株,转基因拟南芥植株体内可以积累大量硅,且表现出对生物、非生物胁迫强烈的抗性[18],这表明硅在植物体内的基因作用网络可能在大多数植物中是保守的。这也为培育抗性品种提供了新思路。
4 硅介导植物产生系统抗性的分子机制
植物体内积累的硅能普遍提高植物的抗性,这已在多种植物中得到充分证实,但不同植物对硅肥处理的反应不一样,通过前面对硅元素在植物体内运输分子机制的了解,可以知道Lsi1基因是影响植物从环境中吸收硅的决定性因素,不同植物由于Lsi1基因的差异,导致利用环境中硅的能力不一样。
关于硅能提高植物抗性的机理历史上曾经有两个假说,一个假说是认为硅能够提高植物细胞的稳固性,是直接对植物细胞加固,从而提高植物的抵抗力,属于物理防御。早期,研究者普遍认为存在于细胞壁和细胞间的硅是阻止病原微生物入侵植物的主要因素,但也有些学者不同意这种说法,早在1941年,日本学者Yoshi [19]指出,植物叶片的硬度与硅处理没有相关性,认为吸收硅后的物理防御不足以解释水稻抗稻瘟病。有学者通过扫描电镜(SEM)技术和X射线扫描黄瓜细胞壁,也没有找到硅能增强植物细胞壁抵抗病原菌侵染的证据[20]。
另一个假说是认为硅能够充当第二信使,能激发植物启动下游抗性反应。在寻找硅能提高植物细胞壁抵抗病原菌受阻后,研究者便开始思考是否存在其他理论来解释植物吸收硅后能提高其抗性,包括从生理、生化角度研究植物吸收硅后在这些方面的变化,最后也鉴定出多种化合物跟植物抗性相关,包括黄酮类化合物、植保素等物质。再后来,随着分子生物学的发展,开始从分子水平解析硅的作用机理,发现硅可以介导由MAPK参与的水杨酸(SA)信号传导途径起作用[21],激活一些与植物抗性相关基因表达,如病程相关蛋白基因(NPR1)。转录组结果也表明,硅处理对健康对照(没有胁迫)基因表达并没有明显差异,但当存在生物、非生物胁迫时,硅处理后基因表达差异显著,这说明在没有生物、非生物胁迫情况下,硅如果只是作为一种肥料施用,并不会有什么效果,只有当植物遭受生物、非生物胁迫时,硅的作用效果才显现出来[22]。虽然转录组可以很好地反映硅处理后植物基因表达差异,从基因组水平上了解基因表达动态,但并不能解释硅的作用机理。而且越来越多的研究表明,硅并不能诱导植物产生防御反应[23]。图3为硅介导的植物抗病反应机理推测。
拟南芥虽然在很多方面都是很好的模式植物,但在研究硅的作用机理方面,拟南芥并不是一个很好的模式植物,因为研究表明,拟南芥基因组中缺少硅运输的关键基因Lsi1,这也与拟南芥中硅的含量很低相吻合。但有研究者通过转基因的方法,将小麦的Lsi1基因转到拟南芥得到转基因拟南芥,结果表明,转入外源Lsi1基因的拟南芥对硅的吸收能力大幅度提高,更为重要的是,转基因拟南芥对白粉病的抗性也大幅度提高,这意味着硅在植物体内的作用机理可能存在共性[24],不同植物硅的作用效果不同可能是由于對硅的转运能力差异导致的,这也为培育抗病品种提供了新的思路。
之前的研究表明,硅主要是对活体寄生菌(如白粉菌)[25-26],或半活体寄生菌(稻瘟菌)[10,27]具有防御作用,这类病原物大多会产生一种被称为效应子的蛋白或物质,这种效应子在感病植株上表现为能抑制植物抗病反应,而在抗病植株上表现为能启动抗病反应,对病原菌来说,效应子是一把“双刃剑”。有研究结果表明,硅大量富集在病原菌侵入部位[28],推测可能与病原物的效应子相互作用,从而降低了效应子的活性,不能有效抑制植物的抗病反应,植物在病原菌侵入后能启动抗病反应,从而提高了植物的抗性。硅介导的抗病反应在不同植物中具有一定的普遍性,这意味着它不是通过特异性互作启动下游的抗病反应,而是启动植物的基础抗性,即PTI,而效应子启动抗病基因的抗病反应是ETI,植物R基因激发的植物抗病反应通常属于ETI。
综上所述,硅增强植物抗性的分子机理可能是硅经根吸收进入植物细胞后,经木质部运输到地上部分,在没有胁迫情况下,硅在植物细胞内并没有明显作用,但当植物受病原菌侵染后,硅在病原菌侵入部位大量富集,与病原菌的效应子互作,阻挡效应子作用,降低效应子抑制植物的防御反应,植物可以启动正常的防御反应,从而增强植物的抗病性(图3)。这与之前的两种假说不同,硅是作为与效应子互作因子,降低了效应子的抑制作用,从而提高了植物的抗病性。
5 展望
近年来,硅肥作为第四大元素肥受到越来越多人的认可,施用硅肥不仅可以提高作物的产量,而且能提高作物的抗性,包括生物和非生物的抗性。但由于其内在机制不清楚,这在一定程度上限制了硅肥的应用。目前,对于硅元素在植物体内的运输已经初步清楚,在多种植物中克隆了编码硅运输蛋白的基因,但硅元素在植物体内是如何发挥作用的,硅在植物体内与哪些蛋白互作,如何调控植物抗性依然不清楚。要回答这些科学问题,需要科学家从分子水平上鉴定更多相关基因的功能。现在已经进入基因组后时代,越来越多的植物基因组序列得到了测定,这为从基因组水平上研究硅元素在植物体内的运输、互作、调控提供了可能。通过比较基因组学、转录组及代谢组等多组学联合研究技术,解析硅敏感植物与硅非敏感植物在功能基因上的差异,比较硅缺乏、硅过量情况下不同植物的基因转录水平,找出与硅相互作用的基因调控网络,另外,硅在植物的不同组织中积累不一样,这与硅的运输是密切相关的,但是如何调控的,这些科学问题的解答可以帮助人们理解硅元素对于提高植物抗性的分子机理,从而为更好地利用硅肥提供理论基础。
参考文献
[1] SAVANT N K,SNYDER G H,DATNOFF L E.Silicon management and sustainable rice production[J].Advances in agronomy,1996,58:151-199.
[2] MA J F,YAMAJI N.Silicon uptake and accumulation in higher plants[J].Trends in plant science,2006,11(8):392-397.
[3] ARNON D I,STOUT P R.The essentiality of certain elements in minute quantity for plants with special reference to copper[J].Plant physiology,1939,14(2):371-375.
[4] DE SAUSSURET.Recherches chimiques sur la vegetation[M].Paris:Nyon,1804.
[5] DAVY H.Elements of agricultural chemistry[M].Hartford:Hudson and Co.,1814.
[6] LIANG Y C,NIKOLIC M,BLANGER R,et al.Silicon in agriculture[M].Dordrecht:Springer,2015.
[7] MA J F,TAMAI K,YAMAJI N,et al.A silicon transporter in rice[J].Nature,2006,440(7084):688-691.
[8] 朱淇, 陈恩凤.钢铁炉渣的性质及施用于不同土类中对农作物的作用[J].土壤学报,1963,11(1):70-83.
[9] LIANG Y C,SI J,RMHELD V.Silicon uptake and transport is an active process in Cucumis sativus[J].New phytologist,2005,167(3):797-804.
[10] CAI K Z,GAO D,LUO S M,et al.Physiological and cytological mechanisms of siliconinduced resistance in rice against blast disease[J].Physiologia plantarum,2008,134(2):324-333.
[11] NING D F,SONG A L,FAN F L,et al.Effects of slag-based silicon fertilizer on rice growth and brown-spot resistance[J].PLoS One,2014,9(7):1-9.
[12] RODRIGUES F A,DUARTE H S S,DOMICIANO G P,et al.Foliar application of potassium silicate reduces the intensity of soybean rust[J].Australasian plant pathology,2009,38(4):366-372.
[13] MENZIES J,BOWEN P,EHRET D,et al.Foliar applications of potassium silicate reduce severity of powdery mildew on cucumber,muskmelon,and zucchini squash[J].Journal of the American society for horticultural science,1992,117(6):902-905.
[14] LIANG Y C,SUN W C,SI J,et al.Effects of foliarand rootapplied silicon on the enhancement of induced resistance to powdery mildew in Cucumis sativus[J].Plant pathology,2005,54(5):678-685.
[15] CHEN X W,SHANG J J,CHEN D X,et al.A Blectin receptor kinase gene conferring rice blast resistance[J].The plant journal,2006,46(5):794-804.
[16] VATANSEVER R,OZYIGIT I I,FILIZ E,et al.Genomewide exploration of silicon (Si) transporter genes,Lsi1 and Lsi2 in plants;insights into Siaccumulation status/capacity of plants[J].BioMetals,2017,30(2):185-200.
[17] YAMAJI N,MA J F.Spatial distribution and temporal variation of the rice silicon transporter Lsi1[J].Plant physiology,2007,143(3):1306-1313.
[18] MONTPETIT J,VIVANCOS J,MITANIUENO N,et al.Cloning,functional characterization and heterologous expression of TaLsi1,a wheat silicon transporter gene[J].Plant molecular biology,2012,79(1/2):35-46.
[19] YOSHI H.Studies on the nature of rice blast resistance[J].Japanese journal of phytopathology,1941,11(2):81-88.
[20] SAMUELS A L,GLASS A D M,EHRET D L,et al.Mobility and deposition of silicon in cucumber plants[J].Plant,cell & environment,1991,14(5):485-492.
[21] VIVANCOS J,LABB C,MENZIES J G,et al.Siliconmediated resistance of Arabidopsis against powdery mildew involves mechanisms other than the salicylic acid (SA)dependent defence pathway[J].Molecular plant pathology,2015,16(6):572-582.
[22] GHAREEB H,BOZS Z,OTT P G,et al.Transcriptome of siliconinduced resistance against Ralstonia solanacearum in the silicon nonaccumulator tomato implicates priming effect[J].Physiological and molecular plant pathology,2011,75(3):83-89.
[23] LIANG Y C,SUN W C,ZHU Y G,et al.Mechanisms of siliconmediated alleviation of abiotic stresses in higher plants:A review[J].Environmental pollution,2007,147(2):422-428.
[24] DESHMUKH R,BLANGER R R.Molecular evolution of aquaporins and silicon influx in plants[J].Functional ecology,2016,30(8):1277-1285.
[25] FAUTEUX F,CHAIN F,BELZILE F,et al.The protective role of silicon in the Arabidopsispowdery mildew pathosystem[J].Proceedings of the national academy of sciences,2006,103(46):17554-17559.
[26] SHETTY R,JENSEN B,SHETTY N P,et al.Silicon induced resistance against powdery mildew of roses caused by Podosphaera pannosa[J].Plant pathology,2012,61(1):120-131.
[27] RODRIGUES F ,BENHAMOU N,DATNOFF L E,et al.Ultrastructural and cytochemical aspects of siliconmediated rice blast resistance[J].Phytopathology,2003,93(5):535-546.
[28] MA J F,YAMAJI N,TAMAI K,et al.Genotypic difference in silicon uptake and expression of silicon transporter genes in rice[J].Plant physiology,2007,145(3):919-924.
