烟草HQT 基因超量表达对次生代谢物质一咖啡酰奎尼酸和黄酮醇合成的影响
2020-07-11丁香玉赵漫王鹏伟丁新华储昭辉赵翔宇李洋
丁香玉,赵漫,王鹏伟,丁新华,储昭辉,赵翔宇,李洋 ,
1 作物生物学国家重点实验室/山东农业大学植物保护学院,泰安市岱宗大街61号 271018;2 山东农业大学生命科学学院,泰安市岱宗大街61号 271018
羟基肉桂酰辅酶A:奎尼酸羟基肉桂酰转移酶(Hydroxycinnamoyl-CoA:quinate hydroxycinnamoyl transferase,HQT)是一类酰基-辅酶A-依赖的酰基转移酶,具有酰基受体的特异性,是催化一咖啡酰奎尼酸最后一步生物合成的关键酶,催化咖啡酰辅酶 A和奎尼酸进行酯交换生成一咖啡酰奎尼酸[1]。
一咖啡酰奎尼酸是许多植物中重要的苯丙酸,包括烟草,番茄,马铃薯,苹果等[2-4]。根据咖啡酰基在奎尼酸上的结合部位不同,一咖啡酰奎尼酸可分为多种异构体:1-咖啡酰奎尼酸(1-CQA)、3-咖啡酰奎尼酸(3-CQA)、4-咖啡酰奎尼酸(4-CQA)和5-咖啡酰奎尼酸(5-CQA,CGA,绿原酸)。1-CQA 经研究发现是 NF-ĸB 途径的潜在抑制因子,具有抗癌能力。3-CQA 在人类膳食中普遍存在,可以有效的预防肝脏系统疾病、高血压血脂症、肥胖等疾病[5-6]。4-CQA则具有非常强的消炎作用[7]。5-CQA 是其中最主要的一种同分异构体,在人体健康中发挥重要作用,能够减缓葡萄糖向血液中的释放,具有预防、降低心脏病和Ⅱ型糖尿病风险的作用[8-10]。绿原酸还是重要的天然抗氧化剂,能够帮助植物细胞抵御低温、紫外线等各种非生物胁迫以及病原,害虫侵害等生物胁迫[11]。目前在烟草叶片中只检测出3-CQA,4-CQA 和5-CQA 三种一咖啡酰奎尼酸[12]。
黄酮醇作为重要的苯丙烷类物质,广泛存在于多种茄科作物中,包括烟草,番茄,马铃薯等[13]。在烟草中主要是芦丁和山奈酚芸香糖苷,但含量极少,在人体中具有强抗氧化、清除人体自由基和抗菌消炎等多种作用[14]。对植物自身而言,黄酮醇类物质能够增强植物抗性,帮助植物抵抗各种生物和非生物胁迫,如病虫侵害和环境逆境等[15-17]。本文旨在分析HQT 基因对烟草次生代谢物质的影响,获得富含一咖啡酰奎尼酸的转基因烟草,并探究HQT 基因对黄酮醇生物合成的影响,对烟草次生代谢机制进行初步研究,为培育富含营养物质且具有更强抗逆能力的烟草做基础铺垫。
1 材料与方法
1.1 材料
三生烟(Nicotiana tabacum cv. samsun),植株在25℃,16 h/8 h 光暗周期,相对湿度为(70±10)%的温室内培养。烟草在营养钵中生长至8 叶期,取中部叶片进行检测。
大肠杆菌Escherichia coli DH5ɑ,农杆菌LBA4404和真核表达载体pCXSN 均由本实验室保存提供。
1.2 方法
1.2.1 NtHQT 基因克隆和表达载体构建
采用TRI Reagent (Sigma,美国)提取三生烟草(Nicotiana tabacum cv. samsun)叶片总RNA,取1 μg 总RNA 用DNaseI(NEB,美国)37℃处理15 min,75℃酶灭活10 min,去除基因组DNA,作为RNA 模板。根据反转录试剂盒(TOYOBO,日本),向RNA 模板中加入2 μL 5 × RT Master Mix Ⅱ,RNase-free ddH2O 补充至10 μL,37℃反转录15 min,98℃酶灭活5 min,获得cDNA,置于-20℃保存。
以上述反应得到的cDNA 为模板,根据烟草HQT基因序列(GeneBank AJ582651)和EST 序列特征,设计NtHQT 基因引物,引物序列为NtHQT-F:5’-ATGGGAAGTGAAAAAATGAT-3’ 和NtHQT-R:5’-CAAAATTCATACAAATACTTCT-3’ (华大基因股份有限公司,深圳)。采用LAmp 高保真DNA 聚合酶(康为,北京)进行扩增,反应体系为50 μL:50 ng 模板,1 U DNA 聚合酶,5 μL 10×PCR 缓冲液,5 μL dNTPs (2 mmol/L),1 μL 引物(10 μmol/L),3 μL MgSO4,ddH2O 补充至50 μL。PCR 反应条件:94℃预变性3 min,35 个循环:94℃变性30 s,56℃退火30 s,68℃延伸90 s(30-60 s/kb),扩增产物切胶回收。回收后NtHQT 基因片段和实验室保存的表达载体pCXSN,用Xcm I(NEB,美国)分别进行酶切,纯化后将载体和目的基因片段用T4 DNA 连接酶(ThermoFisher Scientific,美国)进行连接,形成遗传转化载体pCXSN::NtHQT。转化载体热激转入大肠杆菌DH5ɑ,挑白斑进行菌落PCR 初筛,挑选阳性克隆过夜扩繁,提取质粒送至公司(华大基因股份有限公司,深圳)测序。经测序无误后,转化农杆菌LBA4404。
1.2.2 农杆菌介导的烟草遗传转化
选取烟草叶片作为外植体,采用农杆菌(LBA4404)介导的叶盘转化法[18],将已构载体pCXSN::NtHQT转化三生烟草受体材料。待生根培养基中转基因烟草根系发育良好时,移栽到土壤中。采用C TA B[19]法提取植株叶片基因组D N A,以3 5 S 启动子内部序列设计的引物35S-F:5’-ACGCACAATCCCACTATCCTT-3’和基因内部序列设计的引物N t H Q T - R :5’-TCAAAATTCATACAAATACTTCT-3’进行PCR阳性检测。PCR产物经1%琼脂糖凝胶电泳分析扩增出1438 bp片段,为阳性植株。
1.2.3 一咖啡酰奎尼酸和黄酮醇物质提取和HPLC 含量测定
取转基因和野生型烟草叶片用液氮研磨至粉末状,各取0.3 g,3 mL 100%色谱级甲醇-20℃萃取2 h,每15 min 振荡1 次。4℃、4000 r/min 离心10 min,上清经0.22 μm 有机相微孔滤膜过滤至样品瓶,取10 μL 过滤液用于HPLC 检测分析[20-21],标准品为芦丁,1-咖啡酰奎尼酸,3-咖啡酰奎尼酸,4-咖啡酰奎尼酸,5-咖啡酰奎尼酸(Sigma),山奈酚芸香糖苷(Extrasynthese)。野生型和转基因烟草生长至8 叶期,取中部3 片真叶进行检测。T0代烟草每株检测重复3 次;T1代烟草每个株系3 个生物学重复。
2 结果
2.1 NtHQT 基因克隆和基因表达载体构建
按照三生烟草中NtHQT 的已知序列设计引物,以三生烟草叶片cDNA 为模板,通过PCR 扩增获得长为1311 bp 的清晰目标序列(图1A),切胶回收后酶切,连接Xcm I 酶切后的pCXSN 载体,热激转化大肠杆菌DH5ɑ,通过35S 启动子内部序列引物35S-F 和基因序列内部引物NtHQT-R 对白色克隆进行PCR 阳性验证(图1B),扩增出片段长度为1438 bp 的目标条带,选取3 个阳性克隆过夜扩繁后提质粒测序。
将测序结果与目标序列NtHQT(GeneBank AJ582651)和pCXSN 载体35S 启动子进行比对,比对成功,确认成功克隆NtHQT 的完整基因序列,并成功构建pCXSN::NtHQT 载体。
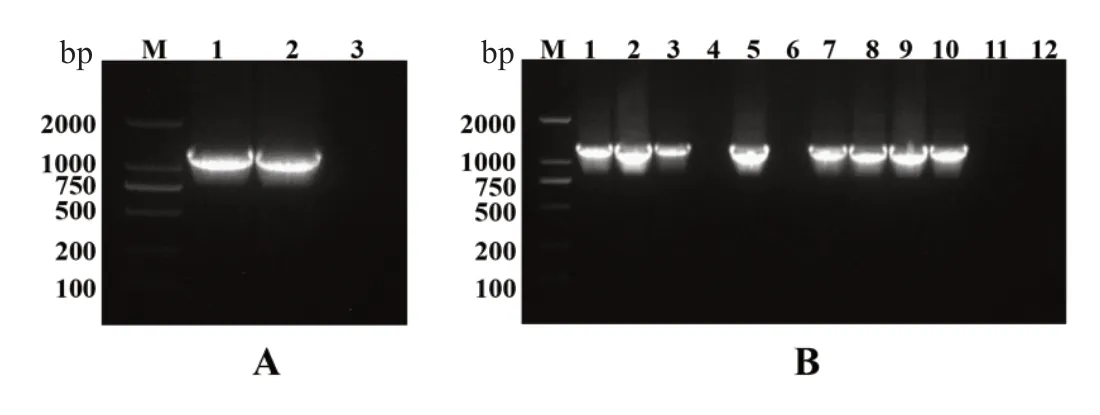
图1 NtHQT 片段扩增电泳及菌落PCR 检测图Fig. 1 NtHQT fragment amplification electrophoresis and PCR detection of colony
2.2 NtHQT 基因遗传转化和转基因烟草的获得
将构建成功的遗传转化载体电转农杆菌LBA4404,通过叶盘转化法转入三生烟草中,获得转NtHQT 基因烟草。通过PCR 对转基因烟草进行阳性检测(图2),在获得的15 株转基因材料中,有11株扩增到了长度为1438 bp 的目标条带,确认为阳性转基因植株,转化率为73.3%。转基因烟草在表型上与野生型没有明显差异(图3)。

图2 T0 代 转NtHQT 基因烟草阳性检测Fig. 2 Positive detection of NtHQT transgenic tobacco in T0 generation

图3 T1 代三个不同株系转基因烟草与野生型烟草表型Fig. 3 Phenotype between three different lines of NtHQT transgenic tobacco in T1 generation and wild-type tobacco
2.3 烟草叶片一咖啡酰奎尼酸和黄酮醇物质含量检测
待烟草叶片生长至8 叶期,参照Luo[21]等HPLC 检测苯丙烷类物质含量的方法,对部分转基因和非转基因烟草叶片进行含量检测。结果显示,转NtHQT 基因的烟草叶片中一咖啡酰奎尼酸、芦丁、山奈酚芸香糖苷的含量都有不同程度的提高(图4,表1)。

图4 T0 代转基因烟草叶片一咖啡酰奎尼酸及黄酮醇含量HPLC 检测 Fig. 4 HPLC analysis of CQAs and flavonols contents in NtHQT transgenic tobacco leaves in T0 generation and wild-type
其中,转NtHQT 基因的烟草叶片一咖啡酰奎尼酸的含量提高了4.04~7.28 倍,最高含量达487.00 μg/g (FW)。芦丁和山奈酚芸香糖苷含量最高分别可达208.00 μg/g (FW)和52.00 μg/g (FW),分别是相同条件下野生型三生烟草叶片中含量的17.83倍和4.3倍。
在T0代阳性植株中随机选择3 个株系,编号为NtHQT-1,NtHQT-2 和NtHQT-3,收取种子后进行T1代种植及阳性检测。每个株系取3 棵生物学重复,对阳性植株叶片进行含量测定。转基因烟草T1代植株三种物质含量变化倍数与T0代基本一致,说明转NtHQT 基因烟草富含一咖啡酰奎尼酸及黄酮醇的优势可以稳定遗传(图 5,表 2)。
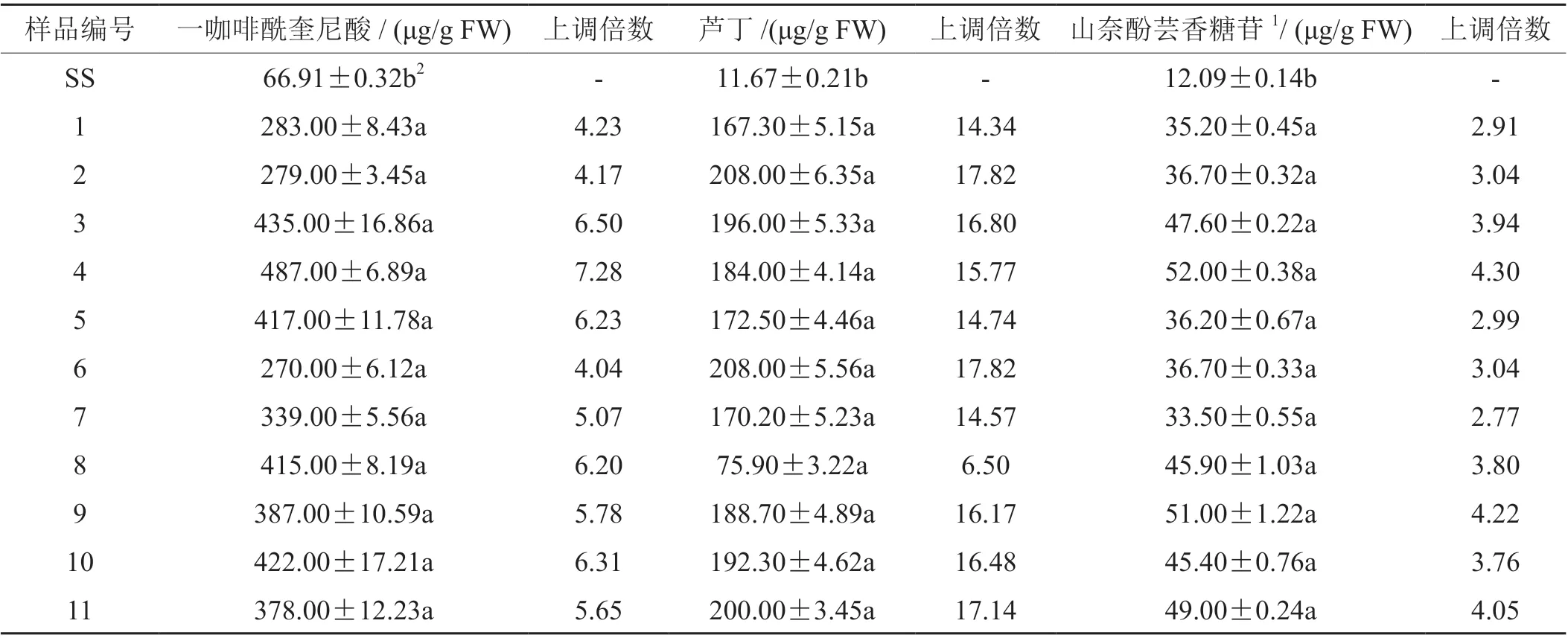
表1 转基因烟草T0 代叶片中各物质含量Tab. 1 Quantification of CQAs and flavonols in NtHQT-expressing tobacco leaves in T0 generation and wild-type (SS)
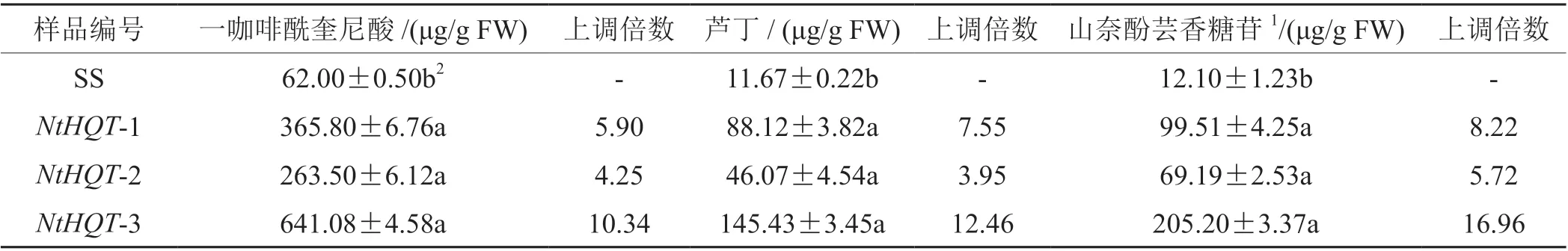
表2 转基因烟草T1 代叶片中各物质含量Tab. 2 Quantification of CQAs and flavonols in NtHQT-expressing tobacco leaves in T1 generation and wild-type (SS)
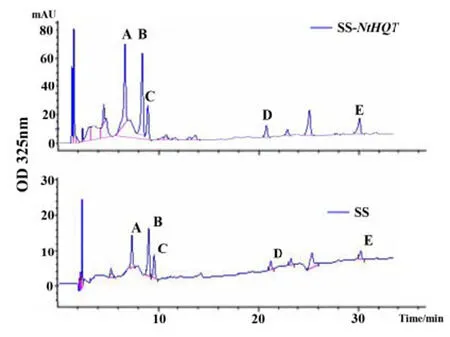
图5 T1 代转基因烟草叶片一咖啡酰奎尼酸及黄酮醇含量HPLC 检测Fig. 5 HPLC analysis CQAs and flavonols contents of NtHQT transgenic tobacco leaves in T1 generation and wide-type
3 结论与讨论
已有文章报导,HQT 基因参与多种植物绿原酸的生物合成。Lepelley M 等通过分析咖啡中HQT 的时空表达模式,发现其与绿原酸积累和分布的动态变化有关[1]。Raja S 在马铃薯中通过RNAi 对HQT 的抑制导致绿原酸下降90%以上[22],结果均表明NtHQT基因可以正向调控绿原酸的生物合成。本研究将烟草自身的HQT基因连接35S启动子构建真核表达载体,由农杆菌介导转化后得到的转基因烟草,不仅绿原酸含量明显提高,其他两种一咖啡酰奎尼酸含量也大幅度提高。而一咖啡酰奎尼酸不仅能够帮助植物增强对各种生物及非生物胁迫的抗性[11],还在人体健康中发挥重要作用,能够抗菌消炎,预防肥胖和心脑血管疾病等[5-6]。一咖啡酰奎尼酸所具有的功效,在一定程度上有可能降低吸烟对消费者造成的衰老、心血管疾病等危害。
芦丁,山奈酚芸香糖苷均具有较强的抗氧化活性,在人体中具有清除自由基,抵抗衰老的功能;在植物中能够增强植物对多种病原菌的抗性[23],帮助植物抵御多种生物与非生物胁迫,但在烟草中含量很少。本实验获得的转NtHQT 基因烟草,除检测到一咖啡酰奎尼酸含量最高可提高10.34 倍外,芦丁,山奈酚芸香糖苷含量也有大幅度提高,暗示黄酮醇合成相关通路可能被激活。
已有研究表明,芦丁和山奈酚芸香糖苷等黄酮醇物质合成与一咖啡酰奎尼酸合成路径共享多种中间酶和前期次生代谢物:以苯丙氨酸为合成底物,经过PAL(L-苯丙氨酸解氨酶)、C4H(肉桂酸羟化酶)、4CL(p-香豆酰-CoA 合成酶)催化合成p-香豆酸辅酶A。然后以p-香豆酰辅酶A 为底物,通过多种酶分别催化合成一咖啡酰奎尼酸,芦丁和山奈酚芸香糖苷等[21]。结合实验结果,转HQT 基因烟草叶片中一咖啡酰奎尼酸的含量最高可提高10.34 倍,芦丁和山奈酚芸香糖苷含量最高可分别提高17.83和16.97倍,推测,烟草HQT 基因的超量表达可能激活一咖啡酰奎尼酸合成路径中上游PAL、C4H、4CL 酶以及前期代谢物质苯丙氨酸和p-香豆酸辅酶A 的积累,这些合成相关酶及代谢物质的增多,同时促进下游多种黄酮醇合成酶的反应,从而更强激活芦丁和山奈酚芸香糖苷合成路径,而这些黄酮醇合成途径中关键酶基因的表达变化情况有待进一步研究。
通过对烟草一咖啡酰奎尼酸合成关键基因HQT的研究发现,HQT 基因不仅参与调控一咖啡酰奎尼酸的合成,也正向调控烟草中芦丁和山奈酚芸香糖苷的生物合成,有助于了解烟草中次生代谢物质,尤其是一咖啡酰奎尼酸及黄酮醇物质的合成机理,并为在实际生产中提升烟草品质和抗逆能力提供研究基础。
