miRNA-lncRNA影响人NK细胞与树突状细胞免疫功能和乳腺癌细胞侵袭力的分子机制
2020-07-01李雅怡张雨洁卓文杏
李雅怡 张雨洁 卓文杏
乳腺癌是最常见的恶性肿瘤之一。虽然早期检测,手术和化疗方面取得显著进步,但乳腺癌患者的5年生存率仍然很低[1]。自然杀伤细胞(natural killer cells,NK)和树突状细胞(dendritic cell,DC)是先天免疫的重要组成部分。增加的NK和DC介导的免疫反应与癌症患者的结果呈正相关,故迫切需要找到针对NK细胞与DC免疫功能以及乳腺癌细胞增殖和转移的新靶标。长非编码RNA(Long non-coding RNA,LncRNA)是一组具有超过200个核苷酸的非编码RNA。已经在各种类型的癌症中观察到lncRNA的异常表达,并且在肿瘤进展中起到关键调节剂的作用[2]。lncRNA SNHG7通过抑制P15和P16的表达促进了乳腺癌细胞的增殖并抑制了细胞的凋亡[3]。据报道,lncRNA UCA1通过miR-195/ARL2信号通路增强膀胱癌的线粒体功能和细胞活力。lncRNA在肿瘤发生过程中起着不同的作用。lncRNA可以作为致癌基因或肿瘤抑制基因发挥作用[4]。在这些肿瘤抑制因子lncRNA中,lncRNA MEG3(母系表达的基因3)引起了很多关注。MEG3过表达通过PI3K途径抑制子宫内膜癌细胞的增殖,侵袭和转移[5]。 MEG3通过调节WNT/β-catenin信号通路抑制口腔鳞状细胞癌的增殖和转移[6]。MEG3可通过靶向microRNA(miRNA)来调节肿瘤进展,miRNA是一组小的非编码RNA[7]。MEG3通过抑制miR-183的表达抑制人神经内分泌肿瘤细胞的生长和转移。MEG3通过靶向miR-93和抑制PI3K/AKT信号通路抑制胶质瘤细胞生长[8]。然而,MEG3是否通过靶向miRNA调节乳腺癌中的肿瘤进展尚不清楚。生物信息学预测显示miR-21是乳腺癌中MEG3的靶标,过表达的MEG3可通过抑制miR-21的表达来抑制NK细胞与树突状细胞免疫功能和乳腺癌细胞侵袭力。本研究可能会为乳腺癌的治疗找到新的药物靶点,报告如下。
1 资料与方法
1.1 一般资料 选择我院30例接受乳腺癌手术治疗的患者,这些患者在2015年9月至2017年3月治疗期间,未接受化疗或放疗。获得了乳腺癌组织和邻近正常组织,将组织立即在液氮中冷冻并储存在-80℃。
1.2 纳入与排除标准
1.2.1 纳入标准:①年龄30~60岁;②性别不限;③患者及全部接受乳腺癌手术。
1.2.2 排除标准:①有肝、肾、造血系统等严重原发性疾病者;②在组织采集前接受化疗或放疗的患者;③自身免疫性疾病患者。
1.3 方法 实验分为4组:乳腺癌组织,正常组织,乳腺癌细胞系,正常细胞系。
1.3.1 乳腺癌细胞系和人胃上皮细胞系:乳腺癌细胞系(MCF7)和人乳腺上皮细胞系DU4475购自中国科学院细胞库,并在RPMI1640培养基(Invitrogen公司,美国)中培养,补充有10%FBS(Gibco公司,美国),37℃,5%CO2。
1.3.2 定量实时聚合酶链反应(qRT-PCR):使用TRIzol试剂(Thermo Fisher Scientfic)按照说明从细胞或组织中提取总RNA。使用PrimeScript TM RT-PCR试剂盒(TaKaRa,Kusatsu,Japan)通过使用1μg总RNA逆转录成cDNA。为了检测MEG3和miR-21的表达,分别使用SYBR Premix Ex Taq试剂盒(Thermo Fisher Scientific)和TaqMan miRNA测定法(TaKaRa)进行RT-PCR。 GAPDH和U6分别用作lncRNA和miRNA的内部上样对照。通过相对量化(2-ΔΔCt)方法计算相对水平。所有实验重复至少3次。
1.3.3 细胞转染:为了增强MEG3外源表达,根据人MEG3(NR_002766)基因序列的GenBank,通过PCR合成MEG3序列的全长,并插入到pcDNA3.1空质粒(GenePharma公司,上海)中,称为MEG3转染。空质粒作为对照(pcDNA3.1载体)。将乳腺癌细胞接种到96孔板中并培养以达到60%的转染影响。根据Lipofectamine 2000(Thermo Fisher Scientific公司)制造商的说明书进行转染。用于转染的miR-21模拟物和miR-21对照的浓度为50 nmol/L。用于转染的MEG3和对照载体的浓度为100 nmol/L。见表1。

表1 探针序列
1.3.4 细胞增殖试验:细胞计数试剂盒-8(CCK-8)测定用于细胞增殖测定。将细胞接种在96孔板(每孔1×104个细胞)中,在37℃,5%CO2湿润的培养箱中培养24 h。向每个孔中加入10 μl细胞计数试剂盒-8溶液(CCK-8)(Dojindo Laboratories公司,日本),并在37℃,5%CO2湿润的培养箱中孵育2 h。光谱仪Varioskan Flash用于测量450 nm吸光度。所有实验重复至少3次。
1.3.5 迁移与侵袭测定:使用24孔transwell板(Corning公司,美国)进行Transwell测定以研究细胞的迁移和侵袭能力。对于侵袭测定,上室用50 μl基质胶基质包被。将数量为1×105的细胞以100 μl无血清培养基接种到上室中,其中600 μl DMEM在下室中补充有10%FBS。培养18 h后,用棉签除去上室中的细胞,同时用Wright's-Giemsa染色法对下表面上的入侵或迁移细胞染色,并计数。对迁移实验,添加5×104个细胞并且不使用基质胶基质。所有实验重复至少3次。
1.3.6 生物信息学方法:MiRDB生物信息学工具用于预测研究中推定的MEG3靶标。
1.3.7 双荧光素酶报告基因测定:通过RT-PCR扩增含有miR-21靶序列的MEG3片段,然后插入pmirG10双荧光素酶miRNA靶表达载体(Promega公司,美国)以形成报告载体MEG3-野生型(MEG3-WT)。另外的表达载体也通过插入突变的结合位点构建,并命名为MEG3-突变型(MEG3-MUT)。使用Lipofectamine 2000将MEG3-WT或MEG3-MUT和miR-21模拟物共转染到乳腺癌细胞系中,并使用双荧光素酶报告分析系统(Promega公司,美国)测试荧光素酶活性。
1.3.8 蛋白质印迹分析:通过使用RIPA裂解缓冲液(碧云天公司,上海)从细胞中提取蛋白质,并通过Bradford Protein Assay Kit(碧云天公司,上海)定量蛋白质浓度。通过10%SDS-PAGE凝胶分离等量的蛋白质,然后转移到PVDF膜(EMD Millipore公司,美国)中。在室温下用5%脱脂乳封闭1 h后,将膜与得自Abcam的一抗(抗-CD56;抗-CD80;抗-MMP2;抗-MMP-9和抗-GAPDH)在4℃下过夜。在室温下与辣根过氧化物酶(HRP)-连接的第二抗体温育1 h后,通过电化学发光试剂盒(碧云天公司,上海)显现信号强度。所有实验重复至少3次。
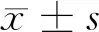
2 结果
2.1 乳腺癌组织和细胞系中MEG3下调表达 qRT-PCR分析乳腺癌组织和正常组织中lncRNA(MEG3)的表达,与正常组织相比,乳腺癌组织中MEG3表达下调(P<0.05);与正常细胞系比较,乳腺癌细胞系中MEG3的相对表达降低(P<0.05)。表明MEG3在乳腺癌组织和细胞系中下调表达。见表2,图1。
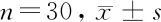

类别lncRNA正常组织 0.82±0.14乳腺癌组织 0.37±0.09正常细胞系 0.75±0.08乳腺癌细胞系0.24±0.05F值22.369P值0.001
图1 qRT-PCR分析MEG3在乳腺癌组织和细胞系中的表达量
2.2 MEG3的过表达抑制NK细胞与DC细胞的活力 检测lncRNA(MEG3)对NK细胞与树突状细胞(DC)免疫功能的影响,将MEG3转染到乳腺癌细胞中来过表达MEG3,然后进行CCK-8测定。MEG3的过表达与对照转染细胞相比,抑制NK细胞与DC细胞的活力(P<0.05)。见表3。
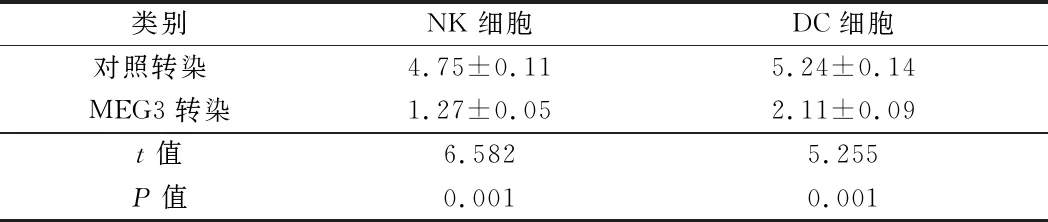
类别NK细胞DC细胞对照转染 4.75±0.115.24±0.14MEG3转染1.27±0.052.11±0.09t值6.5825.255P值0.0010.001
2.3 过表达的MEG3抑制乳腺癌细胞侵袭力 为研究lncRNA在乳腺癌细胞的肿瘤发生中可能的生物学功能,用MEG3转染乳腺癌细胞以特异性地上调MEG3的表达。Transwell测定法测量迁移和侵袭能力。与对照转染组相比,乳腺癌细胞中过表达的MEG3抑制乳腺癌细胞迁移和侵袭能力。表明过表达的MEG3可抑制乳腺癌细胞的侵袭能力。见表4,图2。
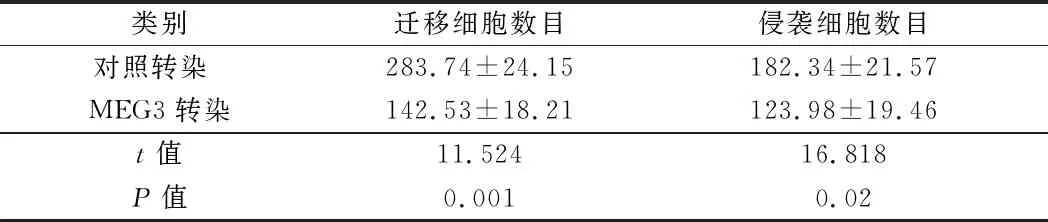
类别迁移细胞数目侵袭细胞数目对照转染 283.74±24.15182.34±21.57MEG3转染142.53±18.21123.98±19.46t值11.52416.818P值0.0010.02
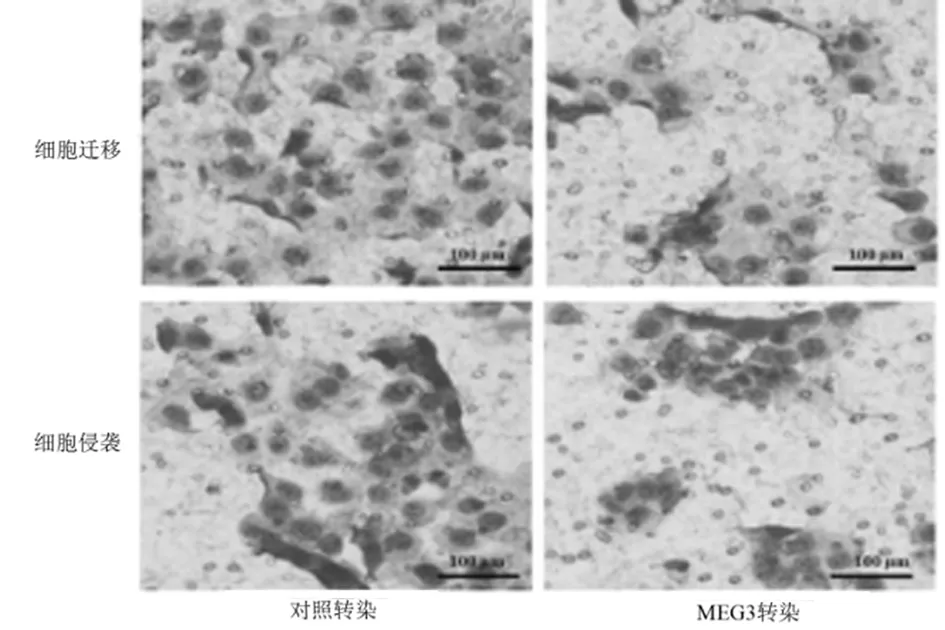
图2 不同转染后细胞(Wright’s-Giemsa染色×200)
2.4 miRNA是lncRNA的靶标 为探索lncRNA在乳腺癌中的分子机制,通过生物信息学分析预测了推定的MEG3靶标。结果显示在MEG3 mRNA中存在miRNA(miR-21)的结合序列(TAAGC)。为检测miR-21是否是lncRNA的功能靶标,通过miR-21模拟物或对照转染MEG3野生型(WT)和突变体(mut)细胞,检测荧光素酶报告基因,结果显示,与对照组相比,过表达的miR-21导致MEG3-WT荧光素酶活性降低,但对MEG3-mut报告基因的荧光素酶活性无明显影响,表明miR-21和MEG3之间的直接靶向关系。见表5。
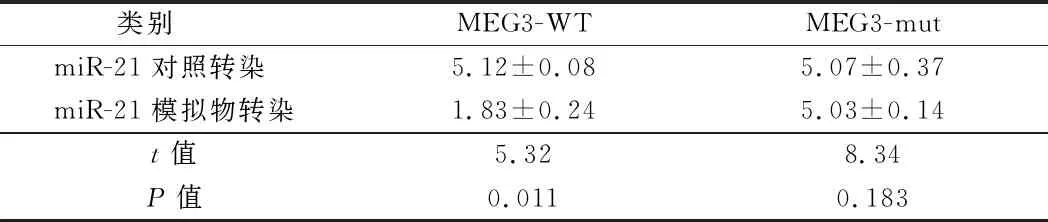
类别 MEG3-WTMEG3-mutmiR-21对照转染 5.12±0.085.07±0.37miR-21模拟物转染1.83±0.245.03±0.14t值5.328.34P值0.0110.183
2.5 过表达的MEG3减弱了miR-21模拟物对免疫侵袭相关蛋白的表达 为探索lncRNA和miRNA之间的潜在调节机制,进一步检测了两者对NK细胞和树突状细胞(DC)免疫功能及乳腺癌细胞侵袭力的影响。通过蛋白质印迹检测NK细胞免疫功能标记蛋白(CD56)、DC免疫功能标记蛋白(CD80)和细胞侵袭标记蛋白(MMP-2和MMP-9)的表达,结果显示,与miR-21对照相比,MEG3转染后CD56、CD80、MMP-2和MMP-9的表达量下降(P<0.05),而miR-21模拟物转染诱导能够上调CD56、CD80、MMP-2和MMP-9的表达(P<0.05);与miR-21模拟物转染组相比,MEG3的过表达导致CD56、CD80、MMP-2和MMP-9的表达量下降(P<0.05)。表明过表达的MEG3抵消了miR-21模拟物对NK细胞和DC免疫功能及乳腺癌细胞侵袭的促进作用。见表6,图3。
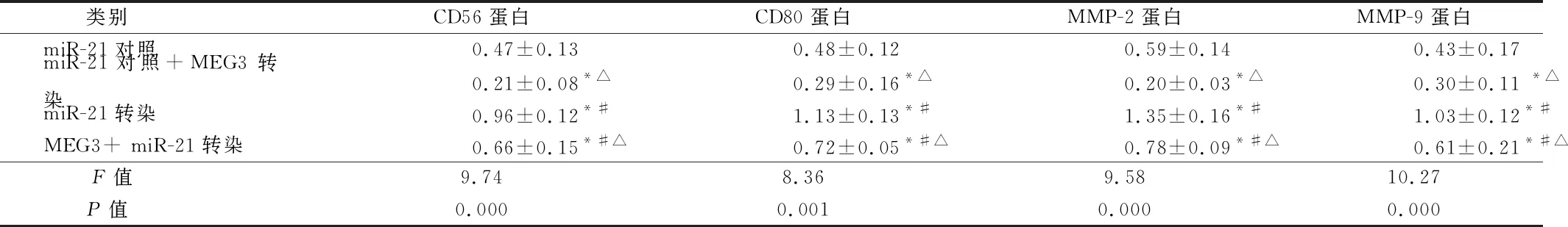
类别CD56蛋白CD80蛋白MMP-2蛋白MMP-9蛋白miR-21对照0.47±0.130.48±0.120.59±0.140.43±0.17miR-21对照+MEG3转染0.21±0.08*△0.29±0.16*△0.20±0.03*△0.30±0.11 *△miR-21转染0.96±0.12*#1.13±0.13*#1.35±0.16*#1.03±0.12*#MEG3+ miR-21转染0.66±0.15*#△0.72±0.05*#△0.78±0.09*#△0.61±0.21*#△ F值9.748.369.5810.27 P值0.0000.0010.0000.000
注:与miR-21对照比较,*P<0.05;与miR-21对照+MEG3转染比较,#P<0.05;与miR-21转染比较,△P<0.05

图3 不同转染后相关蛋白的表达水平
3 讨论
尽管已经实现了临床治疗的改进以改善临床状况,但具有晚期和远处转移的乳腺癌患者仍然具有非常差的预后,使得迫切需要发现针对NK细胞与DC免疫功能和乳腺癌细胞增殖和侵袭的新靶标。最近,已经鉴定出大量的LncRNA参与肿瘤进展并获得了很多关注[9]。越来越多的证据支持LncRNA MEG3在各种类型的肿瘤中充当肿瘤抑制因子[10-12]。例如,在子宫内膜癌组织中发现MEG3表达降低,过表达的MEG3通过激活PI3K途径抑制子宫内膜癌细胞增殖,侵袭,转移和促进细胞凋亡[10]。 MEG3在乳腺癌组织和细胞系中也下调。过表达MEG3可抑制乳腺癌细胞的增殖和侵袭[11]。与邻近正常组织相比,乳腺癌组织中MEG3水平降低,而下调MEG3与乳腺癌预后不良相关[12]。在我们的研究中,在乳腺癌组织和细胞系中观察到MEG3的抑制表达。表明MEG3可能在乳腺癌中起到肿瘤抑制剂的作用。为了验证我们的假设,在乳腺癌细胞系中MEG3转染以增加MEG3的表达,我们发现过表达的MEG3抑制NK细胞与DC活性以及乳腺癌细胞的增殖和侵袭,这表明过表达的MEG3抑制了乳腺癌的肿瘤进展。
一些LncRNA可以作为miRNA的海绵或诱饵。一些LncRNA也可以与miRNA竞争结合mRNA或产生miRNA。上调的MEG3通过海绵状癌中靶向E-cadherin的海绵状miR-421抑制细胞上皮 - 间质转化,MEG3的过表达通过miR-664的负调节抑制肝细胞癌细胞的增殖[13]。据报道,MEG3通过作为miR-19a的竞争性内源性RNA来抑制胶质瘤细胞的增殖,迁移和侵袭[7]。在我们的研究中,生物信息学分析的结果表明miR-21可能是MEG3的潜在靶标。已证明MiR-21在癌发生中起促进作用。miR-21表达在结直肠癌组织中上调,miR-21通过抑制肿瘤抑制因子PTEN促进细胞生长和结肠直肠癌细胞的侵袭。miR-21的减少可以通过PTEN/PDCD4诱导细胞凋亡并抑制SK-N-SH细胞的增殖,表明miR-21可能在神经母细胞瘤发育过程中起着致癌作用[5-12]。本研究结果显示,miR-21的过表达被MEG3抑制。此外,荧光素酶报告基因测定进一步证明了MEG3和miR-21之间的相互结合。因此,我们假设MEG3可通过抑制miR-21表达来抑制NK细胞与DC活性以及乳腺癌细胞的增殖和侵袭。
为探索MEG3和miR-21之间相互作用的确切分子机制,还将miR-21模拟物或MEG3转染到乳腺癌细胞中。我们的结果显示,与miR-21对照相比,MEG3转染后NK细胞免疫功能标记蛋白(CD56)、DC免疫功能标记蛋白(CD80)和细胞侵袭标记蛋白(MMP-2和MMP-9)的表达下降,而miR-21模拟物转染诱导能够上调CD56、CD80、MMP-2和MMP-9的表达。总之,这些结果支持了我们的假设,即MEG3通过抑制乳腺癌细胞中的miR-21表达来抑制NK细胞与DC活性以及乳腺癌细胞的增殖和侵袭。
综上所述,乳腺癌组织和细胞系中MEG3的表达降低。 过表达的MEG3通过抑制miR-21表达在体外和体内抑制NK细胞与DC活性以及乳腺癌细胞的增殖和侵袭。 MEG3和miR-21在乳腺癌进展中的调节作用,并为乳腺癌治疗提供了新的潜在治疗策略。
