利用免疫共沉淀联合质谱技术筛选盐藻MAPK的互作蛋白
2020-06-30岳金荣丛玉婷邢震宇高相楠王明芳柴晓杰
岳金荣 丛玉婷 邢震宇 高相楠 王明芳 柴晓杰
(大连海洋大学/辽宁省省级高校水生生物学重点实验室, 辽宁 大连 116023)
杜氏盐藻(Dunaliella salina),简称盐藻,属于绿藻门(Chlorophyta) 绿藻纲(Chlorophyceae) 团藻目(Volvocales) 多 鞭 藻 科(Polyblepharidaceae) 盐 藻 属(Dunaliella),广泛分布于海洋、盐田及盐湖中。 盐藻细胞形体微小,形状多样,两条等长的鞭毛位于细胞前端,可自由游动,无细胞壁[1]。 盐藻是已知最耐盐的单细胞真核生物,能够在0.05 ~5.00 mol·L-1NaCl 的培养液中生长、繁殖,是研究植物抗盐生理及其分子机制最理想的模式生物[2]。 因此,深入探讨盐藻适应高盐环境的分子机制具有重要的科学意义和应用价值。研究表明,在高盐胁迫下,盐藻通过调节细胞体积[3]、离子转运[4-6]以及大量合成甘油[7-9],以适应外界环境渗透压的变化。 但有关其基因表达调控机制尚未明确。
大多数真核生物中都存在有丝分裂原活化蛋白激酶(MAPKs)信号转导途径,它由三级激酶级联组成[10],即丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)、MAPK 激酶和MAPKK 激酶,这三类激酶都属于丝氨酸/苏氨酸(Ser/Thr)蛋白激酶。 植物细胞受到外界刺激时,先对Ser/Thr 进行磷酸化修饰,使MAPKKK-MAPKK-MAPK 相继激活,将上游信号进行传递,最终传递至下游。 细胞生长、发育、分裂和分化,以及细胞死亡等多个生理过程,都有MAPK 的参与,MAPK 激活后通过磷酸化转录因子或者多种蛋白(酶)等底物参与上述介导过程。 MAPK 广泛参与植物对各种逆境胁迫的应答过程,如干旱[11-13]、温度[14-15]及盐胁迫[16-18]等均有报道,但在水生植物藻类中的研究鲜有报道。 张婷等[19]应用实时荧光定量PCR 技术证实,DsMAPK的表达受盐胁迫诱导,当3 mol·L-1NaCl 胁迫处理0.5 h 时,DsMAPK的表达量显著上调(P<0.05);胁迫处理1 h 时,DsMAPK的表达量是正常对照组的5 倍,达到最大值,差异达极显著水平(P<0.01)。 Lei 等[20]也证明杜氏盐藻MAPK基因对高渗胁迫有响应。 Zhao 等[21]认为杜氏藻MAPK 基因(DtMAPK)调节3-磷酸甘油脱氢酶基因(GPDH)的表达及甘油的合成,说明MAPK在杜氏藻应答高盐胁迫的渗透调节中发挥着重要作用。 因此,进一步研究盐藻MAPK基因的功能及其互作蛋白,对于阐明盐藻耐高盐的分子机制具有重要的科学意义。
前期研究采用反转录PCR(reverse transcription-PCR,RT-PCR)和RACE(rapid-amplification of cDNA ends)技术成功从盐藻中克隆了促有丝分裂活化蛋白激酶基因(GenBank 登录号: JQ782412), 命名为DsMAPK,并成功构建了DsMAPK的原核表达载体[19,22]。 本研究应用基因工程技术使盐藻MAPK基因在大肠杆菌中成功表达,表达产物经His 柱纯化获得了高纯度的融合蛋白,以该融合蛋白作为抗原免疫动物,制备了多克隆抗体,然后采用免疫共沉淀技术、质谱技术 ( liquid chromatography-tandem mass spectrometry,LC-MS/MS)及生物信息学的方法筛选盐藻MAPK 互作蛋白,在蛋白质组学的水平上研究DsMAPK的功能,以期为进一步阐明DsMAPK在盐藻应答盐胁迫信号分子途径中的作用提供新的理论依据。
1 材料与方法
1.1 试验材料
杜氏盐藻(Dunaliella salina)由大连海洋大学水生生物学实验室提供,其培养液为甲液与乙液,比例为1 000 ∶1[23], 25℃,1 250 lx 光强,12 h 光照12 h 黑暗交替静置培养。
菌种:E.coliBL21 (DE3) 购自北京TIANGEN 公司。
载体质粒: 空载体pGS21a 质粒、重组质粒pGS21a-DsMAPK均由大连海洋学院/辽宁省省级高校水生生物学重点试验室保存。
1.2 主要试剂
异丙基硫代半乳糖苷(isopropyl thiogalactosine,IPTG)、His60 Ni SuperflowTMResin,购自美国Clontech公司;硝酸纤维素膜(NC 膜)、Anti-His Antibody、辣根过氧化物酶标记的羊抗鼠IgG、辣根过氧化物酶羊抗兔IgG HRP、蛋白分子量标准Mancer Protein Marker Mid Range,购自生工生物工程(上海) 有限公司。 其他化学试剂均为国产分析纯。
1.3 融合蛋白的诱导表达、纯化及复性
将空载体pGS-21a 质粒(对照组)和重组质粒pGS-21a (+)-Ds MAPK(试验组)分别转化E.coliBL21(DE3), 37℃培养过夜,挑单菌落,接种于LB 液体培养基[含50 mg·L-1氨苄青霉素(ampicillin,Amp)]中,37℃条件下190 r·min-1振荡培养至OD600介于0.6 ~0.8 之间,加入IPTG 诱导6 h,收集菌体[23]。 用PBS缓冲溶液重悬菌体并在冰上进行超声波破碎(功率30 W,工作5 s,间歇5 s)[23]。 12 000 r·min-1离心20 min后用尿素缓冲溶液(尿素、100 mmol·L-1磷酸二氢钠和10 mmol·L-1Tris-Cl,pH 值8.0)复溶沉淀,并将其置于4℃过夜。 2 mol·L-1尿素缓冲液冲洗沉淀之后再用8 mol·L-1尿素缓冲液重悬,过滤(0.45 μm 滤膜)后收集上清液,即为变性的重组蛋白粗品。
用His60 Ni SuperflowTMResin 纯化变性的重组蛋白粗品。 Ni-纯化柱经过平衡缓冲液(8 mol·L-1尿素缓冲液)平衡后,重组蛋白粗品上柱,用平衡液洗3 ~5次去除杂蛋白,最后由8 mol·L-1尿素缓冲液配置的洗脱液(2、30、40、50、200、500 mmol·L-1咪唑,pH 值8.0)洗脱3 次并收集。 取所有洗脱液各10 μL 分别加入等量的2×SDS 上样缓冲液,进行SDS-PAGE 分析。
将纯化后的融合蛋白洗脱液用尿素进行复性。 将蛋白洗脱溶液转移至透析袋,放置于尿素缓冲液[依次为6、4、2、0 mo1·L-1(pH 值8.0)]中,4℃保持12 h,随后在相同条件下依次更换低度浓度尿素缓冲液,最后转移到不含尿素的缓冲液中。
1.4 融合蛋白Western blotting 检测
纯化后的融合蛋白经SDS-PAGE 电泳,电转移至硝酸纤维素膜(200 mA,2 h),5%牛血清白蛋白封闭,以Anti-His Antibody 为一抗(1 ∶2 000),37℃孵育1 h,PBST 洗涤3 次,每次5 min,然后加入辣根过氧化物酶标记的羊抗鼠IgG(1 ∶200) 二抗,37℃孵育1 h,PBST洗涤,每次5 min,然后加入沉淀型单组分3,3,5,5-四甲基联苯胺(TMB)底物溶液显色。
1.5 多克隆抗体的制备及纯化
将纯化后的DsMAPK 样品与等体积的弗氏完全佐剂混合均匀,皮下注射免疫4 月龄健康雌性新西兰大白兔,初次免疫20 d,然后使用DsMAPK 与弗氏不完全佐剂混合物进行第2 次免疫,二次免疫14 d 后进行第3 次免疫,三免7 d 后进行耳静脉采血1 mL,间接ELISA 检测抗血清效价,未达到要求的进行第4 次免疫,四免8 d 后间接ELISA 检测抗血清效价达到要求,颈动脉采全血。 初免剂量为0.4 mg/只,二、三、四次免疫剂量为0.2 mg/只。
采用纯化介质Protein A SepharoseTMCL-4B 纯化抗血清,将纯化后的抗血清稀释至1 mg·mL-1后置于-80℃保存。
1.6 盐藻总蛋白的提取及Western blot 检测
取处于对数生长期的 NaCl 盐藻(含 1.0 mol·L-1), 加入NaCl 使其终浓度达到3 mol·L-1,置于恒温光照培养箱培养(25℃、光强1 250 lx),分别于培养1、3、6、12、24 h 取培养液。 培养液经8 000×g离心5 min 收集盐藻细胞。 用Sangon Biotech RIPA 裂解液Ⅳ提取总蛋白。 取10 μL 少量蛋白进行电泳检测,其余保存备用。
取20 μg 盐藻上清蛋白质,加入上样缓冲液后进行SDS-PAGE 电泳,电转至NC 膜,封闭后与多克隆抗体共孵育(1 ∶1 000 稀释),二抗采用HRP 标记的羊抗兔IgG(1 ∶2 000 稀释),进行TMB 底物溶液显色,拍照记录。
1.7 免疫共沉淀联合质谱技术筛选相互作用蛋白
取5.978 mg 对照组盐藻总蛋白,加入60 μg 普通兔的IgG;取5.978 mg 试验组盐藻总蛋白,加入60 μg DsMAPK 多克隆抗体,各自混合均匀,4℃孵育过夜。向盐藻总蛋白与抗体混合物中加入1/20 体积的蛋白A/G 琼脂糖珠并充分混匀,4℃孵育2 ~4 h。 1 000×g离心2 min,保留沉淀。 沉淀中加入1 mL PBS,1 000×g离心洗涤3 min。 重复上述离心操作3 次,然后向沉淀中加入50 μL 2×loading buffer,静置1 h 后沸水浴10 min,再经8 000×g离心5 min,保留上清液。 吸取20 μL 上清液进行SDS-PAGE 电泳分析。
1.8 LC-MS/MS 质谱分析
对1.7 中获得的上清液进行超滤、还原烷基化、酶解处理。 将冻干的肽段重溶于0.1%甲酸(formic acid, FA)中,进行在线液质联用分析。 色谱系统为nano ACQUITY UPLC(Waters Corporation,英国),与色谱相连的质谱为Q Exactive hybrid quadrupole-Orbitrap mass spectrometer (Thermo Fisher Scientific,美国)。 样品先以10 μL·min-1的流速将其结合在trap 柱上(Thermo Scientific Acclaim PepMap C18 column,100 μm×2 cm, 3 μm particle size),然后再于分析柱(Acclaim PepMap C18,75 μm×15 cm) 上进行分离。梯度分离条件:在40 min 内将乙腈浓度由5%升至45%。 质谱条件:纳升级电喷雾电离方式(nano-ESI),正离子模式扫描,扫描范围m/z 350 ~1 500。 采用PEAKS STUDIO 软件(Bioinfor Inc,美国) 进行数据处理,数据库为大连海洋学院/辽宁省省级高校水生生物学重点试验室提供的转录组数据库。
2 结果与分析
2.1 融合蛋白的诱导表达、纯化及Western blotting检测
由图1-A 可知,对照组表达菌(含空载体pGS-21a)经诱导表达后在35 kDa 左右有一条表达增强的标签蛋白条带;2、3 号泳道为试验组表达菌(含重组质粒pGS-21a-Ds MAPK),经IPTG 诱导表达后的重组菌体经超声波破碎得到的上清液和沉淀,2 号泳道未见清晰的目的条带,3 号泳道在87 kDa 左右有一条表达增强的目的蛋白条带,此条带在对照组未出现,与预期结果相符,即融合蛋白包括预计分子量为52 kDa 的DsMAPK 融合蛋白和35 kDa 的His 标签蛋白。 此外,上清液中未见清晰的目的蛋白条带,表明目的蛋白主要是以包涵体的形式表达。

图1 融合蛋白的诱导表达、纯化及Western blotting 检测Fig.1 Expression, purfication and Western bloting analysis of induced recombinant protein
对融合蛋白进行纯化,纯化产物经SDS-PAGE电泳检测,纯化产物在87 kDa 左右有一条蛋白带,大小与预期相符(图1-B、C)。 将其中纯化条带单一的蛋白样品收集。 为了进一步确定纯化蛋白是否为目的蛋白,对其进行了Western blotting 检测(图1-D),可见1 号泳道在87 kDa 左右有一清晰的杂交信号带,初步证明纯化的蛋白就是带有His 标签的DsMAPK 融合蛋白。
2.2 盐藻总蛋白的提取及Western blotting 检测
由图2-A 可知,各处理组均出现多个清晰的条带,表明盐藻总蛋白的提取效果良好。 Western blotting 检测结果表明(图2-B),对照组和盐胁迫试验组均在50 kDa 左右出现单一条带,与DsMAPK 蛋白大小相符,初步判定DsMAPK 抗体特异性良好且在盐胁迫3 h 和6 h 时盐藻总蛋白的表达量较高。
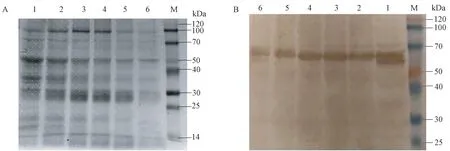
图2 盐藻总蛋白提取及Western blotting 检测Fig.2 Extraction of total protein of Dunaliella salina and detection by Western blotting
2.3 免疫共沉淀筛选DsMAPK 的相互作用蛋白
由3-A 可知,1、2 号泳道均为多个且清晰的条带,表明盐藻总蛋白的提取效果良好。 免疫共沉淀产物的SDS-PAGE 分析结果显示,2 号泳道在约28 kDa 处有明显的差异条带,说明用免疫共沉淀方法成功筛选出DsMAPK 相互作用蛋白。
2.4 LC-MS/MS 及生物信息学分析
盐藻总蛋白免疫共沉淀后经脱色、还原、烷基化、酶解、萃取等处理,将制备好的样品用Q Exactive hybrid quadrupole-Orbitrap 质谱仪进行质谱分析,然后将质谱鉴定结果导入PEAKS STUDIO 软件(Bioinfor Inc.,美国) 搜索盐藻转录组数据库。 筛选肽段可信度高(peptide confidence-high) 的肽段,并由软件根据同一特异肽段(unique peptide)在各样本中的报告离子峰的面积比值计算蛋白质的表达量。 如果某个蛋白的差异倍数(fold change)>2.0,同时统计检验P<0.05,便可认为是差异蛋白。 由图4 可知,试验组鉴定出的特有蛋白165 种。 为了更深入地了解高盐胁迫下DsMAPK 的生物学功能,对筛选的165 差异蛋白进行基因本体GO 功能富集分析和KEGG Pathway 分析。
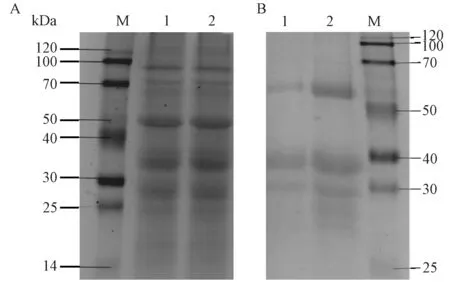
图3 盐藻总蛋白提取及免疫共沉淀结果Fig.3 Total protein of Dunaliella salina and co-immunoprecipitation products identify by SDS-PAGE electrophoresis

图4 盐胁迫下盐藻差异表达蛋白的分布Fig.4 The distribution of differentially expressed proteins of Dunaliella salina under salt stress
GO 注释主要是对蛋白质的分子功能(molecular function)、细胞组分(cellular component)、生物进程(biological process) 进行分析。 图5 是一个GO 富集分析汇总柱状图,展示了MF、CC 和BP 3 种类别富集分析显著性(P<0.05)排名前10 的条目。 生物进程分析表明,差异蛋白主要参与细胞凋亡、小分子生物合成过程以及细胞形态结构的调节;细胞组分分析表明,差异蛋白主要位于细胞质、核糖体;分子功能显示,差异蛋白主要参与了生物小分子、核苷酸、ATP 的结合。可见,盐藻响应盐胁迫的差异表达蛋白具有多种分子功能,并参与多个生物学过程,表明盐藻盐胁迫响应是一个复杂的生理生化过程。
KEGG 是一个利用图形介绍众多的代谢途径及各途径之间关系的数据库。 图6 根据KEGG 富集汇总表中选择的调控通路,展示了显著性排名前10 的条目,条柱均高于蓝线或红线,其所代表的调控通路具有显著性。 KEGG 代谢通路分析表明,盐藻差异蛋白主要参与了新陈代谢、遗传信息的传递,外界环境信息的应答,细胞周期调控,以及其他未知的途径等,表明信号转导相关蛋白、胁迫相关蛋白以及代谢相关蛋白在盐藻应答高浓度盐胁迫过程中发挥了重要作用,为全面了解盐藻的抗盐机制奠定了基础。
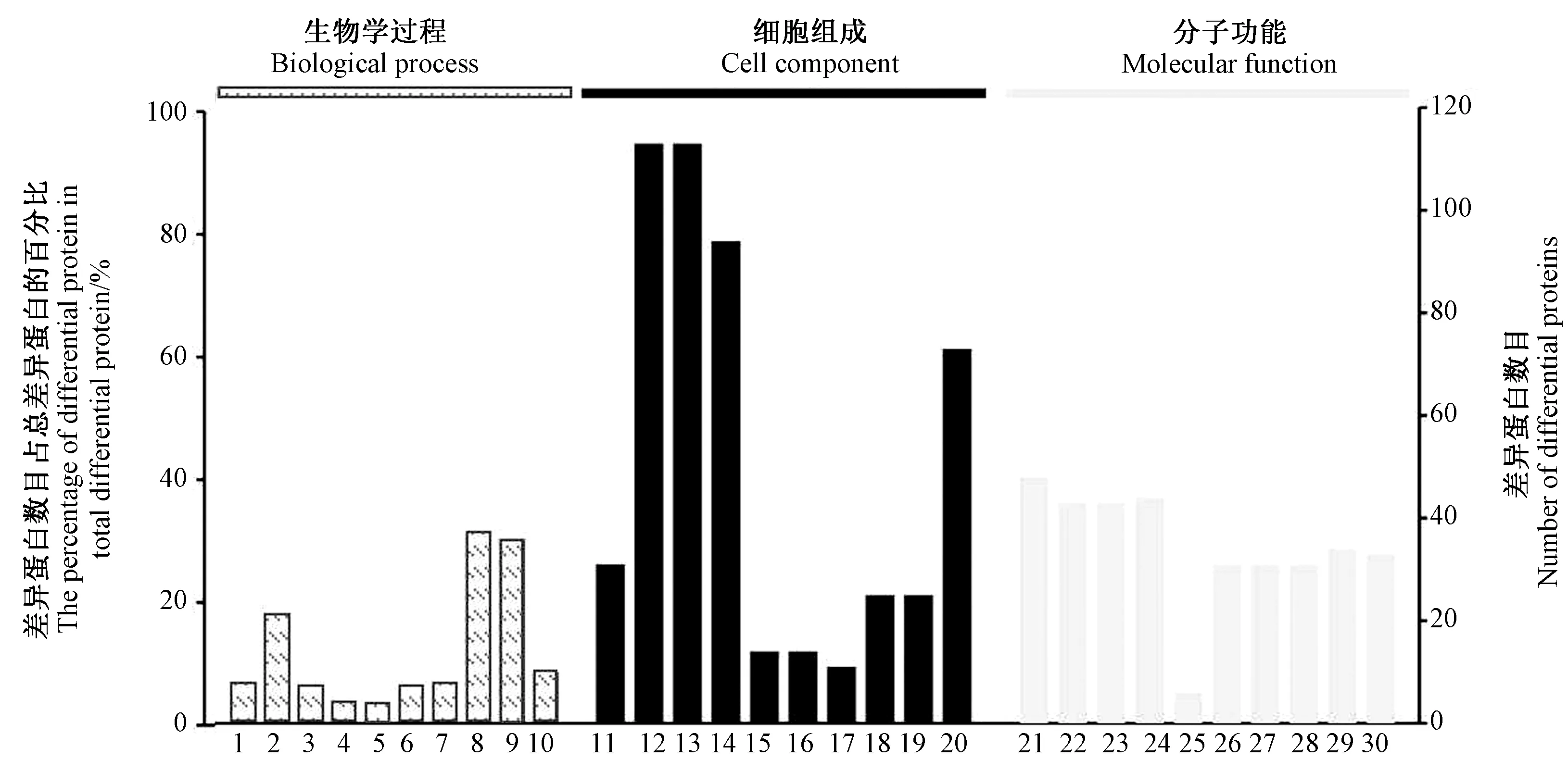
图5 盐藻差异表达蛋白的GO 功能及富集分析结果Fig.5 GO function and enrichment analysis results of differentially expressed proteins of Dunaliella salina
基于string 数据库和cytoscape 软件,制作了蛋白互作网络图(图7),表示了KEGG 富集结果中显著性较高的前10 个通路以及与之互作蛋白之间的互作网络,该网络图中有32 种蛋白。 互作图是以圆点(基因或蛋白)、多边形(代谢产物)、三角形(miRNA)、五角形(转录因子)与圆角矩形(生物过程、细胞定位、分子功能或信号通路)以不同直线(互作关系虚线为未经试验验证,实线为已有相关验证报道)进行连接展示的分子相互作用机制与网络模型图。 由图7 可知,PGK1(3-磷酸甘油酸激酶)、LAS17(肌动蛋白结合蛋白)、BCK1(丝裂原活化蛋白激酶激酶激酶,即为MAKKK)与DsMAPK 互作蛋白得分均在840 以上,URA2 与DsMAPK 互作得分为626,互作的可能性较高。 但PGK1、LAS17、BCK1、URA2 与DsMAPK 是否互作,尚未验证,后续将通过酵母双杂交的方法鉴定以上蛋白质之间是否存在互作关系。
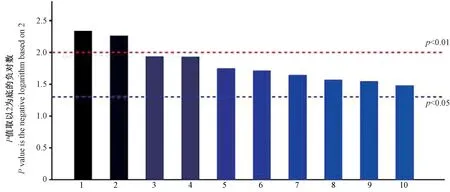
图6 盐藻差异蛋白的KEGG 分析Fig.6 KEGG pathway analysis of differentially expressed proteins of Dunaliella salina
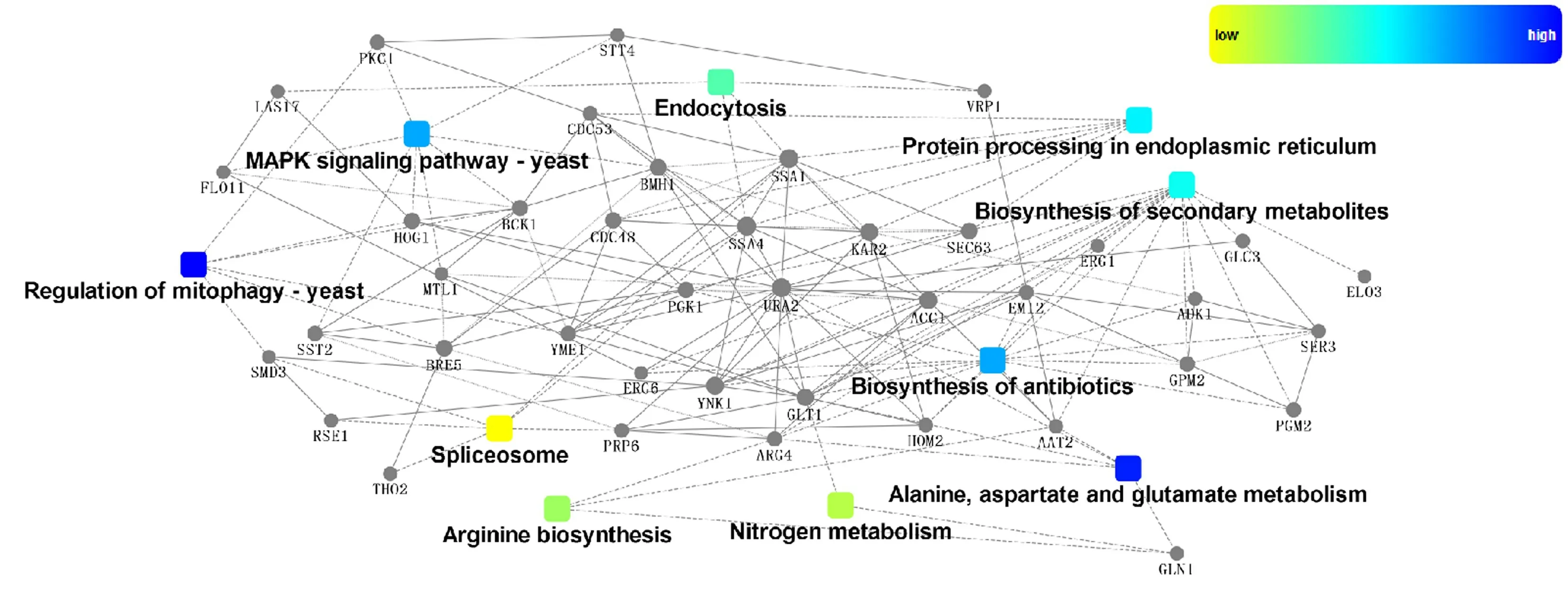
图7 蛋白质相互作用的网络图Fig.7 Network diagram of protein interactions
3 讨论
盐胁迫反应和应答是一个极其复杂的过程,细胞内的蛋白质会组成一个复杂的相互作用网络,构成各项生理活动的基础。 因此,研究蛋白质之间的相互作用关系,对于阐明细胞高度复杂的分子机制具有重要的科学意义。 免疫共沉淀是研究蛋白质相互作用的有效方法,技术成熟,可信度高。 该技术是利用抗原和抗体特异性结合以及免疫蛋白Fc 片段与蛋白A/G 琼脂糖珠的结合,形成“结合蛋白-靶蛋白-靶蛋白抗体-蛋白A/G 琼脂糖珠”复合物,从而获得与诱饵蛋白相互作用的目的蛋白[24]。 本研究采用免疫共沉淀方法筛选出与DsMAPK 相互作用的蛋白质,并结合质谱技术和生物信息学分析方法,共检测到165 种特有差异蛋白。 通过GO 和KEGG 分析发现,这些差异蛋白主要参与了新陈代谢、遗传信息的传递以及信号转导等生物学调控过程。
甘油是盐藻响应盐胁迫最主要的渗透调节物质,盐藻通过大量合成甘油,以适应外界环境渗透压的变化[2,7]。 研究表明,在高盐胁迫初期,光合作用受到抑制,合成甘油所需的磷酸二羟丙酮(dihydroxyacetone phosphate,DHAP)主要来源于淀粉代谢,而非光合作用[25-26]。 随着盐胁迫时间的延长,细胞内储存的淀粉被消耗,盐藻细胞的调控机制逐渐恢复,光合作用逐渐增强[27]。 本研究鉴定到多种参与光合作用的蛋白质,分别是光系统Ⅰ反应中心亚基Ⅱ、铁氧还蛋白、1,5-二磷酸核酮糖羧化酶/加氧酶(RuBisCO)、磷酸烯醇式丙酮酸羧化酶、3-磷酸酸甘油酸激酶(PGK1)及3-磷酸甘油醛脱氢酶。 光系统Ⅰ反应中心亚基Ⅱ和铁氧还蛋白是光合电子传递链的组分,参与氧化还原反应;RuBisCO 是光合作用中的关键酶,催化羧化反应和氧合反应,小亚基rbcS 具有调控RuBisCO 活性的功能,在转基因研究中,RuBisCO 小亚基rbcS 基因的启动子经常作为高效表达的调控元件[28];磷酸烯醇式丙酮酸羧化酶催化磷酸烯醇式丙酮酸不可逆反应生成磷酸和草酰乙酸,是光合作用的关键酶,存在于所有光合生物中,同时还参与氨基酸代谢中碳骨架回补作用[29];磷酸甘油酸激酶催化3-磷酸甘油酸转变成1,3-二磷酸甘油酸,后者在脱氢酶作用下还原为3-磷酸甘油醛,3-磷酸甘油醛可以转变成DHAP。 DHAP 通过3-磷酸甘油脱氢酶转化为3-磷酸甘油,再通过磷酸酶去磷酸化而合成甘油[2,7]。 因此,推测DsMAPK 调控了盐藻在高盐胁迫下的光合作用及甘油合成等生物学过程。
前期研究发现,高盐胁迫下,盐藻会发生鞭毛脱落的现象[30],但其恢复机制未见报道,本研究发现DsMAPK 与S-腺苷高半胱氨酸水解酶存在互作关系。盐藻S-腺苷高半胱氨酸水解酶参与真核生物最主要的甲基化反应,参与盐藻鞭毛的再生,说明DsMAPK可能在盐藻细胞鞭毛的再生中起到重要的调控作用。此外,还发现了细胞分裂调控蛋白48、蛋白激酶C 及肌动蛋白。 Ravanti 等[31]认为MAPK 通过上调某些转录因子基因的表达和生物活性,影响细胞的增殖、分化和细胞因子的合成。 高盐胁迫下,盐藻细胞快速失水变小,平衡胞内外的渗透压,以适应外界的高渗环境[32]。 肌动蛋白组成微丝,普遍存在于真核细胞中,构成细胞的支架,维持细胞的形态。 因此,推断DsMAPK 可能参与了盐藻细胞鞭毛的再生、藻细胞的增殖及细胞形态的恢复。
本研究结果还有待于进一步验证,但这些重要蛋白质的发现丰富和发展了现有理论,为下一步的研究提供了新思路。 后续工作将通过酵母双杂交的方法进一步验定这些蛋白质的功能。
4 结论
本研究通过原核表达纯化的DsMAPK 蛋白成功制备出多克隆抗体且特异性良好,经过免疫共沉淀联合质谱技术以及生物信息学的方法筛选出165 种特有的差异蛋白。 通过GO 和KEGG 分析发现,这些差异蛋白主要参与了新陈代谢、遗传信息的传递以及信号转导等生物学调控过程。 蛋白质互作网络分析发现,直接与MAPK 相互作用的蛋白有4 种。 本研究结果为进一步研究杜氏盐藻响应盐胁迫的分子机制提供了材料。
