长湖浮游植物优势种季节演替及生态位分析
2020-06-13魏志兵罗静波谭凤霞杨德国何勇凤
魏志兵 柴 毅 罗静波 郭 坤 谭凤霞 杨德国 何勇凤
(1. 长江大学湿地生态与农业利用教育部工程研究中心, 荆州 434025; 2. 长江大学动物科学学院, 荆州 434025; 3. 中国水产科学研究院长江水产研究所, 农业部淡水生物多样性保护重点实验室, 武汉 430223)
生态位(Niche)是指物种在其生存环境中时间、空间分布和生物量占有等系列利用资源的能力, 受群落内生物和非生物环境的影响, 与种间竞争密切相关[1,2]。生态位宽度是指生物种群在群落中所利用不同资源的能力总和, 生态位越宽, 其竞争能力越强。生态位重叠体现了两个物种对群落内共同资源的利用程度, 也是研究种间竞争的一个重要因子[3]。目前生态位理论已被广泛应用于陆生生物如昆虫、花卉、森林等[4—7]和水生生物如鱼类、浮游生物和大型底栖动物[8—10]。其中浮游植物生态位的相关研究主要集中在内陆典型湖泊[11—13],可对湖泊富营养化状态进行综合评价并预测其发展趋势。近年来随着多元统计分析的广泛运用, 多种生态学参数如OMI指数(Outlying Mean Index, 边缘平均指数)和TOI指数(Tolerance Index, 耐受指数)等也被用于物种生态位与环境之间的相关性分析, 被认为是较好的生态位研究应用[14]。
长湖作为我国长江中下游和江汉平原地区重要的生态功能区, 具有防洪调蓄、供水灌溉、航运和旅游等多种重要功能, 2010年被农业部批建为鲌类国家级水产种资源保护区。研究发现, 由于人类活动如工业废水排放、农业面源污染、大面积围网围栏养殖等影响, 长湖处于富营养化状态[15—19]。而近年来引江济汉、蒙华铁路和318国道长湖特大桥等涉水工程的陆续建设也会对该水域产生不同程度的影响。本文运用多种生态学参数对长湖浮游植物优势种生态位特征进行分析评价, 并与本课题组2012年的研究报道[11]做对比, 以期对长湖浮游植物生态位进行深入和全面的分析, 揭示其演替规律并科学预测其发展趋势。为长湖水生生物资源的可持续发展提供重要参考, 旨在为下一步长湖生态系统的功能恢复、价值评价以及健康良好生态系统多样性格局的维护提供科学依据。
1 材料与方法
1.1 区域概况
长湖现有流域面积约3240 km, 平均水深2.1 m,年均水温为17.5℃, 常年光照充足, 其年降雨量在1200 mm左右[16]。如图1所示, 长湖自西向东分为庙湖、海子湖、马洪台和圆心湖4个区域。受“引江济汉”工程建设影响, 庙湖已被两条堤坝拦截形成静水, 故在此区域不设置采样点。海子湖为一狭长形湖湾, 分布有5个进出水口, 马洪台和圆心湖为开阔水域, 均有较大面积的围网围栏养殖。
1.2 采样点设置与采样时间
采样点的设置参考《水域生态系统观测规范》[20]进行并根据长湖实际水域形态、进出水口以及水文特征进行(表1), 共设置14个采样点, 其中海子湖区4个(H1—4), 马洪台区(M1—5)和圆心湖区(Y1—5)各5个, 每次取样均用GPS仪进行样点定位。采样时间分别为2015年4月(春季)、7月(夏季)、11月(秋季)和2016年1月(冬季)。
1.3 样本的采集与分析
使用多参数水质分析仪(HQ40D, Hach Loveland, USA)现场测定溶解氧(DO)、水温(WT)和pH,利用塞氏盘测定透明度(SD)。总磷(TP)、总氮(TN)、磷酸盐硝态氮亚硝态氮和氨氮等水化学指标按照国家水质标准方法GB 3828-2002进行测定。
浮游植物定性样本使用25#浮游生物网(网目65 μm)进行采集, 加4%甲醛固定后带回实验室用于种类鉴定。定量样本使用有机玻璃采水器(1 L)在表层至0.5 m深处和0.5—1.0 m深处分别采样, 样本充分混合后取1 L加入15 mL鲁哥氏液固定, 带回实验室后沉淀、浓缩至30—50 mL用以定量计数[21]。
使用显微镜(XSM-20, 宁波舜宇仪器有限公司,宁波, 浙江)对浮游植物样本进行种类鉴定和计数,浮游植物鉴定准则参考《中国淡水藻类-系统、分类及生态》, 且所有种类鉴定到种[22,23]。
1.4 数据分析
优势种采用Mcnaughton优势度指数(Y)来确定,Y>0.02时即视为优势种。生态位宽度(Bi)采用以Shannon-Wiener多样性指数为基础的生态位宽度公式计算。生态位重叠指数(Oik)采用Levins指数计算。各公式如下:


图1 长湖采样点示意图Fig. 1 The sampling sites in Changhu Lake
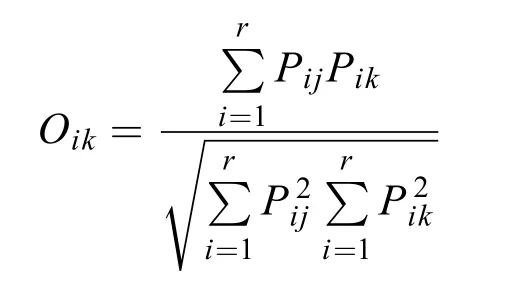
式中,ni为第i种在各采样点的总个体数,N为各个采样点中浮游植物的总个体数,fi为第i种在各个采样点出现的频率,nij为第i种在j采样点的个体数,Pij表示种i在第j个采样点的个体数占该物种所有个体数的比,r和R为采样点总数,Oik为种i与种k的资源利用曲线的生态位重叠指数,Pik为种i在j采样点的个体数占该种所有个体数的比值。
平均边缘指数(OMI)是通过设立其适合生境是所在调查区域平均生境的假定物种,再计算该物种与假定物种之间的距离。耐受指数(TOI)是通过计算物种调查区域物种沿环境梯度分布范围的宽窄。
丰度和优势度的相关计算和做图通过Excel 2013完成。其他生态位相关分析均通过R_V3.4.3的spaa和vegan程序包完成, 使用vegan包中的相关函数进行计算和制图, 同时进行显著性检验(以P表示)[24]。
2 结果
2.1 浮游植物优势种群落结构特征
浮游植物组成及优势种本次调查共鉴定出浮游植物7门67属120种, 主要包括绿藻门(61种,占比50.8%)、硅藻门(25种, 占比20.8%)和蓝藻门(18种, 占比15.0%), 其余裸藻门、黄藻门、金藻门和甲藻门共16种, 共占比13.4%。如表1所示, 优势种共14种, 隶属于5门14属, 其中扭曲小环藻(Cyclotella comta)、小球藻(Chlorella vulgaris)和梭形裸藻(Euglena acus)是4个季节的共有优势种。
以出现频率超过90%的优势种的季节分布来看, 春季是小球藻、镰形纤维藻和四尾栅藻; 夏季是湖泊色球藻(Chlorella limneticus)、类颤鱼腥藻(Osicellariordes)、微小平裂藻(Merismopedia tenuissima)、小球藻和四尾栅藻, 其中除四尾栅藻外其余4种的出现频率均为100%; 秋季是湖泊色球藻、四尾栅藻、肘状针杆藻(Synedra ulna)和梭形裸藻, 其中梭形裸藻为100%; 冬季是扭曲小环藻、肘状针杆藻、帽形菱形藻(Nitzschia cap)和梭形裸藻, 其中肘状针杆藻的为100%。
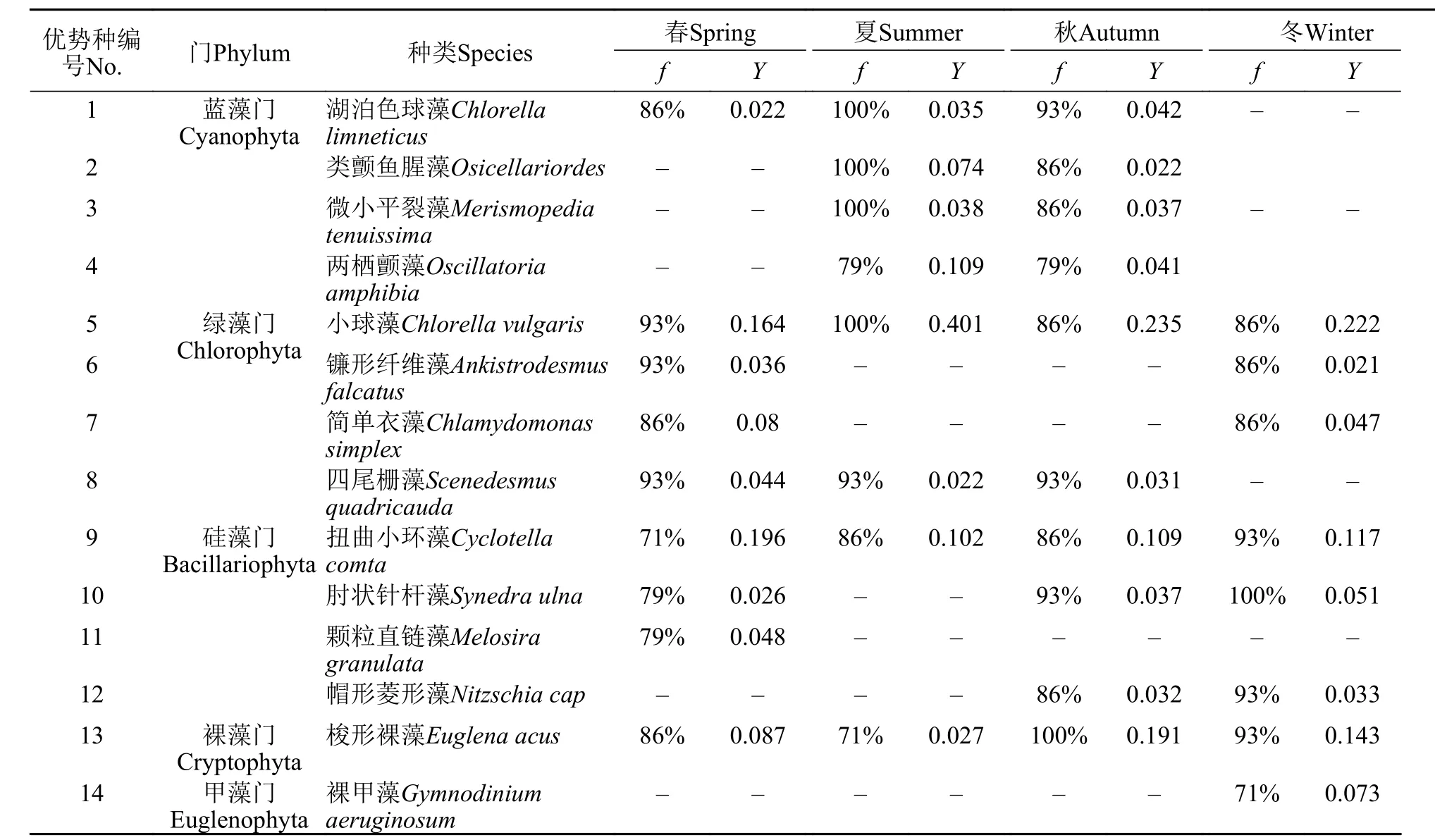
表1 长湖浮游植物优势种出现频率和优势度指数Tab. 1 Occurrence frequency and dominance index of dominant species phytoplankton in Changhu Lake
以各季节分布的优势度来看, 春季优势度指数最高的是扭曲小环藻(0.196), 而小球藻在夏、秋、冬3个季节的优势度指数均为最高, 分别为0.401、0.235和0.222。
优势种丰度如图2所示, 总体来看, 总丰度呈夏>春>秋>冬的变化趋势, 各藻门丰度在季节和采样点的分布上则表现出较明显的差异性。硅藻门在春季整体占比较高, 其中采样点H2(7.86×106cells/L)、H4(9.86×106cells/L)、M2(0.53×106cells/L)和M3(3.83×106cells/L)的占比均超过50%, 其中H4占比最高, 达79.9%。蓝藻门在夏季的占比最高, 其中采样点Y1(9.94×106cells/L)、Y2(11.50×106cells/L)、Y4(4.15×106cells/L)和Y5(9.73×106cells/L)的占比均超60%, 最高达80%(Y5)。绿藻门的整体占比均较高, 其中超过60%的有春M1(2.21×106cells/L)、M4(3.27×106cells/L)、M5(6.83×106cells/L)、Y1(1.60×106cells/L)、Y2(1.94×106cells/L)和夏H3(22.70×106cells/L)、M1(12.10×106cells/L)、M2(8.38×106cells/L)、M3(33.50×106cells/L)以及秋Y1(5.63×106cells/L)、Y2(3.60×106cells/L)和冬M4(2.46×106cells/L)共计12个采样点。其中春M5占比最高, 达83.7%。裸藻门在四季的整体占比均较低。
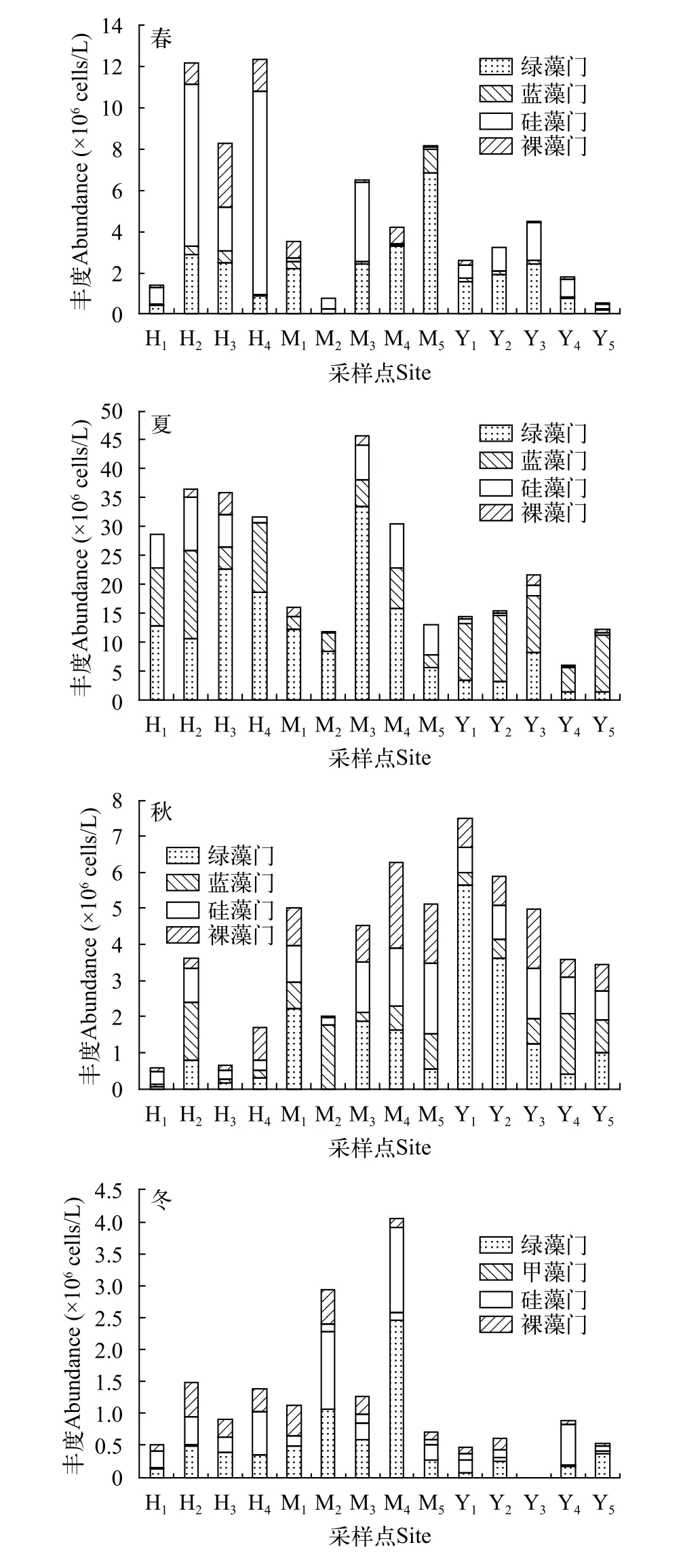
图2 长湖浮游植物优势类群丰度变化Fig. 2 Variation of abundance of phytoplankton dominant group in Lake Changhu
2.2 优势种生态位特征
对长湖浮游植物优势种生态位分析可知(图3),生态位宽度变化范围为1.002—2.377, 其中夏季小球藻最高, 其余大于2的种类分别是春季四尾栅藻、扭曲小环藻、颗粒直链藻, 夏季类颤鱼腥藻、两栖颤藻, 秋季类颤鱼腥藻、两栖颤藻、小球藻、帽形菱形藻以及冬季裸甲藻, 以上11种可以划归为广生态位种类。平均边缘指数(OMI)变化范围为0.25—3.71, 其中小于1的物种共12种。耐受指数变化范围为0.13—3.35, 夏季两栖颤藻最高, 其次是夏季类颤鱼腥藻(3.08)、冬季裸甲藻(2.67)和春季小球藻(2.41)。
通过对P值小于0.05的优势种进行OMI分析结果表明, 共有16种优势种的生态位分化较为显著。对优势种生态位和生态位宽度在第一轴和第二轴上分布情况的OMI分析表明(图4), 优势种和环境因子在各个季节的分化程度差异较明显。
2.3 优势种生态位重叠
优势种生态位重叠值的变化范围为0.02—0.93(图5), 春季肘状针杆藻和梭形裸藻以及秋季扭曲小环藻和梭形裸藻的重叠值均为最高值0.93。生态位重叠值超过(含)0.90的还有春季颗粒直链藻和梭形裸藻(0.91), 秋季湖泊色球藻和四尾栅藻(0.92)、类颤鱼腥藻和微小平裂藻(0.92)以及冬季小球藻和镰形纤维藻(0.90)、小球藻和扭曲小环藻(0.91)。生态位重叠值处于0.80(含)—0.90的共有19组优势种。
2.4 浮游植物优势种与环境因子的CCA分析
先将物种数据进行平方根转化再进行除趋势对应分析(DCA), pH以外的所有理化数据进行lg(x+1)转换, 各个季节4个排序轴的最大梯度皆处于3—4之间, 选用CCA单峰模型进行排序。选取溶氧(DO)、水温(WT)、酸碱度(pH)、透明度(SD)、总磷(TP)、总氮(TN)、氮磷比(N∶P)共7个环境因子与浮游植物优势种进行CCA排序(图6), 四个季节在轴一、轴二的特征值分别为0.632和0.143(春季)、0.591和0.286(夏季)、0.585和0.182(秋季)以及0.761和0.187(冬季), 其对应的解释度分别为73.6%和16.7%(春季)、74.2%和16.4%(夏季)、69.4%和21.7%(秋季)以及72.4%和17.8%(冬季), 再经蒙特卡罗检验(Monte Carlo test)第1轴和第2轴均P<0.05, 表示CCA排序结果可信。
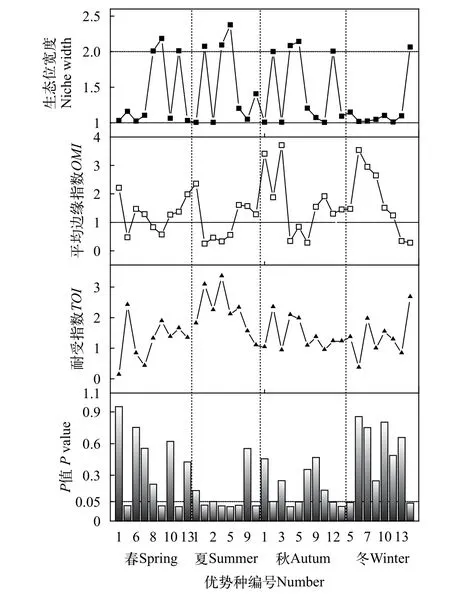
图3 浮游植物优势种生态位参数Fig. 3 Niche index of dominant species of phytoplankton
结合各环境因子与排序轴相关性分析可知, 春季小球藻与pH呈正相关, 镰形纤维藻与TN有较大的相关性, 湖泊色球藻、四尾栅藻、扭曲小环藻、颗粒直链藻和梭形裸藻与环境因子的相关性都很低。夏季微小平裂藻、两栖颤藻、四尾栅藻、扭曲小环藻以及梭形裸藻分布在排序轴的右下方, 表现出与TN和TP呈较强的正相关性, 与其他环境因子相关性较低。秋季湖泊色球藻、类颤鱼腥藻、微小平裂藻、小球藻、四尾栅藻、扭曲小环藻、肘状针杆藻、帽形菱形藻和梭形裸藻共计9种优势种表现出与pH和DO有正相关性。冬季肘状针杆藻与SD有较大正相关性, 梭形裸藻与pH有较大正相关性。
3 讨论
3.1 优势种群落结构特征
浮游植物的种类组成、丰度及分布等群落结构特征对水域环境具有较强的指示作用, 种类数量变化越小、丰度越高, 其富营养化程度就越高[25]。本次调查结果显示绿藻门占比最高(50.8%), 其次为硅藻门(20.8%)和蓝藻门(15.0%), 表明长湖属典型绿藻门为主的水域类型, 浮游植物种类组成单一,多样性较差, 群落结构不稳定。有研究发现[26]当水体受到有机污染时会导致金藻减少甚至消失, 而此次调查未发现金藻门种类, 表明除富营养化程度较高外, 长湖还存在受到有机污染的可能性。
本次调查共鉴定出优势种14种。其中出现频率为100%的种类有春季小球藻、镰形纤维藻和四尾栅藻以及夏季湖泊色球藻、类颤鱼腥藻、微小平裂藻和小球藻, 以上种类均为富营养化水体的常见种。从优势度来看, 夏、秋、冬3季优势度指数最高的种类均为小球藻, 表明多优势度较集中, 多样性较差。优势度较高的种类将会占据更多的生态位, 种间竞争作用降低, 从而导致群落结构稳定性较差, 水体中发生水华的风险将会增加[25]。
从浮游植物丰度的平面分布来看, 各采样点优势种的丰度均超过了106cells/L, 参照湖泊富营养化评价标准[27]表明长湖处于富营养化状态。从4个季节的总丰度来看, 呈现出海子湖、马洪台和圆心湖逐渐降低的分布特征, 这与各湖区的地理环境特点紧密相关。海子湖呈狭长型, 且随着农村城镇化发展, 沿岸带人为因素干扰不断增多, 此外与之相连多个进出水口也可增加该水域营养盐的水平, 以上诸多因素均可影响浮游植物丰度的累积。马洪台和圆心湖为开阔水域, 较大面积的围网围栏养殖会造成水体交换率低、营养盐水平升高等现象, 从而导致浮游植物丰度较高, 水体呈富营养化状态。
3.2 优势种生态位特征分析
生态位宽度是衡量生物利用各种资源的一个指标, 可反映出物种的分布和数量, 依据其大小可分为狭生态位和广生态位[28]。生态位越宽, 表示物种对环境中各种资源的利用能力越强, 其在环境中的竞争能力就越强, 即为广布种或是泛化种, 反之则为特化种。从本次调查结果来看, 生态位宽度大于2的种类共11种(表2), 如夏季小球藻(2.377)、两栖颤藻(2.092)和类颤鱼腥藻(2.075)等, 可划归为广生态位种类。而生态位宽度较小的种类主要如夏季湖泊色球藻(1.005)和微小平裂藻(1.004)等可划归为狭生态位种类。有研究[29]发现优势种生态位宽度侧重于物种和资源位点的数量, 物种与分布站点密切相关。这与本次调查结果表现出了较强的一致性, 广生态位的优势种丰度值较高, 分布站点也较多, 从而获得较高的生态位宽度, 说明对生境有较强的适应性, 这也与长湖夏季常出现区域性绿藻水华(主要是小球藻)一致。
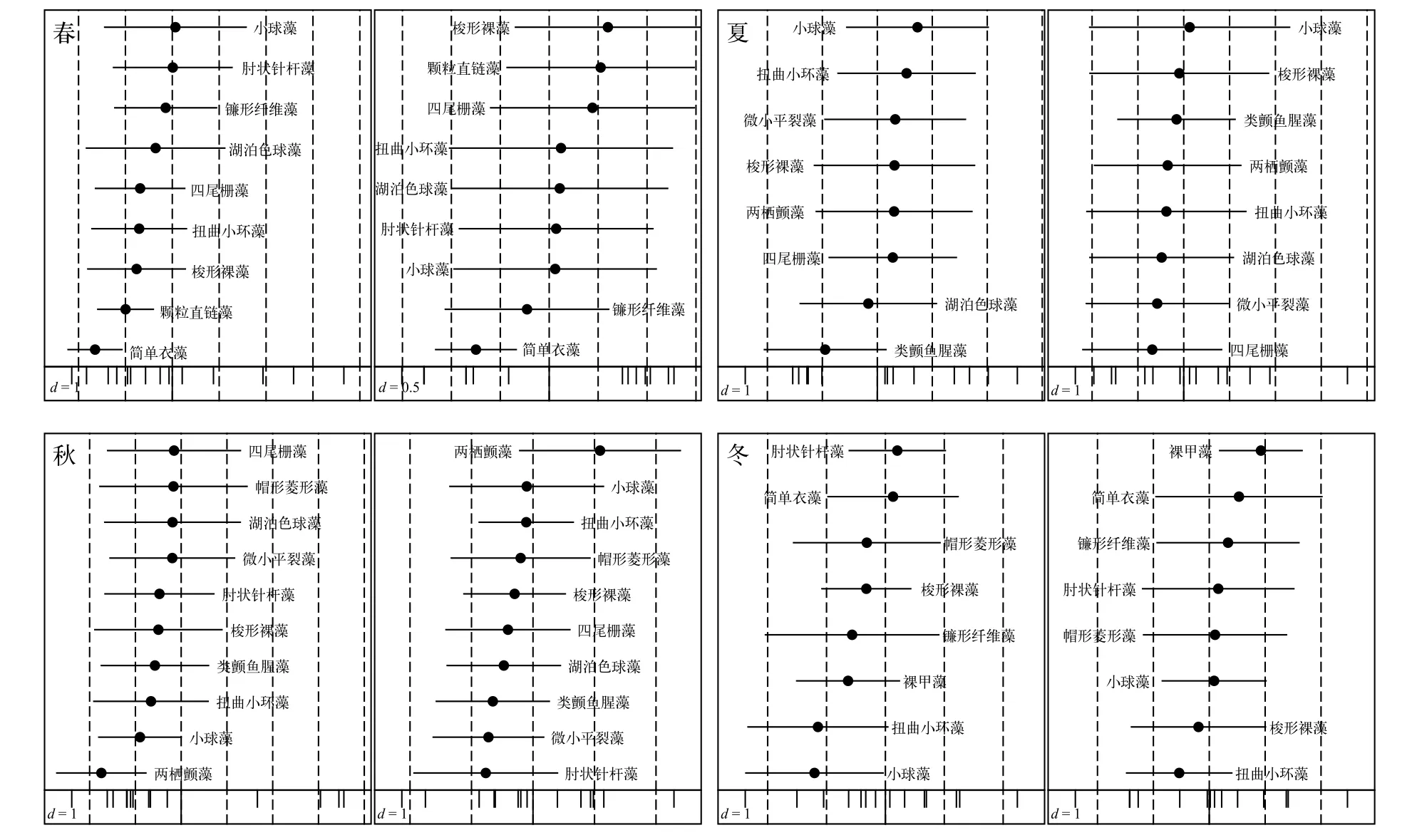
图4 四季优势种生态位和生态位宽度在平均边缘指数分析第一轴和第二轴的分布Fig. 4 Niche position and niche breadth distribution on the first two axes of OMI analysis in each season
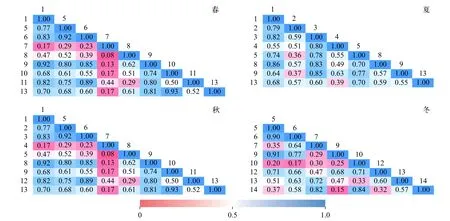
图5 四季优势种群间优势种重叠值Fig. 5 Seasonal niche overlaps among dominant species in each season
平均边缘指数可反映出区域内物种利用资源的能力, 通过OMI值的大小可将物种划分为边缘物种和非边缘物种[30]。本次调查中OMI值较低(小于1)的种类共12种(图2), 利用资源能力较强, 可划归为非边缘物种。通过显著性检验(P<0.05)的OMI分析发现, 除春、秋四尾栅藻以及冬季梭形裸藻外,其余9种OMI值较低的优势种均出现了较明显的生态位分化。以夏季为例,OMI值较小的是类颤鱼腥藻(0.25)、微小平裂藻(0.46)、两栖颤藻(0.33)和小球藻(0.56), 同时这几种也是生态位较宽的藻类, 可能通过分泌化感物质和水生植物竞争从而获得较强的竞争能力, 抑制其他藻类生长, 也可能是对生境特殊因子的较强适应能力从而占有更多的环境资源。
耐受指数侧重于反映物种对环境的适应能力,其TOI值的大小可反映出物种对环境适应能力的强弱[31]。通常来看, 耐受性较强的种类生态位宽度也较大, 如TOI值较高的夏季颤藻(3.25)、类颤鱼腥藻(3.08)和冬季裸甲藻(2.67), 它们的生态位宽度均超过2。
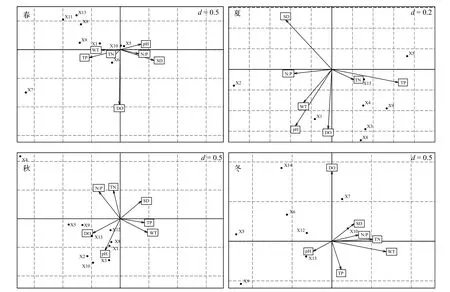
图6 各季节优势种与环境因子之间的典范对应分析Fig. 6 Canonical correspondence analysis between dominant species and environmental factors in different seasons
生态位重叠反映了物种间分享或竞争共同资源的现象, Wathne等[32]认为生态位重叠值大于0.6表示种间生态位重叠较显著。本研究发现生态位重叠值超过0.90的共有7组优势种, 超过0.80的共有19组优势种。这19组优势种的种间生态位重叠显著, 反映出这些物种对资源利用的方式相似, 可能在栖息水层、溶氧、温度、光照、水流、营养盐水平等方面有相似的需求。一些硅藻如菱形藻、小环藻、针杆藻等属浮游和底栖(附生)的兼性浮游植物, 在富营养化水体中一年四季都能形成优势种群, 以春季和秋季最盛。裸藻多喜欢生活在有机质丰富的静水水体中, 温暖季节常大量繁殖成优势种形成膜状或云彩状水华, 是水肥的标志[21]。因此在适温季节, 小环藻与裸藻对环境资源的需求比较相似, 极易出现生态位重叠显著的情况, 这也与本研究结果中春季肘状针杆藻和梭形裸藻以及秋季扭曲小环藻和梭形裸藻重叠值最高(0.93)一致。此外,种间对于资源利用的竞争还受到资源供应的影响,资源供应短缺时竞争程度会趋于加强。
从本研究整体来看, 各优势种生态位参数表现出了较好的一致性。以四季共有优势种小球藻为例, 在夏秋两季, 其生态位宽度均超过2, 属广生态位种类, 平均边缘指数均小于1, 属非边缘物种, 耐受指数也较高, 表现出对环境较强的适应能力。以上表明小球藻生态位宽度、平均边缘指数和耐受指数具有较好的一致性。
3.3 优势种与环境因子的关系
CCA排序可以分析不同物种与环境因子的内在关系, 反映出物种对环境的需求程度[2]。而物种对环境的利用能力也是决定其生态位宽度的重要因素, 环境因子的改变将导致物种对资源的利用分化, 从而使物种间的生态位重叠发生改变[33]。从本次浮游植物优势种与环境因子的CCA分析可以看出, 夏季常见富营养化水体指示种如两栖颤藻、四尾栅藻和梭形裸藻等种类与TN和TP具有较强的正相关性, 而高温和高营养盐水平正是富营养化湖泊在夏季水华爆发的主要原因之一, 表明长湖与其他长江中下游典型富营养化浅水湖泊的研究一致[29]。春季水温由低升高, 此时小球藻生态位宽度适中,而平均边缘指数较低, 耐受指数较高, 表现出了对低温较强的适应性, 这与王菁等[34]的研究结果一致。秋季水温开始较夏季有所回落, 此时硅藻门帽形菱形藻表现出较强的环境适应能力, 与pH有较大正相关性, 生态位宽度和耐受指数均较高。此外,pH、DO、N/P以及SD也是影响长湖浮游植物优势种分布的重要环境因子, 这与长湖地理形状、地貌特征及其他因素如水流速度、降雨量、进出水口及水草分布等有关。
3.4 长湖浮游植物生态位的演替
本次调查是继2012年[11]以来第二次对长湖浮游植物生态位的系统研究, 通过比较发现, 长湖浮游植物的种类组成、优势种以及丰度均没有明显变化, 种类组成仍以绿藻门和蓝藻门为主, 优势种也主要是富营养化水体常见种类, 其中小球藻和扭曲小环藻在两次调查中均为四季共有优势种。各站点丰度均超过106cells/L, 仍处于富营养化状态。本次调查的丰度较2012年有升高趋势, 其中冬季裸甲藻数量明显增多, 为冬季优势种, 而裸甲藻并未出现在2012年的优势种中。根据生态位宽度而划分的广生态位种类和狭生态位种类数量有所增加, 广生态位种类仍以小球藻和扭曲小环藻为代表, 又增加了两栖颤藻、类颤鱼腥藻和裸甲藻等种类, 这些种类通常是富营养化水体的常见优势种,表明长湖水域仍处在富营养化状态, 这与浮游植物种类组成、优势种以及丰度所对长湖的评价结果一致。本研究增加了平均边缘指数和耐受指数分析, 对生态位宽度和生态位重叠进行物种水平分布的测度分析, 揭示一定环境梯度下物种的生态位分布, 并通过种间关系深入分析浮游植物的生态位变化, 也较好地反映了物种对所在环境的适应性和对资源的利用能力。以四季共有优势种扭曲小环藻、小球藻和梭形裸藻为例, 可划归为广生态位种类, 属非边缘物种, 耐受指数也较高, 表现出对环境较强的适应能力, 这些优势种的存在对于湖泊的富营养化治理以及水华的预防都是较难解决的问题。通常认为外部环境因子改善和内部生物种群恢复二者有机结合是恢复湖泊健康生态系统的重要途径, 主要包括控源截污、高等水生植物尤其是沉水植物的种群恢复以及大型滤食性底栖动物的增加等, 上述生态修复措施可以更好地发挥种间竞争和密度制约机制作用下的生态系统组织功能, 实现抑制先锋种或优势种的生长与扩散, 促进其他边缘物种的生长与繁殖, 最终形成良性平衡状态的湖泊生态系统[25,27]。
建议长湖下一步增加对各进出水口营养盐水平和水动力条件等监测并同时开展全面生态调查如滤食性鱼类的摄食压力、高等水生植物、底栖生物及周丛生物的研究, 充分考虑多种环境因子如水文和高等水生植物的克藻效应等措施以控制或减缓藻类水华暴发, 是较好的生态修复手段[35,36]。
