水稻OPR基因的克隆及其在烟草中抗镉性分析
2020-06-13代婷婷姚新转吕立堂
夏 凡, 代婷婷, 姚新转, 吕立堂,
(1.贵州大学生命科学学院/农业生物工程研究院,山地植物资源保护与种质创新省部共建教育部重点实验室, 贵阳 550025;2.贵州大学茶学院, 贵阳 550025)
重金属镉的污染在我国非常严重,它能造成植物生长系统紊乱,导致农作物大量减产甚至死亡[1]。随着基因工程的发展,挖掘抗逆相关基因能够减轻逆境危害,有助于深入地阐释植物在不良环境中的抗逆机制,也为作物的抗逆性分子育种提供基础。12-氧-植物二烯酸还原酶( 12-oxo-phytodienoic acid reductase,OPR) 是茉莉酸生物合成途径的关键性酶,茉莉酸类的化合物( jasmonic acids,JA)也在植物生长过程中发挥着极其重要的作用[2]。茉莉酸在植物体内作为一种内源信号分子,当植物体受到伤害的时候,未受伤害的部位可以通过它让植物进入警戒状态,以此来抵御致病菌和昆虫的危害[3]。
研究发现,茉莉酸化合物参与非生物胁迫的反应,如低温[4]。OPR基因的功能已经在玉米[5]、番茄[6-7]、水稻[8]和豌豆[9]中得到验证,但关于水稻OPR基因抗镉研究目前还未见报告。本研究利用现代分子生物学技术,选用模式植物烟草(Nicotianatabacum),将克隆得到的OPR基因通过农杆菌介导的方法遗传转化到烟草植株体内,应用实时荧光定量PCR,对重金属(CdCl2)处理条件下的OPR基因表达情况及相关基因进行分析,为非生物胁迫分子机制的研究提供理论基础,为作物在抗逆境胁迫及抗重金属污染提供候选基因。
1 材料与方法
1.1 材 料
烟草(NicotianatabacumL. Xanthin)种子由T 1代烟草植株获得,PCR引物由上海英捷生物技术有限公司合成;其它相关试剂采购于天津富宇精细化工有限公司。
1.2 基因克隆与载体构建
利用报道的水稻中的OPR基因,在GenBank数据库中BlastP搜素,得到同源基因的cDNA序列及氨基酸。根据得到的cDNA序列设计并合成引物,基因克隆和载体构建参考邹冰杰等[10]的方法。
1.3 遗传转化及分子鉴定
采用农杆菌(Agrobacteriumtumefaciens)介导的遗传转化方法转化烟草[11]。以野生型烟草植株叶片为外植体,0.1%升汞消毒后进行遗传转化,4周后愈伤组织长出,进行继代培养获得抗性芽,抗性芽长至2~3 cm,然后将抗性芽切下,置于生根培养基上生根培养。2周后待无菌苗长至4~7 cm,炼苗2 d后移栽到花盆中。从烟草叶片中提取基因组DNA,利用根据OPR序列设计检测引物OPRP 1:5’-CCTCTTCTTCCACGACTGCT-3’;P 2:5’-TCGTCAGCACCTTCTTCTCC-3’,PCR反应体系(表1)及反应条件(表2)。

注:RB为T-DNA右边界;35 S为CaMV;tNOS为终止子;LB为T-DNA左边界。
待反应完毕后,将扩增产物经过1.0% 琼脂糖凝胶电泳,在凝胶成像系统下检测结果。
表1 PCR反应体系

试剂体积/ μL基因组DNA2.0ddH2O6.4正向引物(10μM)0.8反向引物(10μM)0.8 Premix Ex Taq10.0
表2 PCR反应条件
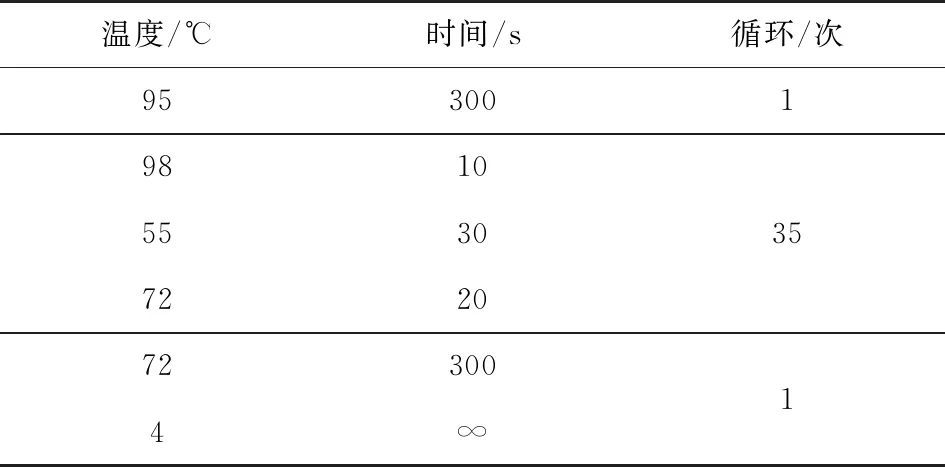
温度/℃时间/s循环/次95300198105530357220723004∞1
1.4 实时荧光定量PCR
用300μmol·L-1CdCl2溶液胁迫处理前胁迫处理后转基因烟草和野生型烟草的叶cDNA为模板。参考OPR基因序列,设计基因的RT-PCR扩增引物,实时定量分析基因的相对表达量。引物设计序列为:F 5′CATGCTCAGTGCCATCAAAG-3′;R 5′CCCAATGAGCAGGAAGAAATG-3′。以烟草Actin基因作内参对照(F:5′TGGTTAAGGCTGGATTTGCT-3′;R:5′TGCATCCTTTGACCCATAC-3′), 在NCBI数据库中找到已报道的烟草抗镉相关基因Snakin-2(Genebank:XM_009623044.2,F:5′TCTCCTTGCTCCTCCTCGAT-3′;R:5′TCAGTGTTGCCAGAAGTGCC-3′)和GRP-2(Genebank:NM_001311181.1F:5′TGGACAGAGGGCTAAGGGAA-3′;R:5′TCAGGGCCGGTAACATCAAC-3′),设计各基因的RT-PCR扩增引物并设计Real-time PCR的反应体系(表3)及反应条件(表4)。
表3 Real-time PCR反应体系

试剂体积/μL2×Power SYBR Green PCR Master Mix 10.0正向引物(10μM)1.0反向引物(10μM)1.0cDNA2.0ddH2O up to20.0
表4 Real-time PCR反应条件
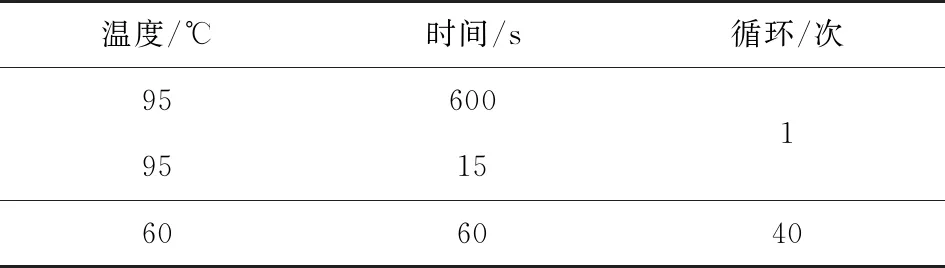
温度/℃时间/s循环/次95600 95151606040
1.5 胁迫处理
用CdCl2溶液(300μmol·L-1)模拟重金属污染。30 d后,选择长势相似的转基因株系和野生型株系,转基因植株经过鉴定呈阳性的植株为试验材料,用CdCl2(300μmol·L-1)溶液胁迫处理,15 d后拍照并剪取叶片,做好标记保存于-80 ℃冰箱中待用。选取T 1代转基因株系(TP 7、TP 8、TP 12)和WT植株的种子为材料,放入含有3个浓度梯度的(0、150、300μmol·L-1)CdCl2溶液的滤纸上萌发进行镉胁迫分析实验(滤纸和空皿提前高压灭菌),分别在第0、5、10天和第15天统计种子发芽率,设置4个重复。
1.6 植物生理生化指标的测定
超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性和丙二醛(MDA)含量都是使用实验室酶标仪来测定。实验选取生长状况基本一致,选取处理前(-80 ℃冰箱保存)和处理后的株系,称取约0.1 g叶片用液氮研磨均匀,加入提取液,酶活测定方法参考苏州科铭生物技术有限公司试剂盒说明书的实验步骤,进行植物生理生化指标的测定。试验3次重复,结果取平均值。
1.7 实验数据的统计学分析
实验所有的数据采用平均值±标准差表示。各平均值差异性的统计学分析是利用SPSS 22.0软件包分析完成。
2 结果分析
2.1 种子的抗镉性分析
挑选饱满的野生型和3株转基因烟草(TP 7、TP 8和TP 12)的种子各50颗,放入含有3个浓度(0、150、300μmol·L-1)梯度的CdCl2溶液的滤纸上萌发进行镉胁迫(滤纸和空皿提前高压灭菌),分别在不同时间段(0 d、5 d、10 d、15 d)统计种子的发芽率。当Cd2+浓度在300μmol·L-1时,转基因种子发芽率极显著(p<0.01)高于野生型,15 d时,转基因种子发芽率在65%左右,而野生型烟草种子发芽率不到20%。实验结果表明,转基因烟草的种子在含有CdCl2溶液的处理下,表现出了一定的抗镉性(图2)。

注:“*”和“**”分别表示所测定项目在转基因与野生型植株之间差异显著(p<0.05)和极显著(p<0.01);WT为野生型(wild type);TP 7~TP 12为转基因株系(Transgenic lines);A、B浓度为0;C、D浓度为150 μmol·L-1;E、F浓度为300 μmol·L-1。
2.2 转OPR基因对烟草植株生理生化指标的影响
在自然条件下,选择生长30 d状况基本一致的烟草植株,用300μmol·L-1CdCl2溶液模拟重金属浇灌, 处理时间为15 d,然后取样品进行相关生理生化指标的测定。
在300μmol·L-1CdCl2溶液胁迫处理下,时间为15 d时(图5),转基因植株和野生型植株的SOD酶活性和POD酶活性都上升,转基因植株的SOD酶活性和POD酶活性都明显高于野生型植株,转基因植株的SOD和POD酶活性分别为277.5 U·g-1和328U·g-1,而野生型植株的SOD和POD酶活性分别为169.9 U·g-1和195.0 U·g-1,与野生型植株差异达显著水平;转基因植物CAT酶的活性增加,比野生型CAT活性增加了57%;野生型植株在处理15 d时MDA含量迅速升高,转基因植株MDA含量升高相对于比较缓慢。

图3 Cd2+胁迫转基因和野生型烟草

图4 Cd2+胁迫下转基因株系和野生型烟草SOD、POD、CAT活性和MDA含量
2.3 转OPR基因烟草相关表达基因的影响
实时荧光定量表达分析,在正常条件下,转OPR基因表达以及抗镉相关基因Snakin-2和GRP-2表达整体水平相似;胁迫15 d,转OPR基因在烟草中的平均表达量平均是野生型的2.48倍;Snakin-2基因的平均表达量是野生型的1.70倍;GRP-2基因的平均表达量是野生型的1.45倍(图5)。结果表明,Cd2+胁迫转OPR基因植株可以显著诱导烟草中Snakin-2和GRP-2基因的上调。说明该基因可能参与镉胁迫的渗透调节过程来提高植物抗镉性。

图5 转OPR基因烟草相关基因表达的影响
3 讨 论
OPR基因在植物调控耐受性方面具有重要作用[12-14]。本研究从水稻中克隆获得了OPR基因,研究其在作物抗重金属的功能,结果发现,OPR基因能够对CdCl2胁迫作出应答,OPR基因及烟草抗镉相关基因Snakin-2和GRP-2对Cd2+处理后,OPR、Snakin-2和GRP-2基因均表达上调。烟草中Snakin-2和GRP-2基因均对Cd2+有一定的耐受性使其基因表达上调[15]与该实验结果趋势基本一致。在Cd2+浓度为300μmol·L-1胁迫下,转基因植株种子发芽率显著高于野生型,说明转OPR基因植株在重金属胁迫过程中表现出较强的抗镉能力。重金属胁迫使植物体内产生大量活性氧(如超氧阴离子、过氧化氢等),过多的积累导致细胞膜通透性变大,当超出了植株可耐的范围,植物将受害甚至导致死亡[16]。重金属胁迫导致植株组织的氧化胁迫和膜损伤,改变酶系活性,对植株生长产生一定的影响。植物通过抗氧化酶系统来提高植株的耐抗性[17]。大量研究表明,重金属Cd2+在一定浓度梯度范围内的胁迫能诱导植物SOD、POD、CAT等抗氧化酶活性的增加,从而及时清理掉体内产生的多余活性氧,维持植物的正常机能,当重金属浓度超过了植株可承受的最大范围就有可能使植物失去维持活性氧平衡的能力[18-19]。研究表明,在不同浓度梯度镉离子的胁迫处理下,测得转基因植株的SOD和POD的酶活性都显著高于野生型植株,而转基因植株的MDA含量虽然有所增加,却低于野生型植株,说明转OPR基因可以清除活性氧和改善膜脂过氧化,并减轻逆境对烟草植株的伤害。
