冬季升温对高山生态系统碳氮循环过程的影响
2020-06-11石培礼
宗 宁, 石培礼
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学, 北京 100049
自工业化革命以来,人类活动包括化石燃料的燃烧和土地利用及覆被变化已使地球大气层中CO2的浓度上升了30%,其结果造成地球表面的平均温度在 20世纪升高了0.67 ℃,预计到本世纪末地球表面的平均温度还将继续上升1.8―4.0 ℃[1]。在全球变暖的背景下,高纬度和高海拔地区升温趋势更加明显,这在实际观测与模型模拟结果中均得到验证[1- 2]。全球温度升高存在明显的季节差异性,呈现出非生长季升温显著高于生长季的季节非对称趋势,在高纬度和高海拔地区更是如此[1,3- 7]。同时,在未来气候变化情景下,这种季节非对称性升温趋势会更加显著[5,8]。虽然目前针对升温对陆地生态系统的影响已有诸多研究,但主要集中于全年升温的影响,季节非对称性升温对生态系统影响的实验研究却鲜有开展[9],在高海拔、高纬度地区忽略了季节非对称性升温的事实。
冬季升温的直接后果是导致积雪覆盖面积的减少以及积雪厚度的降低。有报道显示,地球陆地表面大约60%的面积经历季节性积雪覆盖或土壤冰冻过程,从高纬度寒带、温带森林和中纬度森林和草地地区,以及许多高山地区[10]。季节性积雪覆盖的生态系统在全球生物地球化学循环中起着重要作用,尤其是碳循环过程[11]。例如高纬度地区储存了60%以上的陆地有机碳[12- 13],季节性积雪覆盖的中纬度地区储存了20%―30%[14- 15]。积雪覆盖地区对气候变化异常敏感,极小幅度的气温变化都会引起这些地区生态系统碳氮循环的显著变化[11]。同时,冬季增温还会影响冰冻层厚度和冻融交替循环过程。冬季一般被认为是植物休眠季节,但在温带生态系统,一些植物仍然能进行光合作用[16- 17],并能吸收养分[18- 19]。许多能影响植物生长季节生理生态活动的过程,如土壤矿化作用、养分淋溶、土壤物理性质的改变以及温室气体排放过程等都与冬季积雪覆盖及土壤冰冻厚度有关[11,20- 22]。在高寒生态系统,由于积雪覆盖的绝热作用,积雪下微生物代谢活动活跃,且能利用溶解性有机碳[11,23- 24],这对接下来生长季植物生长发育及碳氮循环都会产生重要影响[24- 25]。
由此可见,冬季是生态过程的关键时期,冬季升温是驱动生态系统结构和功能发生变化的重要因素[8,26]。在高纬度地区,气候变化对冬季的影响作用更大。IPCC[1]报道显示,在过去的30年,北半球高纬度地区冬季升温幅度最大,在未来气候变暖背景下会更加显著。在北半球的冬季,由于升温状况下大气的水分承载能力增强,冬季升温将伴随着更多的降雨[5]。本文系统综述了目前冬季升温的现状及影响,阐述了模拟冬季升温的试验方法,着重介绍了冬季升温对土壤碳、氮循环产生影响的机制,并由此引起的植物生理活动的改变,综述了野外冬季增温实验的主要研究成果,并对存在的问题进行初步分析,期望对有关研究起到一定的借鉴作用。
1 冬季升温现状及影响
1.1 目前冬季升温的现状
IPCC第四次评估报告显示,过去30年观测温度显示,冬季升温显著高于其他季节,而在未来气候变化情景下这种现象会更加严重[1]。统计分析结果显示,全球、中国和青藏高原冬季升温显著高于全年升温幅度,分别0.07―0.42、-0.13―0.52、0.31―0.47 ℃/10年,其中青藏高原的升温幅度大于全球和中国(表1)。
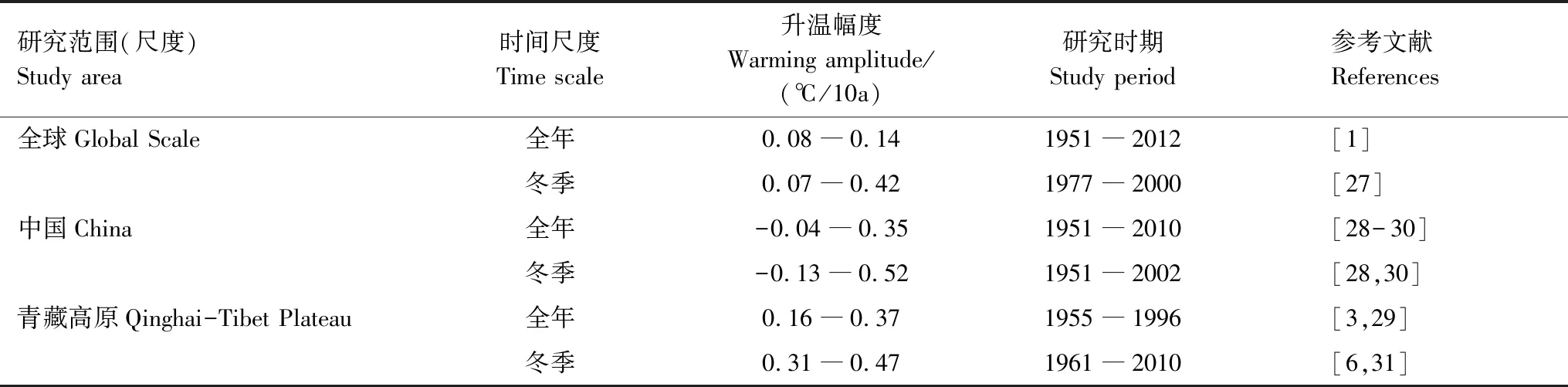
表1 全球、中国及青藏高原全年升温与冬季升温的幅度比较
1.2 冬季升温的影响
1.2.1积雪覆盖变化及其对物质循环的影响
冬季升温的直接后果是导致积雪覆盖面积的减少及积雪厚度的降低。从1972年到2003年,北半球积雪覆盖面积已经减少了近10%[32],其中积雪覆盖面积减少最显著的区域还包括高山地区,这都是温度增加显著的区域[33]。积雪覆盖面积减少的趋势会持续增加,到21世纪末欧洲大陆被积雪覆盖的时间会减少40%―80%[34]。冬季积雪的减少会引起土壤与空气温度隔绝作用的降低,积雪作为热量缓冲库的作用会减弱,这会增大土壤温度的变异,从而增加土壤冻融循环的频率[35],并深刻影响生态系统土壤碳氮循环过程及存储量[20,36- 37],继而对全球变暖产生正反馈效应[5,38]。一项关于积雪深度对陆地生态系统氮循环的研究表明,积雪深度增加提高了4.5%的植被氮和35.9%的土壤微生物氮含量,但降低了34.1%的土壤氧化亚氮排放,而对土壤溶解性有机氮、总无机氮以及氮流失无显著影响[37]。在全球季节性积雪分布区域,一些地区的积雪减少甚至不再出现,而少数地区由于极端气候事件出现的频率增加,可能会遭遇更大的积雪。在海拔1000―2393 m的大部分高山或亚高山地区,积雪厚度和持续期将进一步减少和缩短;而20世纪70年代以来的青藏高原,积雪变化的总趋势是每年积雪持续期缩短[1]。
1.2.2冻融交替变化及其对物质循环的影响
土壤冻融交替是中高纬度和高海拔地区常见的自然现象[38- 39],会通过改变土壤结构和水热状况直接影响土壤元素的生物地球化学循环过程。春季融雪时期的气温变化引起的冻融交替是影响土壤氮素转化过程和氮素流失的重要因素之一[40- 41]。冻融循环过程通过影响土壤理化性质和微生物活动等各种过程而影响土壤中氮素转化及生态系统养分流失[38,40,42- 43]。Zhou等[44]研究发现,冻融交替增加了不同森林类型土壤净氮矿化速率,进而导致土壤无机氮含量升高。这种由于冻融交替增加引起的无机氮含量的增加,在植物不活跃的季节有一定程度的氮流失风险,有关研究发现冻融交替时期输出到临近溪流中的硝态氮主要来源与土壤微生物活动有关[45]。同时,随着冻融循环次数的增加,土壤微生物氮呈现先增加后降低的趋势[46- 47],且较大的冻融温差使微生物量氮减少[48]。在全球变暖的条件下,高纬度和高海拔地区土壤将经受更广泛和频繁的冻融交替作用,冻结强度降低、冻结持续时间缩短、冻融循环次数增加[49- 50],这会对有积雪覆盖生态系统的碳氮循环过程产生深刻影响,导致长期贮存在土壤中有机质以温室气体的形式排放出来,对气候变暖产生正反馈作用[39]。
2 冬季升温对土壤碳氮循环与微生物的影响
2.1 土壤碳循环过程
冬季土壤呼吸是年际CO2排放的重要组成部分,显著影响生态系统碳收支过程。相关研究表明,中高纬度地区冬季CO2排放是年际碳收支的重要组成部分,冬季碳排放的观测对于估算年际碳排放交换量具有重要意义。一项关于冬季和生长季CO2通量的对比观测表明,不考虑冬季CO2通量会高估71%落叶森林和111%针叶林的年际净生态系统碳交换(NEE)[23]。冬季CO2通量占全年碳排放的比例在不同生态系统约为4.9%―26%(表2),而冬季氮通量占全年的比例约为17%[51]。由此可见,冬季碳氮循环通量的观测对于准确研究冬季生物地球化学循环过程至关重要,而冬季升温对生态系统年际碳氮循环过程会产生重要影响。

表2 冬季CO2通量占全年通量的比例
冬季升温引起的积雪覆盖和冻融交替的变化均会对高山生态系统碳循环过程产生重要影响。一项关于积雪覆盖厚度变化对陆地碳循环的meta分析表明,增加积雪覆盖厚度提高了18.4%的凋落物量和15.5%的CO2排放量,显著降低了5.8%的凋落物碳氮比和28.5%的总碳含量,但对土壤溶解性有机碳(DOC)和根系生物量无显著影响,积雪深度增加下土壤温度和湿度的增加是引起这些变化的主要原因[57]。虽然冬季碳排放的绝对量在不同生态系统类型之间变异较大,但冬季碳排放的敏感性对季节性积雪覆盖变化的响应更大。积雪覆盖变化和室内试验都表明,冻融交替引起的可利用碳底物的变化(如土壤DOC)会显著影响冬季土壤微生物活动和CO2通量[23,58- 59]。相对于夏季,冬季土壤中溶解性单糖和氨基酸增加了8―40倍,主要原因是冻融交替过程对植物根系和微生物细胞的物理伤害,并释放溶解性有机碳[11,60]。即便是在冰点以下温度,碳的季节性和可利用性变化使得土壤微生物对溶解性底物存在偏好利用,微生物通过对土壤中溶解性有机碳的偏好吸收来调节渗透压,由此来抵御寒冷的伤害和维持细胞间酶的活性[61- 62]。土壤DOC可利用性是寒冷季节微生物呼吸的重要影响因素,冬季升温引起的积雪减少会引起土壤DOC的固定化,并引起其他生物地球化学过程的变化(如氮的水溶性和气体性流失、CO2通量)[63- 64],对生长季的生态系统碳氮循环会产生重要影响[65]。此外,冬季升温会导致高纬度、高海拔地区冻土层融化加剧,并刺激土壤有机质的分解,降低其生态系统固碳能力。Natali等[66]等通过设置雪墙增温的研究发现,冬季1.6 ℃的升温幅度增加了10%的冻土融化深度,其中包含2 kg C m-2,但这并未导致生长季累积呼吸的增加,这可能是由于土壤活动层达到饱和而对生长季土壤呼吸产生抑制作用;然而冬季升温导致冬季土壤呼吸加倍,从而改变了年际碳平衡。由此可见,高纬度苔原地区冻土层对冬季升温反应敏感,使得碳排放增加而正反馈气候变暖过程。
2.2 土壤氮循环过程
氮素是影响陆地生态系统初级生产的最重要元素之一,氮素的可利用性与生态系统的维持、演替和稳定性的维持密切相关[67- 68],对于受氮限制的高寒生态系统尤其显著。因为受环境因子的限制,高寒土壤矿化速率很低,故导致氮素可利用性很低[69]。一般来说,升温会促进土壤氮矿化速率[51,70],在生长季节会提高土壤氮的可利用性,满足植物生长发育的需求,但这是在水分不欠缺的前提下发生的情况[71- 73]。而在冬季多数植物处于休眠状态,冬季升温以及伴随的冻融交替过程的增加可能会增加土壤氮的损失[74]。一项关于日本北部森林的综合分析表明,冬季气候变化导致的降雪量减少会引起积雪覆盖的热量隔绝效应下降,并使得土壤表层冻融交替循环频率和幅度的增加[75],对土壤净氨化作用影响较大,而对硝化作用影响较小。流域和区域尺度冻融交替循环增加引起的不同生态系统之间的土壤氮动态和流失差异主要归因于土壤氮库和微生物活性的变化。进一步综合分析表明,冬季气候变化对冬季和融雪季节土壤氮素生物地球化学循环过程(如土壤氮库大小、微生物氮转化过程)有重要的影响,且具有延滞效应,对接下来生长季的氮循环过程产生重要影响[75]。这种延滞效应在另外一项原位试验中得到验证,全年升温对土壤净氮矿化速率和淋溶损失影响很小,而冬季升温处理对土壤氮周转速率影响较大,冬季升温处理下生长季土壤净氮矿化速率是对照及全年升温处理的两倍[74]。另外一项关于积雪覆盖变化对陆地生态系统氮循环的meta分析表明,积雪深度增加提高了4.5%的植被氮和35.9%的土壤微生物氮含量,但降低了34.1%的土壤N2O排放,而对土壤溶解性有机氮、总无机氮、硝态氮以及氮流失无显著影响。积雪深度增加会降低土壤硝化作用(-24.8%),说明土壤氮向硝态氮转化减少,这可能有助于降低土壤氮的流失。在落叶森林和针叶林的研究也表明,积雪较厚年份生态系统氮固持较高,而氮输出较少[76-77], 而积雪深度降低会导致生态系统氮输出增加[58,78-79]。由于积雪覆盖增加会使得土壤隔绝冷空气,微生物氮固持增加。积雪较厚时,开始土壤氮会被固持在微生物体内,在生长季开始时会被植被吸收利用[24,80]。同时,较浅的积雪覆盖和强烈的土壤冰冻会导致越冬期植物根系的死亡[81- 84],引起植物对氮吸收的降低,这会导致生态系统氮输入的增加。由此可见,积雪的年际变化会通过融雪之后植物可吸收氮的含量和可利用性来间接影响生态系统碳储存。
积雪覆盖的变化还会影响N2O排放,冬季N2O排放通量与积雪覆盖的厚度和持续时间以及微生物活性有关[49,85]。实地观测显示,高寒生态系统冬季N2O排放比生长季高出一个数量级[86-87],而在温带地区的观测表明冬季N2O排放占年际排放量的一半[88-89],冬季高的N2O排放量可能与积雪下逐步加重的厌氧过程以及冬季植物对养分竞争的减少有关[11]。土壤微生物氮的矿化与固持的平衡过程受土壤溶解性有机碳含量的强烈影响,而积雪深度和冰冻程度的变化通过影响凋落物分解和碎花过程来土壤溶解性有机碳含量[11]。
2.3 土壤微生物与植物养分吸收的季节耦联关系
土壤微生物活性在生长季与冬季不同,因为不活跃季节植物向土壤中碳输入减少,且冰冻期间土壤水分的流动性降低,从而改变土壤养分的可利用性和微生物活性。研究表明,土壤微生物胞外酶对升温无显著影响,升温显著提高微生物碳含量以及磷酸酶活性,而升温对细菌、真菌生物量影响较小[90]。冬季升温导致的冻融交替过程的增加同样对土壤微生物特性产生重要影响。对苔原和泰加林四种土壤室内冻融交替培养实验显示:每次土壤冰冻、融化后,土壤微生物呼吸开始出现脉冲式增加(< 24 h),呼吸释放出的碳在第一轮冻融交替过程中最多,而在接下来的冻融交替过程中逐渐减少;土壤净氮矿化速率在第一轮冻融交替后显著增加,但在第三轮后显著降低。冻融交替开始时,土壤碳、氮的响应主要受微生物生物量的控制,而在反复冻融交替后,微生物群落的降低导致土壤有机质分解减少,这也是碳、氮响应的减少的主要原因[91]。
同时,冬季升温会打破土壤微生物养分释放与植物养分获取的季节耦联关系。已有研究表明,在养分受限的高寒生态系统中,为减少对有效养分的竞争植物与微生物对氮素的利用表现出季节分化的特征[61,92-93]。植物主要在生长季节吸收氮素供给生长发育,而土壤微生物的氮固持主要发生在寒冷的时期(晚秋、冬季),土壤微生物将大量的氮素储存在体内[80,93]。冬春转换时期嗜冷微生物的死亡、底物可利用性的降低、冻融交替的破坏作用以及渗透压的变化会导致氮素从土壤微生物体内被释放出来[61],供生长季植物返青和生长季利用[80,94-95],这就是植物与微生物之间养分利用的季节耦联关系。在自然条件下,冬春转换时期的养分脉冲式释放是高寒生态系统在年际上最大的养分输入过程[61,96],植物与微生物之间这种养分耦联关系在高寒苔原[80]、亚北极白桦林[16]、亚北极白桦冻原[97]、亚北极高寒草甸[94,96]等生态系统的研究中均有报道。同时也有研究发现,冬春转换时期即使温度接近冰点或低于冰点,植物地上部分虽未萌发,但群落中优势植物的根系便能吸收氮素[98-99],这种机制对于防止养分流失具有重要的生态学意义。冬季升温对高寒生态系统植物与土壤微生物养分利用的季节耦联关系的影响主要体现在以下三个方面。首先,冬季升温的影响体现在寒冷季节。冬季升温会引起寒冷时期土壤微生物数量的减少[96],这可能与秋冬冻融时期的影响有关。冬季升温导致微生物养分释放的减少和供给春季植物生长养分数量的降低,反过来影响更高营养级的生产[61]。其次,冬季升温的影响体现在冬春转换的冻融时期。冬季升温会导致冻融事件发生提前、冻融交替频率增加,但此时温度仍较低植物地上部分尚未萌发,可能会打破微生物养分释放与植物养分获取的季节耦联关系。第三,冬季升温的影响还体现在对生态系统的延滞效应。植物与微生物养分吸收的季节耦联关系被打破后可能会导致养分流失增加,进而影响生长季资源供应的数量,并进一步影响生态系统生产、结构与功能等。
3 冬季升温对植物生理-生态过程的影响
3.1 植被物候期
物候是反映生态系统对气候变化响应的一个重要指示因子,是响应温度变化最敏感、最容易观察到的特征[100]。受气候变化的影响,高山生态系统植物物候呈现出不同程度的春季物候提前和秋季物候延迟[101-103],物候的改变会进一步影响草地的生产力,但目前关于冬季升温对物候影响的研究仍然较少。基于样点试验的研究,利用冬季积雪覆盖的方法研究冬季升温对苔原生态系统过程的影响发现,冬季升温导致地上生产力增加20%,而全年升温对其无影响。冬季升温延长了生长季,植物发芽时期提前,而凋亡时间推后[66]。Wipf等[104]研究了积雪深度降低和融雪提前对阿尔卑斯山地区高山生态系统树线以上4种广泛分布的植物的影响,研究发现融雪提前会导致所有植物种类的返青期提前,3种植物芽生长期提前,但推后了欧洲越桔(VacciniummyrtillusL.)芽的生长期;融雪提前导致所有植物种类繁殖期提前,积雪覆盖减少进一步提前了植物的开花期和花凋谢期;但物候期的延长并未对植物生长产生积极作用,有三种植物的地上生长由于融雪提前而降低。由此可见,融雪时间对物候期的影响比积雪覆盖要大。
在区域尺度上,同样发现冬季增温对物候期产生重要影响。邱丹和张国胜[105]通过对青海省草本植物物候期观测分析得出:上年10月至当年 4月平均气温升高 1.0 ℃,全省草本平均返青期提早 2.2d左右,上年 10月至当年9月平均气温升高 1.0 ℃,平均黄枯期提早0.4d。而在弃耕地的研究发现,升温并未延迟秋季植物的凋亡时间,升温对物候期的影响同时受升温引起的土壤水分含量降低的调节[4]。区域尺度上气温的升高同样对植物物候有重要影响。青藏高原是近几十年来全球升温最显著的区域之一[103,106],这对高寒生态植物的物候及生产产生一定影响。利用归一化植被指数(NDVI)与温度的关系,Zhang等[103]研究发现冬季温度对植物春季的物候期有重要的影响,冬季升温会导致植物春季物候期的提前。生长季的提前可以为当地牧民提供更长时间的牧草资源[102],而冬季升温对物候的影响还受到其他环境因子的调控。
3.2 植物群落结构
气候变化正在引起冬季气候条件的改变,而由此引起的冻融交替频率的改变具有重要的生态学意义,冻融交替在引起土壤氮循环改变的同时,还会导致植物群落结构发生改变,这在不同类型的生态系统有不同的影响。Kreyling等[22]研究发现,草地生态系统在冻融交替处理后的第一个生长季群落结构变化较大,随着时间的增加冻融交替的影响逐渐消失;而在灌丛生态系统,冻融交替处理导致的群落结构改变发生在第二个生长季,且随着时间的增加这种影响持续增加。同时Kreyling等[22]还发现植物对冻融交替的响应模式与生长类型有关:冻融交替处理后,一些草本植物增加,而灌木盖度降低;冻融交替增加对物种多样性丰富及豆科植物存在的群落影响较小。这表明冬季气候变化是生态系统变化的重要驱动因子,短期的气候事件对生态系统结构有长期的影响,植物群落会调节由于土壤温度增加而导致的植物群落的发展与竞争之间的平衡。
冬季气候变暖导致的积雪覆盖减少同样会引起植物群落结构发生变化。一项研究表明,积雪覆盖减少引起阿拉斯加扁柏(Chamaecyparisnootkatensis)的减少,这种植被芽的耐寒性适应于目前冬季的气候,而根系耐受温度不能高于-5 ℃,在积雪存在时的土壤绝缘隔热保护对该物种的根系具有足够的保护作用。对这种保护作用的实验研究表明,在没有积雪的隔热保护时,当前气候条件会对植物的根系系统造成损害;而在积雪隔热保护下细根损伤仅为10%,从而表明该物种的存在依赖于足够厚的积雪覆盖[107]。冬季变暖情况下,植物生理活动(光合作用)和保护(耐寒性)之间的权衡是植物生存和性能的重要决定因素[5]。因此,植物可以通过减少保护投资和提高生产力来应对气候变暖。然而由于脆弱的植物组织暴露于短期霜冻事件,可能会增加植物霜冻损害的风险。2007年美国东部的春季霜冻事件表明,短期霜冻会导致作物和自然植被的大量被冻伤[108]。天然植被的物种组成可能受到这些霜冻事件的强烈影响,因为霜冻后的恢复力在物种之间存在很大差异。由此可见,由于植物对极端气候事件的忍耐力和恢复力不同,冬季增温引起的极端气候事件会对植物群落结构产生重要影响。
3.3 植物生产及养分利用
冬季升温会对土壤冻融过程产生重要影响,在植物不活跃的季节会增加生态系统氮损失,这会限制生态系统生产对温度的响应过程。研究发现,在干旱年份升温对植物生产无影响,而在湿润年份,冬季升温和全年升温与氮添加处理导致地上生产力加倍,冬季升温与全年升温对生态系统生产的增加幅度相同,而这种变化与降雨的年际变化密切相关[4]。这种由于气候变化引起的群落生产改变会影响土壤与大气碳库间CO2的传输。另外一项研究发现,冬季升温导致20%的ANPP的增加,并提高20%的冠层氮含量。冬季升温提高氮的可利用性,使ANPP生长率较高。升温导致的生产力提高会抵消部分由升温导致的微生物分解速率的加快[66]。而对亚寒带地区苔藓地衣的研究发现,冬季升温事件对绿皮地卷(Peltigeraaphthosa)生长季光合作用无影响,而降低了光滑岩黄耆(Hylocomiumsplendens)48%的光合速率和52%的生长速率,主要原因是地衣不会经历关键物候期(如脆弱器官的生长),而苔藓有明显的物候特征,在温暖冬季事件打破休眠时产生的嫩芽会被冻伤[109]。由此可见,冬季增温对高山生态系统的影响受降雨年际变化的调控,且冬季升温的影响会被土壤微生物分解速率的加快所抵消。
Turner和Henry[110]通过设置全年增温与冬季增温处理,研究模拟增温与氮添加对温带草地植物的氮固存的影响。在春季解冻时期向土壤中添加15N标记物,到生长季结束时,增温或施氮对植物根系和土壤中15N的回收率无影响。经历冬季之后,植物叶片中半数的15N损失,在施氮样方内大量15N从土壤中损失,而在非施氮样方内大量15N增加。到下一个春季,施氮处理中植物组织中15N回收率显著降低,这会抵消全年增温导致的植物组织中15N回收率的提高。15N回收的主要发生在土壤中,且不施氮土壤中15N富集量比施氮土壤高40%。由此可见,增温导致植物组织中氮固持的增加,但这种增加会被氮沉降所导致的生态系统氮损失所抵消,对生态系统产生不确定的影响。
3.4 凋落物分解
植物凋落物作为联系地上和地下的桥梁,是研究植物多样性与土壤微生物多样性之间关系的良好媒介。温度通过促进微生物的活动直接影响分解作用,通过改变土壤湿度和输入到土壤中有机物的数量和质量间接地影响分解作用。在一个较宽的温度范围内,温度升高导致微生物呼吸呈指数增加,加速有机碳向CO2的矿化作用。冬季升温可通过冻融过程影响分解作用。冻结过程会杀死许多存在于正在分解的凋落物和土壤有机质中的微生物,将溶解性有机物释放到土壤中。冻融过程会在物理上破坏土壤团聚体和凋落物的结构,暴露更多的新鲜土壤表面而刺激分解作用。在某些极地生态系统中,一年中秋季、冬季和春季的分解作用占年凋落物质量损失的大多数[111]。也有研究通过实地观测和实验室模拟实验发现,多数凋落物损失发生在秋季,而冬季升温对亚寒带石楠灌丛新鲜凋落物分解无影响[112]。温度对分解作用还有间接影响,高温通过增加蒸发和蒸腾作用而降低土壤湿度。在干旱气候中土壤变干可降低分解速率,但在过湿的土壤中由于湿度限制氧气供应,土壤变干可加速分解作用。
冬季气候变暖导致的积雪覆盖减少同样会引起凋落物分解发生变化。对于具有较厚积雪覆盖的高山生态系统而言,冬季凋落物分解过程与在生长季同等重要。有研究指出,第一年凋落物损失的质量大约占总凋落物质量的30%,其中冬季损失的质量可达全年损失总量的40%―60%[113]。一般来说,积雪覆盖去除后导致的冻融频度提高会加速凋落物的物理破碎,从而提高凋落物的分解率[114]。有研究发现,冬季升温对凋落物分解速率和质量均无影响,因为冬季升温减少积雪覆盖,增加地表温度的变异性[115]。但积雪覆盖导致冬季升温呈现不同的规律,随着积雪深度的增加,凋落物分解率显著提高,而在积雪去除的处理中,年际纤维素分解率比对照处理低40%[116]。这说明,积雪的缓冲保温作用对凋落物分解率的影响可能大于冻融循环对凋落物分解的影响[117]。积雪对凋落物分解的间接影响主要体现在对于土壤微生物的数量及活性上。积雪去除后频繁的冻融循环和更大的温度波动导致的冰冻事件均可以直接影响微生物的死亡率,从而导致凋落物分解率下降,而深雪覆盖的绝热作用仍可以维持足够的热量支持广泛的土壤生物活动[8,118]。
4 问题与展望
4.1 非极地苔原冬季升温的研究相对匮乏
由于高纬度地区生态系统对全球变暖的敏感性和特殊性,目前绝大多数冬季升温的影响都是在极地苔原和高山生态系统中开展,而对于非极地苔原生态系统的研究十分匮乏。有研究表明,冬季气候变化对温带植物的生长发育极其重要[5],故探讨冬季气候变化以及由此带来的极端冰冻事件和气候因子的变异性十分必要。同时,除了研究冬季升温对植物和土壤过程的影响,还要研究冬季升温对病原体、传粉者、取食者以及菌根共生体的影响。
4.2 注意选择合适的模拟冬季升温的方式
作为全球变化生态学的重要实验方法,模拟增温实验是研究全球变暖与陆地生态系统关系的主要方法之一,生态系统增温实验能够为生态模型的预测提供参数估计[119]。由于冬季积雪的存在,冬季增温与全年增温方式存在差异。目前已有的研究中模拟冬季增温主要采用开顶箱增温法[52,120]、红外辐射增温法[4,74,90,110]、积雪覆盖[66,121- 123]和埋设增温管[22,51]等方式。开顶式增温室是在冬季安装增温室,在生长季节撤掉增温装置来模拟冬季增温过程[120]。开顶式增温室维持费用不高,可以用于一些偏远没有电力的高纬度和高海拔地区。但开顶箱存在阻挡积雪、降低混合气体的扩散和湍流等负面效应。红外辐射增温法是利用红外辐射器从冠层上面进行加热,由于能真实地模拟全球变暖机制而被广泛应用于生态系统模拟控制实验中[119]。红外辐射增温能非破坏性地传递能量,而且不改变微环境,对于积雪较厚的地点也可行[119]。但该方法并不能直接加热空气,也不适用冠层密集的植被,且耗费较多电力,在没有电力和高大灌丛及森林地区不适用[119,124]。所以在土壤中埋设加热管道或电缆加热土壤的方法被应用于森林生态系统中,因为其他增温方式在高层灌木和森林生态系统中受到限制。但这种方法不能加热空气和植物地上部分,不能真实地模拟全球变暖对陆地生态系统的影响,且电缆所造成的恒定增温并不能真实的模拟自然状态下全球变暖的季节变化和日变化特征[119]。积雪作为热量传输的绝缘体和辐射屏障[27,38],为冬季增温实验提供了新的方法与可能,通过积雪覆盖隔绝寒冷空气达到增温效果[66,121],为保证不同处理间积雪融化时间的一致,在积雪开始融化前移除雪盖。但该方法只局限于积雪较厚的极地地区,在积雪较少的地区并不适用。
由于这些增温方法在设计、技术和增温机制上的差别,实验增温后陆地植物和生态系统的响应各不相同[119]。未来的研究应针对在相同生态系统中开展不同增温方法的对比研究,比较这些增温方法之间模拟冬季气候变暖机制和温度变化情景的有效性,有利于对陆地生态系统的响应与适应得出更全面和令人信服的结论,也会为不同生态系统选择最合适的增温方法提供科学依据。
4.3 关注冬季升温对植物-土壤微生物之间反馈作用的影响
植物与土壤微生物是陆地生态系统的两个重要组成部分,植物-微生物相互作用在调节群落结构和生态系统功能方面起着重要作用[125],目前越来越多的研究意识到理解全球变化的后果需要深入研究植物与土壤微生物之间的关系[126-127]。植物作为生产者通过叶片的光合作用向根系及土壤生物提供光合产物等资源,而土壤微生物作为分解者为植物提供生长所需的养分。植物的养分获取与土壤微生物的养分固持同时存在,所以在养分受限的生态系统中,植物与土壤微生物对养分存在竞争作用,这在诸多高寒生态系统中都得到验证[128- 130]。目前研究主要针对升温对自然生态系统中植物与微生物的影响,冬季升温对植物与微生物养分季节耦联关系的研究鲜有开展,更没有深入研究其对生长季资源有效性以及植物与微生物的养分利用策略有何延滞效应。
4.4 重点关注冬季升温对生态系统的延滞效应
虽然已有研究发现冬季升温会增加土壤养分流失[26,131],也揭示了冬季升温与全年升温对生态系统的影响存在差异[9,74,132],这种效应受年际间降雨格局的调控[4]。但相关研究更多关注氧化亚氮释放或淋溶等物理过程,并未考虑土壤微生物如何介导养分流失过程,以及冬季升温对生态系统的延滞效应。植物与微生物养分吸收的季节耦联关系被打破后可能会导致养分流失增加,进而影响生长季资源供应的数量,并进一步影响生态系统生产、结构与功能等。同时,研究冬季升温对生长季延滞效应的研究可为深入理解高寒生态系统的群落生产和碳循环等过程提供机理解释。为此,研究冬季升温下植物与微生物的养分利用关系及其延滞效应,对于探讨未来气候变化情景下生态系统碳、氮循环的响应机制势在必行。
