三峡库区消落带池杉-土壤碳氮磷生态化学计量特征
2020-06-11刘明辉谢婷婷李丽娟李昌晓
刘明辉,谢婷婷,李 瑞,李丽娟,李昌晓,*
1 西南大学生命科学学院,三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室, 重庆 400715 2 西南山地生态循环农业国家级培育基地, 重庆 400715 3 西北农林科技大学人文社会发展学院, 杨凌 712100
三峡水库建成完工后,采取“冬蓄夏排”的水库调水方式,形成了垂直落差达30 m的大型水库消落带[1]。在这种特定的水文节律下,由于大部分原有植物不能忍受长时间高强度的反季节淹水胁迫[2],消落带原有植物大量消失、生物多样性降低、水土流失严重及生态系统退化等问题[3- 5]日益突出。为解决上述问题,进行三峡库区消落带的植被恢复重建是很有必要的[2]。研究表明池杉(Taxodiumascendens)具有耐水淹的特点,适合在高海拔的消落带及河岸带生长,是三峡库区消落带植被重建的优良树种之一[6- 7]。
池杉-土壤生态系统的关键特性是由碳(C)、氮(N)和磷(P)3种元素含量及其比率动态决定。池杉-土壤系统中,实生土壤对于池杉养分的供给,需要所有养分以一个合适的生态化学计量比存在时,生态系统才能健康、稳定的发展[8- 9]。池杉各构件的生态化学计量学在池杉-土壤生态系统尺度上平衡了多种元素[10]。同时,生态化学计量学已经成为当前生物地球化学循环与生态学研究的前沿热点之一,是生态系统生态学研究领域的新方向,能为池杉及其实生土壤C、N、P循环以及池杉-土壤互作的研究提供新思路。因此,在三峡库区这种特定的水文节律下,研究池杉-土壤系统的化学计量学特征是很有必要的。
在三峡库区淹水条件下,缺氧是池杉生存的主要限制因素[11]。它可能导致池杉的功能紊乱,能量代谢受阻,ATP合成不足,从而限制池杉对营养元素(尤其是C、N、P元素)的主动吸收与运输[12- 13],进而导致实生土壤中各种营养元素含量和有效性的改变。目前关于生态化学计量学的研究多集中在沉水植物[14- 15]和陆生植物[16],包括从全球、全国及地区尺度上对不同植物的不同器官[17]、不同林龄[18- 19]的C、N、P含量及其化学计量进行了研究。另外,对土壤养分化学计量特征的研究也较多[20- 22]。然而,在三峡库区消落带这种特定的水文节律变化下,将植物不同器官与实生土壤联系起来研究的却相对较少。因为植物-土壤系统作为陆地生态系统的重要组成部分,植物营养依赖土壤养分的供给,而植物又以枯落物的形式将营养元素C、N、P等养分逐步补偿给土壤[23]。此外,植物和土壤中C、N、P元素会因两个子系统中的循环而耦合[24]。关于土壤和植物C、N、P之间的关系,以及这些关系如何影响生态系统的功能和过程,人们知之甚少。为此提出以下科学问题:1) 三峡库区消落带水文节律下,池杉的C、N、P生态化学计量特征是否会由于生态内稳性而不受水淹胁迫的显著性影响?2) 池杉的生长是否受到N、P元素限制?因为生态化学计量学中的内稳性特征主要是指C、N、P等元素含量以及比值关系的稳定。当某种元素限制植物的生长时,植物可通过多种生理生化机制改变该元素的可利用性和利用效率,从而维持机体的养分含量以及相关性状的稳定[25]。因此,本研究选取三峡库区消落带的池杉-土壤系统为研究对象,研究池杉与实生土壤的C、N、P化学计量耦联特征,以深入探究池杉-土壤生态系统物质流动和能量循环及多元素的动态平衡关系。
1 材料与方法
1.1 试验地概况
本试验样地位于重庆市忠县石宝镇共和村汝溪河(长江一级支流)消落带植被修复示范基地(107°32′—108°14′E,30°03′—30°35′N),面积13.3 hm2,属于亚热带东南季风区山地气候。≥10℃年积温5787℃,年均温18.2℃,无霜期341 d,日照时数1327.5 h,日照率29%,太阳总辐射能83.7×4.18 kJ/cm2,年降雨量1200 mm,相对湿度80%,土壤主要为发育于亚热带地区石灰性紫色砂页岩母质的紫色土。示范基地原为弃耕梯田,2012年3月按照1 m×1 m的行间距,将2年生池杉幼苗种植于165—175 m海拔高程之间。池杉树苗栽种完成后立即进行浇水1次,并在同年6月中旬进行人工除草。
1.2 样品采集
于2018年7月在三峡库区消落带试验样地165—175 m海拔进行实地数据测定,并基于三峡库区现行水位调度节律进行165 m(深度水淹组,Deep Submergence,DS)、170 m(中度水淹组,Moderate Submergence,MS)和175 m(浅淹组,即对照,Shallow Submergence,SS)3个海拔的采样,不同水淹处理组5个水淹周期内的水淹深度和水淹持续时间见表1。取样时,池杉的生长状况见表2,对应的实生土壤性质见表3。

表1 不同水淹处理组5个水淹周期内的水淹深度和水淹持续时间
在每个海拔高程内分别随机选取5株长势相近且健康的池杉进行取样。叶片的采集是分别设置东、西、南、北4个方向采集每株植物中上部位健康成熟全展叶片,然后将所采下的叶片混匀后装入自封袋;由于考虑到试验样地的池杉栽种时间较短,用生长锥采集树干样品会对树木本身造成较大程度的损伤,因此本试验未采集树干样品;枝条的选择与叶片同步,选择生长状况良好的每株植株末端分枝5支,然后将其剪碎混匀装入自封袋;有研究表明,植物根系中最具生理活性的根区是细根(d≤2 mm),其对土壤养分及周围环境的变化比较敏感[26],根系的采集是在样方内以0.25 m半径等距离挖取池杉的细根,小心清理根系表面的土壤和杂质,最后将收集的根系混匀,四分法取部分装入自封袋;陆生植物对土壤的反馈作用会更多地被限制在表层土壤中[27],土壤的采集是去除表面枯枝落叶等杂物后,用平板利铲挖取对应植物的表层土壤(0—20 cm),采集的土样混合均匀,用四分法取足0.5 kg后密封带回。
1.3 指标测定
所有样品冷藏保存并立刻带回实验室,植物样品用自来水和超纯水清洗干净,并置于烘箱,先105℃杀青30 min,然后65℃烘干至恒重,将植物样品用莱驰MM400型球磨仪(Ball Mill, Germany)粉碎,用于测定C、N、P元素含量;土壤样品自然风干,碾磨后分别过1 mm和0.25 mm筛[28],用于测定土样含水量、容重、孔隙度、pH值、C、N、P元素含量等指标。
用测高杆测量池杉株高,用卷尺测量冠幅,用游标卡尺测量基径、胸径。采用土壤氧化还原电位计测定样地土壤温度和氧化还原电位(ORP),其中当ORP值介于+400—+700mV之间时土壤含氧量高且通气良好;淹水后ORP值变化从+400到+72mV;当ORP值低于+350mV,表明土壤氧气匮乏[13,29]。采用酸度计法(土∶水=1∶2.5)测定土壤pH值;采用环刀法测定土壤容重;采用烘干法测定土壤含水量;土壤孔隙度根据土壤容重按公式P=(|1-γ/ρ|)×100%计算,其中γ为土壤容重,ρ为土壤密度,采用“常用密度值”(2.65 g/cm3)[30]。所有样品的C和N元素含量采用元素分析仪(Elementar Vario EL, Germany) 测定,P元素含量先用微波消解仪(SpeedWave MWS- 4, Germany)进行消解,然后采用电感耦合等离子体发射光谱仪(ICP-OES, Thermo Fisher iCAP 6300, UK)测定。
1.4 数据处理
用Excel 2003和SPSS 22.0软件对测定的原始数据进行处理,采用单因素方差(One-way ANOVA)统计分析水淹处理对植物各组分C、N、P含量及化学计量比的影响,均采用Duncan多重比较(Duncan′s multiple range test)进行显著性检验,显著性水平设为α=0.05。采用回归分析计算植物与土壤之间的内稳定系数。因为4个轴中梯度最大值小于3,线性模型比较合适,采用Canoco 4.5软件进行冗余分析(Redundancy Analysis,RDA),并通过蒙特卡罗置换检验(Monte-Carlo permutation test)分析土壤理化性质对植物生态化学计量及生长指标变异解释度的显著性检验。所有图像均用Origin 9.0和CanoDraw for Windows 4.5软件制图。土壤和植物C、N、P元素浓度均以干重计(g/kg),所有C/N/P比值均以质量比计算。
内稳性指数根据内稳性模型[9]计算:y=cx1/H,y是植物不同器官的C、N、P元素含量及比值;x是环境中土壤的C、N、P元素含量及比值,c是常数。H是内稳性指数。1/H作为一种有用的工具,可以量化植物的化学计量稳态[31]。为方便统计,多用1/H来衡量内稳性的强弱[32]。
2 结果
2.1 池杉的生长特征参数
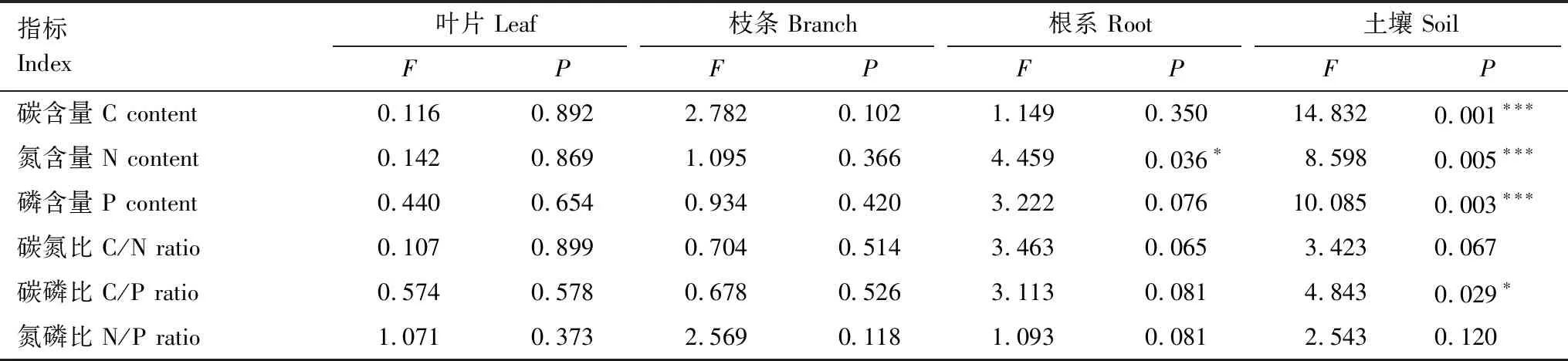
随着水淹时间和强度的增加,池杉的株高和冠幅均显著降低(P<0.05,表2)。基径的趋势为DS组 表2 不同水淹处理池杉的生长特征参数 表中的数据为平均值±标准误(n=5);不同小写字母表示植物在不同水淹处理间差异显著(P<0.05);初始值为2012年3月栽种时两年生池杉树苗的平均值(±标准误);DBH:胸径Diameter at breast height;DS:深度水淹组Deep Submergence;MS:中度水淹组Moderate Submergence;SS:浅淹组(对照)Shallow Submergence 随着水淹时间和强度的增加,池杉土壤温度均无显著性差异。土壤氧化还原电位趋势为DS组 单因素方差分析表明,水淹处理对池杉及实生土壤生态化学计量特征造成的影响不同(表4)。其中,池杉根系中的N元素受到水淹处理的显著影响(P<0.05);土壤中的C、N、P元素受到极显著影响(P<0.01),C/P比值受到显著影响(P<0.05)。 表3 不同水淹处理池杉的土壤理化性质 表中的数据为平均值±标准误(n=5);不同小写字母表示土壤指标在不同水淹处理间差异显著(P<0.05);ORP:氧化还原电位Oxidation reduction potential 表4 水淹处理对池杉及实生土壤生态化学计量特征的影响 ***P<0.01;*P<0.05 随着水淹时间和强度的增加,池杉各组分的C、N、P含量及其化学计量比表现出不同的变化规律及差异显著性(图1)。不同处理池杉枝条、叶片中的C、N、P及其比值无显著性差异,不受水淹的影响。根系组分中,P含量表现为DS组 另外同一水淹处理不同组分间的C、N、P含量表现出相似的趋势,总体来看,三个水淹处理组的N、P含量均表现为叶片>根系>枝条>土壤。除枝条P含量外,其他植物组分C、N、P含量均显著高于各自对应的土壤组分(P<0.05)。C/N比值和C/P比值表现出相似的趋势,均为枝条>根系>叶片>土壤。N/P比值均为枝条>叶片>根系>土壤,其中根系N/P比值显著高于土壤(P<0.05),但两者均显著低于叶片和枝条N/P比值(两组比较的P<0.05)。 图1 消落带不同水淹处理池杉-土壤系统各组分C、N、P含量及化学计量比Fig.1 The C, N and P concentrations and C∶N∶P stoichiometry in different components of T. ascendens and soil system under different submergence treatment 图中的数据为平均值±标准误(n=5);不同大写字母表示同一水淹处理不同组分间存在显著差异,不同小写字母表示不同水淹处理同一组分间存在显著差异(P<0.05) 池杉枝条、叶片、根系分别与实生土壤的内稳性系数不同,不同的器官内稳性存在差异(表5)。总体来看,池杉的稳定性较强。C元素稳定性相差不大,表现为叶片≈根系≈枝条;N元素稳定性表现为叶片>根系>枝条;P元素稳定性表现为叶片>枝条>根系;C/N比值的稳定性表现为根系>叶片>枝条;C/P比值的稳定性表现为枝条>叶片>根系,根系属于弱敏感型;N/P比值的稳定性表现为叶片>枝条>根系,根系属于弱敏感型。 池杉与实生土壤的C、N、P元素内稳性整体表现为P>C>N,比值内稳性表现为C/N>N/P>C/P,地上部分(枝条、叶片)C、N、P元素及其比值的稳定性较地下部分(根系)强。池杉地上和地下部分的元素间内稳性大小相反,地上部分内稳性为P>N,而地下部分内稳性为N>P。 冗余分析(RDA)表明土壤性质(C、N、P元素含量、温度、含水量、氧化还原电位、容重、空隙度、pH值)解释了数据总变异的74.7%,轴1和轴2的解释程度分别为36.1%和13.4%(图2)。池杉C、N、P含量与环境因子关系的累计解释量已达49.5%,并且轴1和轴2的物种与环境因子相关系数分别为0.945和0.763,表明池杉C、N、P元素含量及生长指标与土壤性质密切相关。RDA排序结果表明,土壤N含量、温度、容重、pH值与1轴均为正相关,土壤C含量、氧化还原电位与1轴相关性较小,其余土壤理化性质与1轴均为负相关;土壤C含量、土壤N含量、土壤P含量、含水率、pH值与2轴均为负相关,温度、氧化还原电位、孔隙度与2轴相关性较小。经过蒙特卡罗检验显示容重(F=2.893,P=0.008)、孔隙度(F=2.892,P=0.017)、氧化还原电位(F=2.694,P=0.031)、pH值(F=2.222,P=0.047)对池杉C、N、P元素含量的影响达到显著水平(P<0.05)。 表5 池杉与土壤之间的化学计量内稳性系数 Persson等[31]和Makin等[33]将1/H划分为五个程度,分别为(1)1/H≤0:强内稳态;(2)0<1/H≤0.25:内稳态;(3)0.25<1/H≤0.5:弱内稳态;(4)0.5<1/H≤0.75:弱敏感型;(5)1/H>0.75:敏感型 图2 池杉C、N、P含量及生长指标与土壤理化性质间的RDA二维排序图Fig.2 RDA two-dimensional sequence diagram of the relationship between C, N and P concentrations in different components of T. ascendens and growth indexes and soil physicochemical properties B-C:枝条碳含量Branch C;B-N:枝条氮含量Branch N;B-P:枝条磷含量Branch P;L-C:叶片碳含量Leaf C;L-N:叶片氮含量Leaf N;L-P:叶片磷含量Leaf P;R-C:根系碳含量Root C;R-N:根系氮含量Root N;R-P:根系磷含量Root P;P-H:株高Height;P-C:冠幅Canopy;P-BD:基径Basal diameter;P-DBH:胸径Diameter at breast height;C:土壤碳含量Soil C;N:土壤氮含量Soil N;P:土壤磷含量Soil P;Tem:温度Temperature;ORP:氧化还原电位Oxidation reduction potential;MC:含水率Moisture content;BD:容重Bulk density;pH:pH值pH value;Por:孔隙度Porosity 植物-土壤C、N、P三种元素之间的化学计量关系,在生态系统能量循环和物质流动及多元素的动态平衡中起着重要作用[34]。总体来讲,根系是吸收作用,枝条是运输作用,叶片是光合作用,而土壤将植物的不同器官联系起来,在根系等器官吸收土壤提供的养分同时,植物又以枯枝落叶等凋落物和根系分泌物的形式为土壤提供基质,将养分归还给土壤[35-38]。因此,对三峡库区消落带池杉-土壤系统的生态化学计量特征研究尤为重要。 池杉的生长受到了不同水淹时间和强度的影响。随着水淹时间和强度的增加,池杉的株高、冠幅、基径和胸径等生长指标均有降低的趋势,但与栽种时池杉相比长势较明显,表明经历6个水淹周期后三个水淹处理组的池杉在恢复期均生长较好,这与其稳定的化学计量比密不可分。 许多关于陆生植物化学计量关系的研究表明,N和P的化学计量关系受到许多因素的影响,如土壤肥力、温度、发育阶段等土壤性质有关[39-40]。RDA分析显示容重、孔隙度、氧化还原电位、pH值对池杉C、N、P含量的影响达到显著水平(P<0.05),其中土壤容重对其影响最大。因此,水淹胁迫越强,土壤中的O2含量就越少,这几项指标都与土壤中的含氧量有关,综合反映了O2含量对土壤中的C、N、P含量影响最大。有研究表明,在一定的温度范围内,土壤温度越高,植物的生长发育就越快。本研究中,土壤的表层温度介于26.92—27.34℃之间,处于合适的温度范围且SS组及MS组略大于DS组,适合植物生长。氧化还原电位表现为DS组 池杉C、N、P含量及其化学计量比的变化是由于植物的器官差异性,其含量及比值在各器官的分布规律证实了植物不同器官具有不同的生理机能与养分吸收和蓄积能力[42-43],也可能是归因于不同采样时间内植物物种营养元素吸收效率的变化和叶片等植物组分化学含量的变化[44-45]。本研究中,三个水淹处理组的池杉N、P含量表现为叶片>根系>枝条>土壤的趋势,这反映了池杉的不同组分对不同养分需求的相对大小。有研究表明,代谢活跃的组织或生长速率较快的器官含有较高比例的N和P含量[46-47],因为蛋白质(富含N的化合物)和核酸(富含P的化合物)的获得对生长至关重要[48-49]。总体来看,叶片和根系的N、P含量较高。其中叶片中最高,根系次之,枝条最低,说明7月为植株生长旺盛期,池杉吸收的N、P元素主要分配到叶片内,主要是因为树木叶片需要更多富含N和P的物质(如酶、转运蛋白和氨基酸)来参与代谢活动(如光合作用和呼吸作用)[50-51]。 植物叶片和细根是表征生态系统中环境因子和生物因子的代表,对周围环境的变化比较敏感[52-55],在功能上也联系的非常紧密。三个水淹处理组的池杉叶片C元素含量均无显著性差异,且远远大于土壤C含量,说明该消落带地区池杉叶片C含量及固C的能力可能相对较高,以及植物中的C元素主要来自大气,土壤C元素(含C化合物及有机质等)对其影响相对较小。本研究中DS组和MS组的池杉叶片N含量低于SS组,这是由于N元素含量是决定叶片光合作用强弱的主要因素之一,DS组池杉的光合作用受阻所致。池杉叶片P元素含量无显著性差异,SS组和MS组高于DS组,由于P元素对细胞分裂等生物生长至关重要,表明SS组及MS组池杉在消落期(7月)生长较好,与其生长指标随着海拔的升高相一致。 三个水淹处理组的池杉细根C含量与叶片趋势相似。N含量为SS组高于DS组和MS组,由于细根N含量在一定程度上反映了土壤肥力,细根N含量的减少可能是土壤N供应减少的结果[56]。DS组的P含量则最低,但是平均P元素含量远高于中国植物细根的平均P含量(0.954 g/kg)[57],且有研究表明亚热带东南季风区山地气候的样地普遍存在缺P的现象[58],这也是本研究结果差异存在的原因之一,同时还与土壤和植物C、N、P化学计量关系随着池杉种植年龄的增长变化有关。三个水淹处理组的池杉枝条中N、P含量无显著性差异且含量低于叶片与细根,与其自身生长节律及其结构特点有关。枝条中的N含量呈降低趋势,这是因为7月份为池杉生长旺盛期,大量的营养元素需要运输至叶片进行光合作用。 陆地生态系统中的生物经常受N和P限制[39],N/P比率用于表示生态系统的N限制或P限制。有研究表明,N/P比率小于14时表示受N限制,N/P比率大于16时表示受P限制[59-60]。在本实验中,池杉N/P的平均值较低,枝条为8.61,叶片为7.53,根系为4.44。这些值均远远低于N/P临界比率(<14),表明池杉的生长受到N元素限制,原因与池杉淹水-退水过程中,土壤中N元素含量下降比P元素含量更为明显有关。SS组和MS组的植物细根和叶片C/P比值和N/P比值均低于DS组,与Elser等[61]提出的“生长速率假说”一致,他们认为快速生长的细根和叶片会表现出更低的C/P比值和N/P比值。整体来看,池杉枝条中C/N、C/P比值无显著性差异但比值较高,N/P比值较低,主要是由于枝条中木质素等C含量较高的化合物稀释效应导致枝条中N含量较低,这在其他植物物种中也发现了类似的结果[46-47]。三个水淹处理组的细根C/N比值显著高于叶片,N/P比值显著低于叶片,主要是因为N、P元素较多地运输至叶片,来进行光合作用所致[62]。综上,本研究MS组对池杉生态化学计量特征影响较小,这对我们前面提出的科学问题“三峡库区消落带水文节律下,池杉的N、P生态化学计量特征是否会由于生态内稳性而不受水淹胁迫的显著性影响?”有了较好的解释。 对于生态系统的可持续性,土壤养分含量和土壤养分化学计量都很重要[63],它可以反映土壤的有机质组成以及质量程度等[64]。RDA分析中,轴1和轴2的物种与环境因子相关系数分别为0.945和0.763,表明池杉生态化学计量特征与土壤性质密切相关,植物体内C、N、P含量的变化会影响土壤养分含量和C∶N∶P化学计量比,因为植物通过凋落物和根际沉积为土壤提供了丰富的基质[65-66]。大多数植物营养物质受限于其在土壤中的可利用性[22,67],尤其是在热带森林中,植物C、N、P的动态主要受土壤P供应的影响[68]。DS组和SS组的土壤C、N、P含量差异不显著,均显著高于MS组,出现这样的原因可能与DS组和SS组存储养分的能力较好,出现富集等现象,也有可能是因为淹水期MS组侵蚀冲刷较大,导致营养元素流失,这有待于后续进一步观察研究。有研究表明,植被恢复年限较短(约12年之内)的生态系统通常缺乏大气来源的元素,特别是N元素,但富含岩石衍生元素,如P元素[69]。本研究中,C、N含量远远低于全球及中国陆地土壤平均C、N含量,P含量与我国的土壤平均值相当[70],主要原因可能与土壤中P的主要来源有关,因为P是岩石衍生元素,并且不能直接从土壤中大量获得[71];如果P不足会阻碍含P化合物(即DNA和ATP)的合成,从而限制植物的的生长[72]。 各水淹处理组的表层土壤元素比值维持相对稳定,这主要因为植物和土壤C、N、P元素之间的强烈耦合关系,使它们对环境的响应几乎是同步的。有研究表明,土壤C/N比值大于30表示硝酸盐淋溶风险较低;土壤C/N比值小于30表示硝酸盐淋溶风险较高[73]。在本研究中,三个水淹处理组的土壤C/N比值均远远小于30,可见三峡库区消落带土壤硝酸盐淋溶风险较高。土壤C/P比值通常被认为是土壤P矿化能力的标志,也是衡量微生物矿化土壤有机物质释放P或从环境中吸收固持P潜力的一种指标[74],较高的C/P比值是P有效性低的一个重要指标。P的浓度和有效性可以抑制含磷化合物(即DNA,RNA,ATP,NADPH和NADP+)的合成[72],这样的结果会改变光合作用、N固定和植物的其他生理过程[75- 76],最终导致植物生长的变化。土壤C/P比值为MS组 植物在长期水淹胁迫过程中逐渐形成了较强的适应能力,当外界环境发生变动时,有机体能保持其化学元素组成的稳定性而适应该水文环境[79],生态化学内稳性正是反映了当生物体对其周围环境作出反应时许多潜在生理和生化调整的结果。生态化学计量学稳定性的大小与植物的适应策略以及对环境适应性的强弱有关,在生态系统中植物群落的生态化学计量学稳定性越高所占的优势就越大。总体来看,池杉的生态化学计量学稳定性较强。这与Persson等[31]研究的一致,他们认为内稳性较强的植物,养分利用方式较为保守,在多变贫瘠的环境中也能维持机体的生长。池杉枝条、叶片、根系与实生土壤的C元素稳定性较好,表明池杉体内的C元素充足,尽管C元素的来源有大气中的CO2,但是从生态化学计量学的角度来看,其植物体内的C元素仍然遵循生态内稳性理论。在三峡库区淹水的条件下,当O2的供应受到限制时会发生反硝化作用,反硝化作用使硝酸盐还原成N2和N2O,从而降低了土壤中氮素营养的含量[80-81]。P 元素稳定性大于N元素,这与Yu等[40]和庾强[82]的研究结果不一致,可能是由于池杉在生长旺盛期利用N元素来进行光合作用等活动。 池杉细根是主要与土壤进行接触并进行养分交换的器官[83],所以植物的细根会很快感应到土壤环境的变化。与粗根相比,细根结构更为复杂,具有较高的生理活性[84],且在贫瘠多变的土地上,大部分植物细根集中在表层土壤中。本研究中,根系的稳定性较低,C/P比值和N/P比值甚至出现弱敏感型,这可能与细根寿命较短,以及距离同化器官较远等有关。幼嫩细根的生长首先出现于根原基,未成熟的细胞体积增大并分裂,细胞中的酶系统非常活跃,呼吸和代谢作用等活动加强,很快变成成熟的细根,使其处于不断更新的状态。而在细根整个代谢过程中,P元素对细胞分裂等生物生长至关重要,因此会导致细根部分的P元素变动或者移动性比较强,出现敏感型。由此说明,在消落带植被修复重建时应尽可能地减少对池杉细根的损伤,以此来减少元素的损失。此外,三峡水库在退水期间会带走大量的凋落物,从而使该区域土壤营养元素流失,因此应合理利用凋落物的分解来维持该区域的养分平衡。尽管如此,但池杉幼林仍在三峡库区这种特定的水文节律下生长良好,这证明其仍然遵循生态内稳性理论,这种独特的机制使池杉能够适应环境,并加强了我们对三峡库区消落带地区碳氮磷化学计量学的理解。 地上部分C、N、P元素及其比值的稳定性整体较地下部分强,与Garrish等[85]、Minden和Kleyer[86]、Schreeg等[87]研究灌木植物和乔木幼苗叶片的化学计量内稳性高于根系基本一致,反映出其保持自身体内化学组成相对恒定的能力,这极有可能是池杉适应三峡消落带的一种生长策略,即在养分限制的条件下,池杉对其生长过程中地上与地下部分器官养分分配的平衡,通过内稳性较低的地下部分(根系)对养分的吸收、贮存来维持地上部分(枝条、叶片)较高的内稳性[40],从而使植物能够稳定、健康生长,以应对接下来的水淹逆境,使其在变化的环境中占有优势,最终成为三峡库区消落带适宜生长的优势树种。 本研究通过利用原位实验来评价三峡库区消落带恢复期适生木本植物池杉及其实生土壤C、N、P含量及化学计量比的特征,发现在经历了6个水文节律周期后,水淹是影响池杉生长的主要因素,对池杉生态化学计量特征影响较小;池杉各组分能够对水位变化做出积极的响应,保持稳定的生态化学计量特征,能够有效权衡池杉各个器官中元素的含量配比。RDA分析表明池杉生态化学计量特征及生长指标与土壤性质密切相关。总体来看,池杉的生态化学稳定性较强,地上部分C、N、P元素及其比值的稳定性较地下部分强。池杉在多变的消落带水文环境中能够维持体内的化学计量平衡以响应水淹,是三峡库区消落带适宜生长的优势树种。此外,本研究区域可能存在N元素的限制,可考虑拦截利用凋落物的分解积累土壤养分来改变该区域的生态问题,以维持整个生态系统的稳定和健康。
2.2 池杉的土壤理化性质
2.3 池杉各组分C、N、P含量及化学计量比的变化特征


2.4 池杉生态化学计量内稳性差异
2.5 池杉C、N、P含量及生长指标与土壤理化性质间的冗余分析


3 讨论
3.1 不同水淹处理对池杉生长状况及其实生土壤基本特征的影响
3.2 不同水淹处理对池杉不同器官C、N、P含量及化学计量比变化特征的影响
3.3 不同水淹处理对池杉实生土壤C、N、P含量及化学计量比变化特征的影响
3.4 池杉生态化学计量内稳性特征
4 结论
