林麝化脓病病原菌化脓隐秘杆菌与铜绿假单胞菌互作机制研究
2020-06-11袁阳李静张爱雪林家富褚以文王欣荣赵克雷
袁阳, 李静, 张爱雪, 林家富, 褚以文, 王欣荣, 赵克雷
(抗生素研究与再评价四川省重点实验室,四川抗菌素工业研究所,成都大学,成都610052)
林麝Moschusberezovskii是一种重要的经济野生动物资源,主要分布在我国西南地区。雄性林麝产生的麝香既是药用价值较高的中药材,也是名贵香料的主要原料之一。由于人为捕杀以及栖息地的缩减,林麝被列为国家Ⅰ级重点保护野生动物,林麝人工养殖获取麝香可以大大缓解市场对麝香不断增加的需求。我国人工养麝可以追溯到20世纪50年代,但到目前为止仍然没有形成大规模养殖,其中化脓性疾病是重要制约因素之一(罗燕等,2006;赵克雷等,2011)。
近年研究发现,化脓隐秘杆菌Trueperellapyogenes是导致林麝化脓病的原发病原菌,在林麝化脓性病灶中检出率达到100%(Zhaoetal.,2011)。化脓隐秘杆菌是一种革兰氏阳性条件致病菌,可以导致多种反刍动物多器官与组织的化脓性病变。虽然化脓隐秘杆菌是引起林麝化脓性病变的主要病原菌,但是在濒临死亡或者已经死亡林麝的化脓性病灶中往往能检测到大量的铜绿假单胞菌Pseudomonasaeruginosa、大肠杆菌Escherichiacoli等病原菌(赵克雷等,2011;Zhaoetal.,2019a)。研究发现,病原菌之间存在多种物理或化学的交流方式,可以联合各病原菌种群共同促进疾病的进程,导致治疗变得更棘手(Shortetal.,2014;Pragmanetal.,2016;袁阳等,2019)。在林麝化脓病发病过程中,化脓隐秘杆菌作为原发致病菌通常不会直接导致宿主死亡,而机体免疫力下降引起铜绿假单胞菌等强毒致病菌的继发性感染可能是导致林麝死亡的根本原因(Jost & Billington,2005;Zhaoetal.,2011,2013)。
与其他常见病原菌相比,铜绿假单胞菌具有相对较大的基因组和较多的基因,以灵活多变的胞内调控网络适应多种生存条件,同时还具有复杂而庞大的群体感应(quorum-sensing,QS)网络调控系统,在营养获取、毒力发挥和种间竞争中发挥着关键作用(Stoveretal.,2000;Balasubramanianetal.,2013;Valentinietal.,2018;Zhaoetal.,2018,2019a)。铜绿假单胞菌的QS系统由3个层级排列的次级调控系统(las、rhl和pqs)组成,其中las系统处于顶端调控位置(Dandekar & Greenberg,2013)。las和rhl系统具有完整的信号分子合成蛋白(LasI/RhlI)和调控蛋白(LasR/RhlR),属于自诱导调控系统,且rhl系统的表达激活主要受las系统调控。pqs系统则只具有调控蛋白PqsR(MvfR),其表达激活需要接收其他路径(LasR和RhlR相关)产生的信号分子Pseudomonasquinolone signal(PQS),进而调控部分受LasR和RhlR控制的基因的表达,属于外源诱导调控系统(Choietal.,2011;Balasubramanianetal.,2013;Dandekar & Greenberg,2013)。Zhao等(2017)研究发现,铜绿假单胞菌的QS信号分子可以抑制化脓隐秘杆菌的生长,而Huang等(2018)进一步研究证实了该过程同样发生在体内感染。这些研究结果暗示,林麝化脓病病灶中出现的优势菌替换现象可能与铜绿假单胞菌利用QS系统抑制化脓隐秘杆菌的生长有关。本研究通过构建铜绿假单胞菌多种QS突变菌株,利用平板互作模型详细研究了化脓隐秘杆菌与铜绿假单胞菌的种间关系,可为林麝化脓病的诊治方案和抗菌药物开发、改进提供参考。
1 材料
1.1 菌株来源
野生型铜绿假单胞菌模式菌株PAO1(Zhaoetal.,2018)、rhlR基因敲除菌株PAO1-ΔrhlR(Zhaoetal.,2019b)和pqsR基因敲除菌株PAO1-ΔpqsR(Jacobsetal.,2003)为实验室保存。化脓隐秘杆菌TP13早期分离自林麝肺脏脓液(Zhaoetal.,2013)。
1.2 培养基
LB肉汤培养基、LB营养琼脂培养基、金氏B培养基(Zhaoetal.,2019b)。
1.3 主要试剂和仪器
庆大霉素、磷酸盐缓冲液(PBS)、PCR(Bio-Rad)、琼脂糖凝胶电泳仪(Bio-Rad)、UV5500PC型紫外可见分光光度计(上海元析仪器有限公司)。
2 方法
2.1 铜绿假单胞菌群体感应基因敲除菌株构建
参照课题组前期已建立的铜绿假单胞菌基因敲除方法(Zhaoetal.,2019b),利用基因同源重组和转化接合对野生型铜绿假单胞菌PAO1、PAO1-ΔrhlR、PAO1-ΔpqsR菌株进行lasR基因敲除,获得PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔlasRpqsR菌株。所用引物如表1所示。

表1 基因敲除所使用的引物Table 1 Primers used for gene knock-out
以PAO1基因组DNA为模板,用引物lasR1/lasR2、lasR5/lasR6分别扩增出前同源臂(496 bp)与后同源臂(577 bp);以实验室保存的pRU1103质粒为模板,用引物GM-lasR3/GM-lasR4扩增出庆大霉素抗性基因元件(917 bp)。这3个片段通过Overlap PCR连接成一条1 990 bp片段,目的是用庆大霉素抗性基因去替换lasR基因的564 bp片段。利用限制性内切酶SacI-XbaI双酶切Overlap PCR产物以及质粒pCVD442构建pCVD442-KOlasR重组质粒。将该重组质粒转入E.coliS17中,随后将带有pCVD442-KOlasR的E.coliS17与PAO1、PAO1-ΔrhlR、PAO1-ΔpqsR进行转化接合,使E.coliS17中的pCVD442-KOlasR重组质粒进入受体菌中。利用含有15 μg·mL-1庆大霉素的金氏B平板筛选阳性克隆(铜绿假单胞菌在金氏B培养基上会显示出明显的荧光绿);扩大培养阳性克隆,提取基因组DNA后用引物lasR1/lasR6进行PCR扩增,以PAO1基因组作为阴性对照,扩增结果送成都擎科梓熙生物技术有限公司测序验证。
2.2 病原菌平板距离培养实验
挑取化脓隐秘杆菌、PAO1、PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔrhlR、PAO1-ΔpqsR、PAO1-ΔlasRpqsR单克隆于5 mL LB肉汤培养基中,37 ℃,220 r·min-112~16 h,化脓隐秘杆菌由于生长较慢,培养24 h。1 2000 r·min-15 min后收集菌体,并用PBS冲洗菌体3次。然后用PBS调节菌液OD600=1作为母液备用。
参照课题组前期已建立的多病原菌平板互作研究方法(Zhaoetal.,2018),将不同基因型铜绿假单胞菌菌株和化脓隐秘杆菌成对接种于LB平板,同种菌株间距1 cm,不同菌株的间距依次增加(初始间距为0.5 cm,依次增加0.5 cm),每个平板上设置6个梯度,每个点的接菌量均为2 μL母液,将平板置于37 ℃培养1~4 d,每个实验设置3个独立重复。每隔1 d,使用无菌竹铲挖出化脓隐秘杆菌与 PAO1、PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔrhlR、PAO1-ΔpqsR、PAO1-ΔlasRpqsR平板互作的近距离培养和远距离培养的菌落至1 mL PBS中,用移液器反复吹打菌体进行充分分散、冲洗后,用紫外分光光度计在波长600 nm下测量光密度。
2.3 基于胞外产物作用的平板互作实验
为进一步确认PAO1、PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔrhlR、PAO1-ΔpqsR、PAO1-ΔlasRpqsR的胞外产物对化脓隐秘杆菌的抑制作用,基于上述平板距离培养实验方法,先在LB平板的左侧分别只接种PAO1、PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔrhlR、PAO1-ΔpqsR或PAO1-ΔlasRpqsR,培养2 d后移除菌落,然后在右侧依次接种化脓隐秘杆菌(与左侧原接种铜绿假单胞菌菌落标记初始间距为0.5 cm,依次增加0.5 cm)。
3 结果
3.1 基因敲除菌株的获得
以野生型铜绿假单胞菌PAO1基因组DNA为模板,分别扩增出横跨lasR基因上游496 bp序列(前同源臂)与下游577 bp序列(后同源臂),并以pRU1103质粒为模板扩增出庆大霉素抗性基因元件(917 bp),胶回收后用Overlap PCR进行片段连接与扩增。琼脂糖凝胶电泳结果显示,3条片段连接后的产物大小为1 990 bp,符合预期(图1:A),并且DNA测序结果无误。将片段用lasR1/lasR6引物进行富集扩增,胶回收产物经双酶切后连接穿梭质粒pCVD442,转化E.coliS17感受态后于庆大霉素抗性平板筛选获得多个阳性菌株,提取质粒后验证插入片段测序无误,获得E.coliS17 pCVD442-KOlasR重组菌株。以E.coliS17 pCVD442-KOlasR为供体菌,以PAO1、PAO1-ΔrhlR、PAO1-ΔpqsR为受体菌进行转化接合实验后,于庆大霉素抗性的金氏B平板筛选获得多个产荧光绿的阳性克隆;以野生型铜绿假单胞菌PAO1为对照,用lasR1/lasR6引物进行扩增,如未敲除,则电泳条带大小为1 637 bp,如lasR基因已被庆大霉素抗性基因元件替换,则扩增后电泳条带大小为1 990 bp(图1:B)。挑选1 990 bp条带对应的菌株用lasR1/lasR6引物测序验证,并对菌株进行16S rDNA扩增测序后鉴定物种,最终获得PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔlasRpqsR突变菌株。

图1 不同铜绿假单胞菌菌株lasR基因的敲除Fig. 1 Knock-out of lasR gene from different Pseudomonas aeruginosa strains
A. 前、后同源臂和庆大霉素抗性元件经Overlap PCR拼接后的产物; B.lasR基因敲除菌株的筛选; 分子量标记: DL2000, 1. 空白对照,2、3. 野生型对照
A. Ligation of the upstream and downstream fragments and gentamycin resistance cassette by Overlap PCR; B. Selection oflasRmutant strains; Marker: DL2000, Lane 1. blank, Lanes 2 and 3. wild-type control
3.2 铜绿假单胞菌QS突变菌株与化脓隐秘杆菌的种间互作
铜绿假单胞菌菌落近端的化脓隐秘杆菌菌落均不同程度地小于远端的菌落,而铜绿假单胞菌的生长则没有显著变化(图2)。不同的铜绿假单胞菌QS突变菌株均对化脓隐秘杆菌的生长表现出一定的抑制作用,且抑制效果随着培养时间的延长而更明显(图3)。

图2 不同基因型的铜绿假单胞菌与化脓隐秘杆菌平板互作表型实验Fig.2 Proximity assay of different genotypes of Pseudomonas aeruginosa (PA) with Trueperella pyogenes (TP)

图3 平板互作表型实验中化脓隐秘杆菌的菌落规模Fig.3 Colony sizes of Trueperella pyogenes in the proximity assay
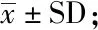

对上述平板互作中近端化脓隐秘杆菌的菌落规模进行横向比较发现,不同铜绿假单胞菌群体突变菌株对邻近化脓隐秘杆菌的生长有不同程度的抑制作用,尤其从第2天开始,各突变菌株对化脓隐秘杆菌的抑制效果均弱于野生型铜绿假单胞菌菌株。与野生型菌株相比,只有将铜绿假单胞菌QS调控系统中的lasR和rhlR基因同时敲除后,化脓隐秘杆菌的菌落才有较大程度回升(图4)。

图4 不同铜绿假单胞菌群体感应突变菌株对近端化脓隐秘杆菌的抑制作用对比Fig.4 Comparison of the inhibitory effect of different Pseudomonas aeruginosa (PA) strains on the growth of proximal Trueperella pyogenes colonies
3.3 铜绿假单胞菌胞外产物对化脓隐秘杆菌的生长抑制
将铜绿假单胞菌不同突变菌株分别成排接种于平板培养2 d后,将菌落完整挖去,然后将化脓隐秘杆菌按距离接种于铜绿假单胞菌分泌到培养基内的胞外产物上,继续培养1 d。结果发现,铜绿假单胞菌不同QS系统突变菌株的胞外产物对化脓隐秘杆菌的生长具有不同程度的抑制作用,以野生型菌株PAO1和lasR-pqsR双突变菌株的抑制效果最明显,而lasR-rhlR双突变菌株的胞外产物对化脓隐秘杆菌的抑制效果最弱(图5)。
4 讨论
近年来研究发现较多的慢性感染都是由多种病原微生物共同定殖引起,这些病原菌通过复杂的种间相互作用共同抵御宿主免疫和抗生素的清除作用。因此,探索病原菌之间的相互作用对疾病的诊断和治疗具有重要意义(Littleetal.,2008;Petersetal.,2012;Orazi & O’Toole,2019)。化脓隐秘杆菌是林麝化脓病的主要诱发菌,也是初期感染病灶中的优势菌。随着感染的持续恶化,林麝脓肿病灶中能够检测到大量铜绿假单胞菌(赵克雷等,2011)。铜绿假单胞菌是一种具有较强致病能力的条件病原菌,这很大程度上归因于其具有一套由3个层级排列的las、rhl和pqs调控系统组成的QS系统。这3个系统共同控制着绝大多数胞外物质的产生,也赋予了铜绿假单胞菌对其他细菌的竞争优势(Schusteretal.,2003;Choietal.,2011;Balasubramanianetal.,2013;Guoetal.,2014;袁阳等,2019;Zhaoetal.,2019a)。本研究就林麝化脓病发病过程中出现的化脓隐秘杆菌与铜绿假单胞菌的优势菌替换现象,围绕铜绿假单胞菌的QS系统在这2种菌之间的种间互作展开研究。

图5 不同铜绿假单胞菌群体感应突变菌株胞外产物对化脓隐秘杆菌的生长抑制作用Fig.5 Inhibitory effects of the extracellular products of different Pseudomonas aeruginosa (PA) strains on the growth of Trueperella pyogenes (TP) colonies
利用平板距离培养实验发现,靠近PAO1、PAO1-ΔlasR、PAO1-ΔlasRrhlR、PAO1-ΔrhlR、PAO1-ΔpqsR、PAO1-ΔlasRpqsR生长的化脓隐秘杆菌均被抑制,而lasR-rhlR双敲除菌株对化脓隐秘杆菌的抑制作用明显减弱。这些发现与其他研究发现的铜绿假单胞菌las-和rhl-QS系统产生的2种高丝氨酸类酯3-oxo-C12-HSL和C4-HSL信号分子均能抑制化脓隐秘杆菌的生长结果相一致(Zhaoetal.,2013;Huangetal.,2018)。从平板互作表型来看,虽然某些基因缺陷型铜绿假单胞菌胁迫下的近端化脓隐秘杆菌菌落大小只略小于远端,但肉眼无法确定菌落厚度、密度等信息,尤其是某些革兰氏阳性菌(如金黄色葡萄球菌Staphylococcusaureus)会通过产生小菌落变异体、提高生物膜产量等来应对铜绿假单胞菌的胁迫(Biswasetal.,2009)。本研究中光密度精确测定结果与表型的差异反映出同为革兰氏阳性菌的化脓隐秘杆菌可能也存在类似的压力反应机制。其他研究发现,铜绿假单胞菌QS系统产生的其他毒力因子如弹性蛋白酶、鼠李糖脂、PQS、绿脓菌素等对大肠杆菌、金黄色葡萄球菌的生长代谢均有抑制作用,这些毒力因子的表达激活受las-、rhl-和pqs-QS系统共同调控,可通过抑制附近其他病原菌的有氧呼吸和膜系统合成使铜绿假单胞菌获得显著的竞争优势(Baronetal.,1989;Guoetal.,2014;Khare & Tavazoie,2015;Hotterbeekxetal.,2017;Tognonetal.,2019)。这也为本研究中发现的铜绿假单胞菌不同QS系统缺陷菌株均能对化脓隐秘杆菌表现出不同的抑制作用提供了解释。这些结果提示针对铜绿假单胞菌QS系统信号分子进行结构类似物筛选,获得可以同时抑制铜绿假单胞菌QS系统和抑制化脓隐秘杆菌生长的小分子药物,可能是攻克林麝化脓病的新策略和新方向。
