尼罗罗非鱼p38MAPK基因克隆与表达分析
2020-06-09马银花邹芝英祝璟琳李大宇胡平各陈炳霖
马银花,邹芝英,祝璟琳,喻 杰,肖 炜,李大宇,杨 弘,胡平各,陈炳霖
(1.南京农业大学无锡渔业学院,江苏无锡 214081;2.中国水产科学研究院淡水渔业研究中心,农业农村部淡水渔业和种质资源利用重点实验室,江苏无锡 214081)
丝裂酶原活化蛋白激酶(mitogen-activated protein kinase,MAPK)是一类丝氨酸/苏氨酸蛋白激酶,能够通过磷酸化级联反应将细胞外的刺激信号带入细胞核内[1]。p38MAPK是MAPK家族中的重要组成部分,位于细胞众多信号转导通路的中枢位置,在炎症反应[2]、细胞应激、凋亡[3]、细胞周期[4]和生长等生理、病理过程中起重要作用。
p38MAPK基因包括四种亚型:p38α、p38β、p38γ、p38δ,其中p38α和p38β几乎存在所有组织中,p38γ和p38δ只在肌肉、胰腺、肠和肾等组织中表达[1]。此外,p38MAPK家族的所有成员均具有Thr-Gly-Tyr(TGY)双磷酸化位点、Ala-Thr-Arg-Trp(ATRW)底物结合位点。在MAPK级联反应中,MAPK激酶(MAPKK)可以通过磷酸化TGY位点,从而活化p38MAPK,启动下游基因的表达,进而发挥调节作用[5]。研究表明,p38MAPK能被多种细胞外刺激所激活,如炎症细胞因子、环境胁迫、病原体感染等。当其被激活时,能介导中性粒细胞的活化,促进巨噬细胞产生IL-1、IL-6、TNF-α等炎症因子,参与机体免疫反应[6],帮助机体抵御病害威胁。Barruet等[7]曾用脂多糖(LPS)刺激CD14+原代单核细胞,结果显示p38MAPK表达量上调,细胞因子、趋化因子和转化生长因子-β(TGF-β)分泌增加。可见,p38MAPK在机体免疫系统中发挥着重要作用。目前在大西洋鲑[8]、斑马鱼[9]、鲤[10]、文昌鱼[11]等水产动物中都克隆出p38MAPK基因,但关于罗非鱼p38MAPK的研究很有限。
罗非鱼(Oreochromisspp.)作为联合国粮农组织(FAO)重点推广的养殖对象,被誉为未来动物性蛋白质主要来源之一,具有较高的经济价值[12]。然而近年来罗非鱼链球菌病频繁暴发,使罗非鱼养殖产业的经济损失日趋严重。其中,无乳链球菌(Streptococcusagalactiae)是主要致病菌,能造成罗非鱼全身性组织炎症反应和器官损害,尤其是肝脏、脾脏、肾脏和脑等重要器官功能障碍和衰竭,严重时甚至引起鱼类的急性死亡[13]。因此,为了研究p38MAPK与罗非鱼的免疫相关性,本实验克隆获得尼罗罗非鱼p38MAPK基因(ntp38MAPK)cDNA序列,初步分析了其在尼罗罗非鱼不同组织以及感染无乳链球菌后不同时间点的表达变化规律,以期能为应对链球菌刺激后的调控机理提供一定的基础数据,为罗非鱼的疾病综合防控提供理论基础,进而为罗非鱼养殖健康可持续发展提供科学依据。
1 材料与方法
1.1 实验材料
实验所用的尼罗罗非鱼埃及品系NE来自中国水产科学研究院淡水渔业研究中心农业部罗非鱼遗传育种中心。实验用鱼在水温28 ℃的循环水箱中暂养一周后使用。
无乳链球菌(LB110808-2)保存于本实验室-80 ℃超低温冰箱[14]。该病原菌已经通过API20 Strep快速鉴定及16S rDNA分子鉴定确定为无乳链球菌,回归感染证明其具有较强毒力。
1.2 实验方法
1.2.1 总RNA的提取及cDNA第一链的合成
总RNA的提取采用Trizol 试剂(Invitrogen),按照说明书进行操作,通过琼脂糖凝胶电泳与紫外分光光度计确定其完整性及质量。利用反转录试剂盒(TaKaRa)合成cDNA,利用SMARTTM RACE Amplification 试剂盒(Clontech)合成3′RACE和5′RACE cDNA第一条链。
1.2.2 尼罗罗非鱼p38MAPK基因的cDNA克隆
根据NCBI中罗非鱼p38MAPK转录组预测序列(XM_003441527),用Primer 5.0软件设计特异性引物(表1),将引物送至上海生工生物工程有限公司进行合成。
以罗非鱼肝脏组织 cDNA为模板进行中间片段PCR扩增,反应体系(20 μL):rTaq 0.2 μL,10×PCR Buffer 2 μL,dNTP Mixture 2 μL,5 μmol/L的p38-F1引物1.6 μL,5 μmol/L的p38-R1引物1.6 μL,cDNA模板1 μL,RNase-free Water添加至20 μL。PCR反应程序:95 ℃预变性5 min;94 ℃变性45 s,55 ℃退火50 s,72 ℃延伸1 min,35个循环;最后在72 ℃再延伸10 min。PCR扩增后使用10 g/L的琼脂糖凝胶电泳检测PCR目的片段产物的大小。将目的片段与pMD19-T载体(TaKaRa)连接,转入到E.coil感受态细胞(TaKaRa)中,铺板于氨苄-LB培养基上培养。PCR检测阳性菌落,送至生工生物(上海)工程股份有限公司进行测序。在NCBI中进行比对,确定p38MAPK基因片段。结合RACE技术获得p38MAPK基因的cDNA全长,具体操作详见说明书。所用引物见表1。

表1 实验所用引物Tab.1 The primers used in this study
1.2.3 序列分析
利用Emboss(http://emboss.bioinformatics.nl/cgi-bin/emboss/showorf)软件预测氨基酸序列;ProtParam(https://web.expasy.org/cgi-bin/protparam/ protparam)软件对推导出的蛋白序列进行蛋白理化特性预测;磷酸化位点预测使用NetPhos(http://www.cbs.dtu.dk/services/NetPhos)软件;用NCBI中的Blast程序搜索用于p38MAPK基因同源性分析与系统树构建的氨基酸序列;采用Clustal X软件进行多重序列比对;利用MEGA 5.05软件构建NJ系统进化树;利用GOR方法(http://npsa-pbil.ibcp.fr/cgi-bin gor4)进行二级结构预测。
1.2.4ntp38MAPK基因的组织表达分析
选6尾二龄的健康尼罗罗非鱼,选取肝、脾、头肾、体肾、脑、血液、肌肉、鳃、肠、精巢、心脏等11种组织,放入Trizol中,保存在液氮中,用于ntp38MAPK基因的组织表达分析。
根据ntp38MAPK基因的全长序列设计荧光定量PCR引物p38-F4、p38-R4(表1),以β-actin为内参基因(表1),不同组织cDNA为模板,进行荧光定量PCR。反应体系20 μL,包括SYBR®Green PCR Master Mix (TOYOBO)10 μL,0.8 μL 引物(5 μmol/L),2 μL cDNA模板(1 μg),RNase-free Water 补足到20 μL。反应程序为95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火20 s,72 ℃延伸40 s,40个循环;融解曲线分析。荧光定量PCR检测结果采用2-ΔΔct法进行分析。
1.2.5ntp38MAPK基因链球菌感染后的表达分析
将经鉴定的无乳链球菌原种接种并复壮,划线培养。实验时,将菌种接种于脑心浸液液体培养基(BHI)中,210 r/min、29 ℃的条件下扩大培养24 h,用无菌的8.5 g/L NaCl溶液洗涤后配置成浓度为6.9×108cfu/mL的菌悬液原液[14]。感染浓度通过连续10倍稀释后涂布于脑心浸液琼脂平板,28 ℃培养24 h后,计数单克隆菌落倒推计算所得。
选取尼罗罗非鱼埃及品系(NE)90尾,体质量(182±30)g,于循环养殖系统适应一周,每天适当提高温度,实验前使水温达到(33±1)℃。实验开始,对实验鱼腹腔注射浓度为1.4×105cfu/mL的无乳链球菌,每尾0.3 mL,分别于攻毒后0、2、4、8、12、24、48、72 h共8个时间点进行取样(0 h设为对照组),每个时间点取6尾鱼,每尾鱼取头肾、肝、脾组织,保存于液氮中用于RNA的提取。人工感染3 d后随机取濒死的罗非鱼解剖,无菌操作取脑和肾脏组织在血平板上分离、纯化病原菌,并用梅里埃API 20 Strep试剂条(Biomerieox)进行快速鉴定。
按照前述方法提取攻毒后各个时间点尼罗罗非鱼肝、脾、头肾组织的RNA,逆转录成cDNA。以cDNA为模板,β-actin为内参进行荧光定量PCR。反应体系、反应程序以及数据处理方法同上组织表达。
1.2.6 统计学分析
实验数据用统计学软件SPSS 19.0的单因素方差(ANOVA)分析和Duncan′s多重比较进行组间比较分析,当P<0.05时认为差异显著。实验数据用平均值±标准误(Mean±SE)表示。
2 结果与分析
2.1 ntp38MAPK基因cDNA的克隆及序列分析
通过RACE技术克隆得到尼罗罗非鱼p38MAPK基因cDNA全长序列,命名为ntp38MAPK(GenBank登录号:MN478475)。ntp38MAPK的cDNA全长1 789 bp,ORF长1 086 bp,5′UTR长342 bp,3′UTR为361 bp。3′端含有polyA尾,不含多聚腺苷酸AATAA加尾信号。ORF可编码361个氨基酸,预测分子量为41.598 kD,理论等电点为5.1。本序列包括35个磷酸化位点(其中丝氨酸位点14个,苏氨酸位点13个,酪氨酸位点8个),同时含有p38家族典型的Thr-Gly-Tyr(TGY)双磷酸化位点、紧密相连的底物结合位点Ala-Thr-Arg-Trp(ATRW)以及CD结构域(图1)。利用GOR法对蛋白质二级结构预测后发现,整条氨基酸序列的二级结构有 α 螺旋、延伸链以及无规则卷曲构成,其中α螺旋占37.95%,延伸链占14.40%,无规则卷曲比列最大占47.65%。
2.2 同源性分析
经NCBI中BLAST比对分析发现,尼罗罗非鱼p38MAPK蛋白与其他物种的具有较高的相似性。利用Clustal-X软件进行多重序列比对,结果表明:该序列与大黄鱼(Larimichthyscrocea:XP_010754403.1)、鲈(Dicentrarchuslabrax:CBN8089 3.1)的相似性最高,达到97%。与其他脊索动物的相似性:与斑马鱼(Daniorerio:AAQ91248.1)的相似性为94%;大西洋鲑(Salmosalar:XP_013991178.1)的相似性为93%;与牙鲆(Paralichthysolivaceus:XP_019942433.1)、虹鳟(Oncorhynchusmykiss:XP_021472540.1)以及牦牛(Bosmutus:QAB47435.1)的相似性都为92%;与人(Homosapiens:NP_002742.3)和小鼠(Musmusculus:AAH12235.1)的相似性都为90%。此外与其他节肢动物的相似性也很高,与拟穴青蟹(Scyllaparamamosain:AHH29322.1)的相似性为76%;与蝇蛹金小蜂(Nasoniavitripennis:NP_001136337.1)的相似性为75%;与家蚕(Bombyxmori:NP_001036996.1)、棉铃虫(Helicoverpaarmigera:AHL46461.1)的相似性都为73%(图1)。
利用MEGA 6.0软件,采用NJ法构建了MAPK系统进化树(图2)。从系统进化树可以看出,尼罗罗非鱼的p38MAPK与脊索动物大黄鱼、鲈的p38MAPK聚为一支,后与大西洋鲑、虹鳟聚为一支,最后与其他节肢动物聚合。

图1 ntp38MAPK的氨基酸序列与其他物种的氨基酸序列比对Fig.1 Comparison of amino acid sequence of ntp38MAPK with multiple amino acid sequences of other species双磷酸化位点TGY、底物结合位点ATRW和CD结构域用方框标出

图2 利用MEGA5.05软件构建的基于ntp38MAPK氨基酸序列的NJ系统进化树Fig.2 NJ phylogenetic tree based on ntp38MAPK amino acid sequence constructed with MEGA5.05 software尼罗罗非鱼ntp38MAPK用星号标出
2.3 ntp38MAPK基因的组织表达分析
如图3所示,ntp38MAPK在心脏、肝脏、脾、头肾、体肾、脑、肌肉、肠、鳃、血液以及精巢中都有表达,且存在明显的组织差异性。其中肌肉组织中表达量最高(为头肾中相对表达量的6.9倍),随后是心脏、肝脏、血液以及脑,精巢、鳃、体肾、脾脏、肠的表达量相对较低,头肾中最低。
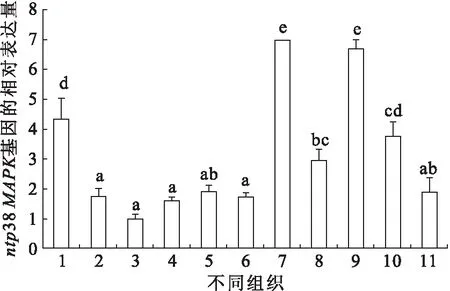
图3 尼罗罗非鱼ntp38MAPK在各组织表达情况Fig.3 Expression of ntp38MAPK in O.niloticus in various tissues1.肝脏,2.脾脏,3.头肾,4.体肾,5.鳃,6.肠,7.肌肉,8.脑,9.心脏,10.血液,11.精巢。小写字母不同表示差异性显著(P<0.05),标有相同小写字母者表示组间无显著性差异(P>0.05)
2.4 无乳链球菌感染后ntp38MAPK表达分析
无乳链球菌感染后ntp38MAPK在肝脏中的表达变化情况如图4。结果显示,感染2 h,表达量逐渐上调,在4 h达到峰值,为对照组的2.1倍。8 h下调为对照组的0.8倍,与对照组差异不显著。感染12 h表达量上调到又一峰值,为对照组的3.9倍。24~72 h表达量趋于对照组水平。
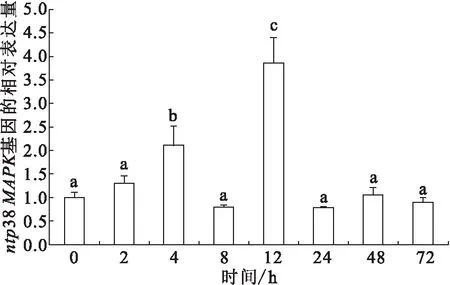
图4 无乳链球菌感染后肝脏中ntp38MAPK的表达情况Fig.4 Expression of ntp38MAPK in liver after streptococcal infection小写字母不同表示差异性显著(P<0.05),标有相同小写字母者表示组间无显著性差异(P>0.05)。下图同。
在无乳链球菌感染的过程中,ntp38MAPK在脾脏中的表达量在感染后2 h明显上调,为对照组的1.6倍。之后表达量开始下调,8 h表达量下降到最低,为对照组的0.06倍。12 h表达量有所回升但显著低于对照组,为对照组的0.5倍。24~72 h,表达量趋于平稳,但显著低于对照组 (图5)。

图5 无乳链球菌感染后脾脏中ntp38MAPK的表达情况Fig.5 Expression of ntp38MAPK in spleen after streptococcal infection
在头肾中的表达量变化趋势与肝脏有些类似,均呈现上调与下调波动性往复变化。无乳链球菌感染后2 h,表达量上调到对照组的1.9倍。之后开始下降,8 h达到最低值,为对照组的0.3倍。12 h表达量又回升,为对照组的1.4倍。24~72 h趋于对照组水平(图6)。

图6 无乳链球菌感染后头肾中ntp38MAPK的表达情况Fig.6 Expression of ntp38MAPK in head kidney after streptococcal infection
3 讨论
p38MAPK在炎症反应、细胞因子产生、转录调节等过程中具有重要作用[5]。为了深入研究尼罗罗非鱼p38MAPK在抗菌免疫方面的作用,本实验克隆了尼罗罗非鱼埃及品系p38MAPK全长序列。该基因编码的氨基酸序列含有p38家族典型保守的Thr-Gly-Tyr(TGY)双磷酸化位点、紧密相连的底物结合位点Ala-Thr-Arg-Trp(ATRW),以及CD结构域。经序列比对以及同源性分析发现,ntp38MAPK与其他物种的相似性都在70%以上,可见p38MAPK在进化中十分保守。通过系统进化分析,尼罗罗非鱼p38MAPK与同为鲈形目的大黄鱼、鲈亲缘关系最近,这与传统的分类系统相一致。综上所述,本实验克隆得到的是尼罗罗非鱼埃及品系的p38MAPK基因。
组织测定结果显示ntp38MAPK在尼罗罗非鱼各组织均广泛表达。类似地,在斑马鱼[9]、文昌鱼[11]等水生生物中,p38MAPK在各组织中均有表达,这与p38MAPK在不同组织参与不同的生理功能有关[15]。但是在不同的物种中,p38MAPK在不同组织中的mRNA丰度并不相同。本实验中,ntp38MAPK在肌肉中的表达量最高,哺乳动物中的研究表明p38MAPK主要在骨骼肌中表达,并参与肌肉的分化调控[16]。然而,拟穴青蟹[17]中p38MAPK表达量最高的组织是血细胞、肠道,而肌肉中的表达量最低;这可能与p38MAPK在不同生物组织中功能的多样性有关[15]。
诸多研究表明,p38MAPK参与调控多种生物的免疫过程,尤其在调节先天免疫反应方面[2]。Jiu等[11]发现在文昌鱼中脂多糖(LPS)刺激能引起p38MAPK表达量变化。Yan等[18]证实p38MAPK在调控中国对虾抵抗病毒细菌感染、清除病原、提高存活率过程中起至关重要的作用。本实验中,无乳链球菌刺激后,尼罗罗非鱼肝脏、脾脏和头肾等免疫组织中ntp38MAPK的表达量显著变化,推测ntp38MAPK参与了罗非鱼抗菌免疫过程。
腹腔注射无乳链球菌后,尼罗罗非鱼脾脏中ntp38MAPK快速响应,在感染后2 h表达量上调到对照组的1.6倍;肝脏中ntp38MAPK在4 h开始响应,12 h达到峰值,为对照组的3.9倍,推测可能与无乳链球菌对组织感染入侵途径有关。利用免疫组织化学定位技术发现,腹腔注射无乳链球菌后病原菌阳性信号首先出现在脾脏,之后是肝脏[19]。脾脏是罗非鱼的主要免疫器官,同时是无乳链球菌的主要攻击对象[20]。而肝脏是鱼类的代谢器官,同时具有一定解毒和防御功能。无乳链球菌侵入鱼体内后,少量病菌入侵肝脏,肝脏内吞噬细胞即可吞噬清除病原菌,但随着病原菌在体内大量繁殖,其毒力因子累积增多,机体需逐渐扩大炎症反应来抵抗病原菌[21],因此肝脏中的反应稍有滞后。同时,不同种类致病菌引起p38MAPK变化时间也有差异。石斑鱼感染海豚链球菌(革兰氏阳性菌)、迟钝爱德华氏菌(革兰氏阴性菌)和巨细胞病毒(病毒)后,p38MAPK的表达量分别在感染3 h、12 h和48 h达到峰值[22];凡纳滨对虾[18]感染革兰氏阴性菌后,血细胞、鳃中p38MAPK的表达量分别在8 h、12 h达到峰值,感染革兰氏阳性菌后,鳃中p38MAPK的表达量在8 h即达到最大值,血细胞中在12 h才达到峰值。另外,头肾是鱼类重要的淋巴组织,在免疫应答过程中起主要作用[21],在感染后的2 h检测到头肾中ntp38MAPK表达量显著上调。感染8 h后肝脏、脾脏和头肾中p38MAPK的表达量均有下调,推测与机体自身免疫调节有关。类似的,海参[23]、对虾[24]感染弧菌后,p38MAPK表达量分别在8 h、4 h明显下调。
尼罗罗非鱼无乳链球菌感染24 h后,肝脏、头肾中ntp38MAPK表达量恢复至对照组水平。同样地,Yu等[17]用金黄色葡萄球菌感染拟穴青蟹,24 h后血细胞中p38MAPK基因表达量恢复到对照组水平。已有研究表明,p38MAPK在免疫过程中具有双重调节作用。当炎症反应到达一定程度后,p38MAPK能通过双特异性磷酸酶(DUSP)介导的MAPK去磷酸化来形成负反馈调节以降低免疫反应对组织带来的损伤[25]。Keranen等[26]发现增加双特异性磷酸酶MKP-1的表达,能够抑制p38MAPK信号通路,从而抑制细胞因子的产生,进而降低炎症反应,使机体达到稳定状态。然而,本实验中脾脏中ntp38MAPK表达量显著低于对照组。推测可能是由于无乳链球菌的长时间感染造成了脾脏组织功能和结构的损坏。病理形态学分析发现,无乳链球菌感染能造成脾脏内红细胞大量破坏,进而导致贫血,免疫功能降低[14]。另外,贺扬[20]运用转录组测序技术分析发现Toll样受体 (TLR5)在无乳链球菌感染4~72 h表达量显著上调,进而介导促炎因子的大量表达,这可能是导致脾脏过度炎症,出现脾脏急性坏死的主要原因。ntp38MAPK在尼罗罗非鱼免疫组织中表达量的变化,说明ntp38MAPK参与了机体抗菌免疫反应,在先天免疫反应中具有重要作用,然而具体的作用机制需要进一步研究。
本实验首次成功克隆获得尼罗罗非鱼的ntp38MAPK全长序列,并利用实时荧光定量PCR技术检测ntp38MAPK的组织表达情况,分析了无乳链球菌感染过程中尼罗罗非鱼肝脏、脾脏、头肾中ntp38MAPK相对表达量的变化,推断p38MAPK基因在尼罗罗非鱼抵抗细菌侵染中起重要作用,为深入研究尼罗罗非鱼p38MAPK信号通路的作用机制以及调控机理奠定了基础。
