温度和外源激素对白蜡吉丁肿腿蜂翅型分化的影响
2020-06-08胡帅王小艺杨忠岐张彦龙
胡帅 王小艺 杨忠岐 张彦龙


摘要 为探讨影响寄生蜂翅型分化的环境因素,促进天敌昆虫的利用,在室内比较了白蜡吉丁肿腿蜂Sclerodermus pupariae在21、24、27、30℃和33℃ 5个温度条件下翅型分化情况,并在24℃下通过对该蜂寄主麻竖毛天牛Thyestilla gebleri幼虫注射早熟素Ⅱ和蜕皮激素,比较不同激素处理对寄生蜂子代雌蜂有翅率的影响。研究结果表明,白蜡吉丁肿腿蜂在27℃和30℃下子代雌蜂的有翅率显著高于其他温度,分别高达62.02%和68.04%。24℃下该蜂雌蜂有翅率最低,仅为0.97%,确定24℃为该蜂的无翅型温度Twl。在Twl下早熟素Ⅱ处理寄主幼虫显著提高了寄生蜂子代雌蜂的有翅率(15.62%)。而用3种浓度的蜕皮激素溶液处理寄主幼虫,其育出的寄生蜂子代翅型均无显著分化,蜕皮激素对白蜡吉丁肿腿蜂的翅型分化无显著影响。在对白蜡吉丁肿腿蜂人工繁育过程中,建议将饲养温度设置为30℃来提高该蜂种群的雌蜂有翅率。结合早熟素Ⅱ具有翅化效应的结果,可以对寄主幼虫进行早熟素Ⅱ处理或者直接在幼期处理子代蜂,从而实现对该天敌资源的高效利用。
关键词 白蜡吉丁肿腿蜂; 温度; 早熟素; 蜕皮激素; 翅型分化
中图分类号: S 476
文献标识碼: A
DOI: 10.16688/j.zwbh.2019089
Effects of temperature and exogenous hormones on wing dimorphism of the
parasitoid Sclerodermus pupariae (Hymenoptera: Bethylidae)
HU Shuai, WANG Xiaoyi, YANG Zhongqi, ZHANG Yanlong
(Key Laboratory of Forest Protection, State Forestry and Grassland Administration, Research Institute of
Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China)
Abstract
In order to determine the effect of environmental factors on wing dimorphism of parasitoid to enhance efforts to use natural enemy insects, the female winged proportions of Sclerodermus pupariae (Hymenoptera: Bethylidae) were compared at five constant temperature regimes (21, 24, 27, 30, and 33℃) in the laboratory. The wing dimorphism of S.pupariae females resulting from the host Thyestilla gebleri treated with prococene Ⅱ and ecdyterone were investigated at 24℃. The results indicated that the winged female proportions of S.pupariae reared under 27℃ and 30℃ reached up to 62.02% and 68.04%, respectively, and were significantly higher than those at other temperatures. The lowest under 24℃ was only 0.97% and it was determined as the wingless temperature (Twl). The hosts treated with P Ⅱ under Twl could increase the proportion of winged females to 15.62%. Ecdyterone of three concentrations had no effects on wing dimorphism of this parasitoid. It is suggested that the rearing temperature of S.pupariae should be set at 30℃ to increase winged female proportion of this parasitoid. P Ⅱ could induce the production of winged females, and thus we propose that the host be appropriately treated with P Ⅱ or directly treat immature parasitoids with P Ⅱ for higher application efficiency of the natural enemy resources.
Key words
Sclerodermus pupariae; temperature; prococene; ecdyterone; wing dimorphism
昆虫是动物界中最早具翅的类群,飞行翅的获得对其进化成功有着重要意义[12]。翅多型现象在很多昆虫中均有发现[34],被认为是昆虫应对环境变化中在种群扩散和繁殖能力之间权衡的一种生态策略,对昆虫种群的适应性进化亦具有重要的生物学意义[5]。大量关于昆虫翅的非遗传多型性的研究结果表明,翅型分化受到诸多环境因子的影响,如温度[69]、光照条件[7,1011]、寄主营养[1213]、拥挤度[1415]等,其中温度对昆虫的翅型分化普遍产生影响。一般认为,温度的升高会促进种群长翅型或有翅型个体的增多,但是不同物种往往对温度的翅型应答模式也显著不同,如低温会刺激桃蚜Myzus persicae (Sulzer)翅的发育,高温反而抑制翅型分化[16];同时也存在昆虫的翅型分化并不受温度影响的情况,如巨膜长蝽Jakowleffia setulosa在4个试验温度下长翅型比例无显著差异[17]。此外,内分泌激素可能在昆虫的翅型分化中发挥了重要的调控作用。Roff[4]提出阈值假说模型,认为昆虫血淋巴中保幼激素滴度超过临界值后,将会发育成短翅型,否则为长翅型。一些通过对昆虫进行外源激素处理的试验结果已证实保幼激素(JH)确有短翅化效应[1820],同时早熟素作为与JH产生生理拮抗的植物类激素显示出良好的长翅化效应[2122],目前这些研究主要集中于飞虱、蟋蟀、蚜虫等农业害虫。
天敌昆虫白蜡吉丁肿腿蜂Sclerodermus pupariae 是近年来发现的肿腿蜂新种[23],因其较强的寄主搜索和主动攻击能力而被广泛应用于我国天牛和吉丁类蛀干害虫的生物防治[2425]。然而,野外无翅型雌蜂只能依靠爬行扩散,使得该蜂的生防效率一定程度上受限,如能通过人为干预培育出较高比例的有翅雌蜂将在生产上具有良好的应用价值。本文以白蜡吉丁肿腿蜂为研究对象,室内比较不同温度下该蜂子代雌蜂翅型分化情况以及用早熟素Ⅱ(precocene Ⅱ, P Ⅱ)和蜕皮激素(20-hydroxyecdysone, 20E)作为外源激素处理寄主麻竖毛天牛Thyestilla gebleri幼虫后寄生蜂有翅比例变化情况,探讨温度对该蜂翅型分化的调控作用,并从生理生化角度揭示寄生蜂翅型分化机理,为提高白蜡吉丁肿腿蜂野外防治效率提供理论基础。
1 材料与方法
1.1 材料
供试昆虫:白蜡吉丁肿腿蜂野外雌蜂种群采自天津市大港区官港森林公园(38°56′N,117°29′E),在室内用麻竖毛天牛幼虫繁殖33代。寄主昆虫麻豎毛天牛幼虫购自天津市花鸟虫鱼市场,选取质量230 mg±10 mg麻竖毛天牛幼虫作为供试寄主。
外源激素类物质:早熟素Ⅱ和蜕皮激素购自Santa Cruz Biotechnology 公司。
微量进样器:5 μL进样管(7633-01)和针头(尖头,87900)均购自Hamilton公司。
1.2 研究方法
1.2.1 不同温度对白蜡吉丁肿腿蜂翅型分化的影响
在扩繁至第33代的白蜡吉丁肿腿蜂种群内,随机挑选75头无翅雌蜂,单头置入装有1头麻竖毛天牛幼虫的指形管(1.0 cm×5.0 cm)内,用脱脂棉塞紧管口,随机分为5组分别放入21、24、27、30℃和33℃的人工气候箱(宁波海曙赛福PRX-450)中,光周期为L∥D=14 h∥10 h,相对湿度为(65±5)%,各处理15个重复。每天观察寄生蜂寄生情况和子代发育情况,待子代蜂羽化后,记录不同温度下各处理子代蜂雌蜂的有翅率(子代有翅雌蜂数量/子代雌蜂总数×100%),并确定白蜡吉丁肿腿蜂的无翅型温度(Twl)。
1.2.2 外源激素对白蜡吉丁肿腿蜂翅型分化的影响
激素溶液配制:取100 mg P Ⅱ溶于少量丙酮,再用ddH2O(双蒸水)稀释为100 μg/μL,同时以丙酮∶ ddH2O=1∶1溶液为对照;取100 mg 20E溶于少量乙醇,再用ddH2O稀释为10 μg/μL,再分别稀释得到1 μg/μL和0.1 μg/μL的20E溶液,再以乙醇∶ddH2O=1∶1溶液作为对照。不注射任何溶液为空白对照。激素处理方法:将寄主天牛幼虫放在冰板上冷冻1 min后,用微量进样器吸取5 μL供试激素溶液快速注射至天牛后胸部柔软处,将处理后的试虫分别单头置入指形管(1.0 cm×5.0 cm)内,管内接入1头寄生蜂F33的无翅雌蜂,脱脂棉塞紧管口。所有供试昆虫蜂均置于Twl,(65±5)% RH,L∥D=14 h∥10 h的人工气候箱内培育。对照组溶液同以上注射方法,各处理组和对照组重复数为15,激素处理参考Khafagi[26]的方法。待子代羽化后统计各处理下子代雌蜂的有翅率。
1.2.3 外源激素对白蜡吉丁肿腿蜂寄生率和子代羽化率的影响
按照1.2.1所述方法,每天记录寄生蜂寄生和子代发育情况,寄生蜂在寄主幼虫上产卵则记为被寄生,寄主幼虫成功繁育出子代蜂则记为成功羽化。各处理寄生率=被产卵处理数/处理总数×100%,子代羽化率=最终成功羽化处理数/被产卵处理数×100%。通过比较外源激素处理寄主幼虫后寄生蜂的寄生率和子代羽化率进一步验证1.2.2中外源激素是否成功注射并作用到寄主幼虫从而干扰寄生蜂发育。
1.3 数据分析
采用卡方检验(χ2)评价不同处理下寄生蜂子代有翅率的差异以及不同处理对寄生蜂寄生率和子代羽化率的影响,组内比较的显著水平经过Bonferroni 校正(P=0.05/两两比较的次数)。所有数据统计分析使用SPSS 20.0 软件完成。
2 结果与分析
2.1 温度对白蜡吉丁肿腿蜂翅型分化的影响
结果显示,温度对白蜡吉丁肿腿蜂子代雌蜂的有翅率有极显著的影响(df=4, χ2=1 786.128, P<0.000 1)(图1)。高温显著提高了子代雌蜂的有翅率,当温度升高至27℃时,子代雌蜂有翅率锐增至60%以上。但当温度超过30℃时,有翅率再次降低。在5个试验温度下,白蜡吉丁肿腿蜂子代雌蜂有翅率在24℃下最低,为0.97%,确定该蜂的Twl为24℃。
2.2 早熟素Ⅱ处理对白蜡吉丁肿腿蜂翅型分化的影响
在无翅型环境(24℃)下,早熟素Ⅱ溶液和其他两组对照溶液处理寄主幼虫时,白蜡吉丁肿腿蜂子代雌蜂有翅率差异极显著(图2)。用100 μg/μL的早熟素Ⅱ处理的寄主幼虫上的子代雌蜂有翅率显著高于丙酮对照组和空白对照组(df=2, χ2=95.118, P<0.000 1),说明P Ⅱ具有明显的具翅化效应。
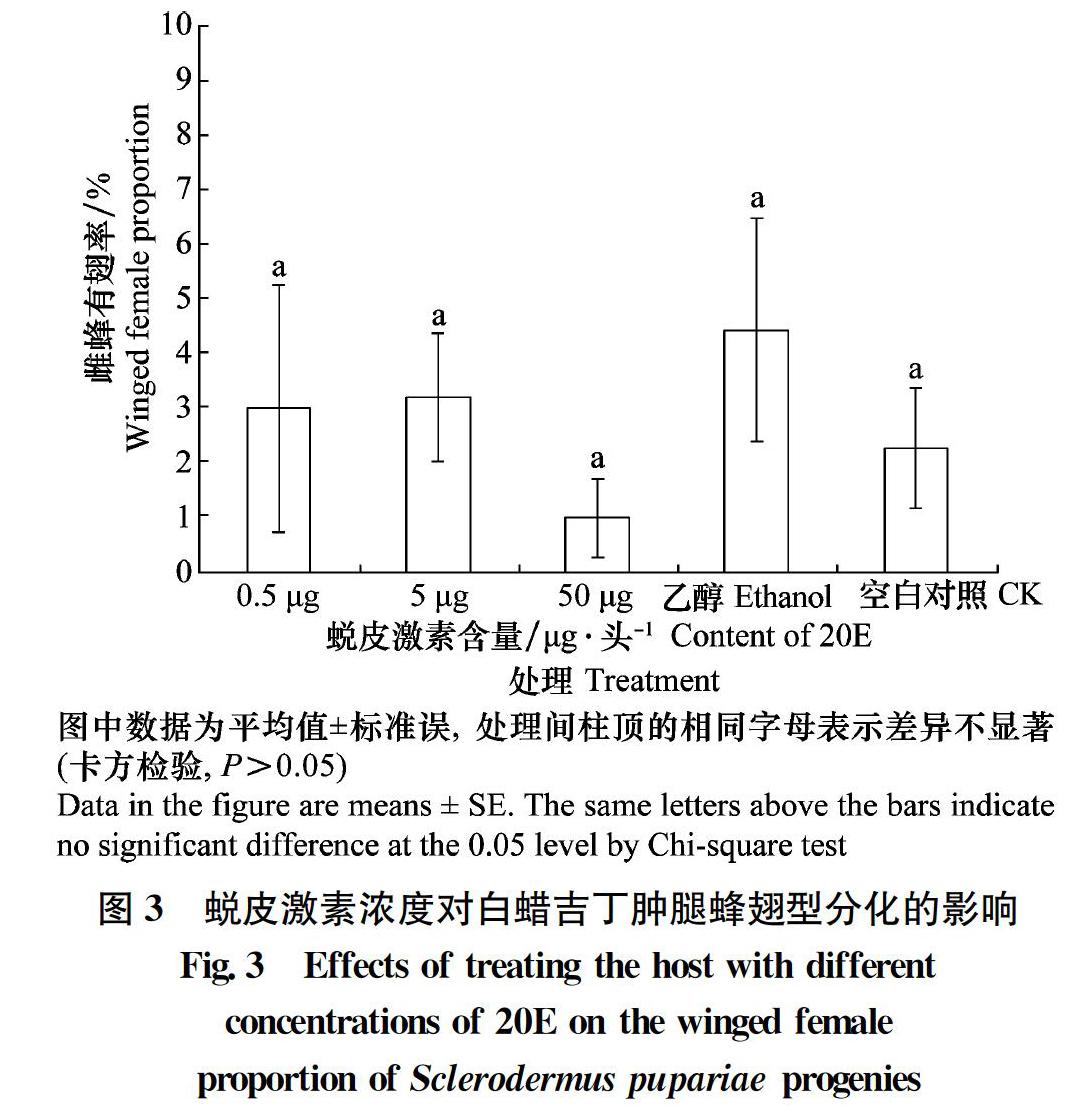
2.3 蜕皮激素浓度对白蜡吉丁肿腿蜂翅型分化的影响
在无翅型温度(24℃)下,不同浓度蜕皮激素(10、1 μg/μL和0.1 μg/μL)分别处理寄主幼虫时,寄生蜂子代雌蜂有翅率均低于5%,和对照组无显著差异(df=4, χ2=8.185, P=0.085)(图3)。
2.4 早熟素Ⅱ和蜕皮激素处理对寄生蜂寄生率和羽化率的影响
研究结果表明,外源激素和对照溶液处理寄主幼虫后,白蜡吉丁肿腿蜂可以寄生试验组的所有寄主,但是寄生蜂对不同溶液处理的寄主幼虫的寄生率和子代羽化率显著不同(表1)。寄生蜂对用10 μg/μL蜕皮激素处理的寄主幼虫的寄生率以及子代羽化率均显著低于其他试验组(卡方检验: χ2=24.798,df=5,P< 0.001; χ2=15.508,df=5,P=0.008),说明通过向寄主幼虫注射外源激素成功使得激素影响到寄生蜂子代。
3 讨论
温度作为重要的季节信号对昆虫的翅型分化有一定的调控作用[2729]。昆虫的翅型分化在不同的温度区段情况各异,如葡萄根瘤蚜Daktulosphaira vitifoliae在16~28℃范围内有翅蚜率随着温度升高而升高,但当温度继续升高至32℃时有翅蚜率明显下降[8];正常范围内的温度变化(25℃、30℃)对曲脉姬蟋Modicogryllus confirmatus Walker的翅型分化无显著影响,而在35℃的极高温环境下显著降低了种群长翅率。 温度对白蜡吉丁肿腿蜂翅型分化的调控作用明显,随着温度的升高,寄生蜂有翅个体比例逐渐升高,在30℃时子代雌蜂有翅率接近70%,说明高温是促进该蜂种群有翅型个体出现的重要诱因。但是当温度继续升高至33℃时,有翅率显著下降,这可能是高温胁迫作用导致[30]。此外,白蜡吉丁肿腿蜂在24℃下的有翅率为0.97%,极显著低于其他4个处理组。冬末初春气温回暖,越冬代高比例有翅型个体扩散后寻找到新栖息地[3132],此时高繁殖力的无翅个体的大量出现显然更有利于寄生蜂种群的繁衍。因此,我们推测24℃即为室内饲养该蜂的无翅型温度Twl。
前人的研究认为环境因子对翅型分化的作用实际上是因为昆虫内分泌能力受到环境影响,从而抑制或者促进体内激素的分泌最后导致了不同翅型分化的结果[3334]。在昆虫翅多型现象中,保幼激素及其类似物促进昆虫向无翅化或者短翅化发育在昆虫翅多型现象的研究已得到证明[18,3536]。因此本文用蜕皮激素和早熟素Ⅱ来探究外源激素处理是否能调控白蜡吉丁肿腿蜂产生更多具翅型个体。已有研究证明通过给寄主昆虫人为注射外源性激素,从而间接对寄生蜂进行激素处理的方式可以影响幼蜂的生长发育,如用微量进样器对寄主灰翅夜蛾Spodoptera littoralis (Boisd.) 进行保幼激素和早熟素处理后,侧沟茧蜂Microplitis rufiventris Kok.的子代数量减少并且高比例的子代蜂无活动能力[26];也有研究发现,从保幼激素类似物处理过的寄主麻蝇Sarcophaga bullata上羽化的麗蝇蛹集金小蜂Nasonia vitripennis数量和对照组无显著差别,推测可能是因为寄主的新陈代谢将外源激素类似物进行了去活性转化[37]。本文沿用寄主注射法,研究了激素处理麻竖毛天牛幼虫后,其育出的白蜡吉丁肿腿蜂翅型分化的情况。白蜡吉丁肿腿蜂是一种卵育型抑性外寄生蜂,对其寄主幼虫进行不同激素和不同浓度蜕皮激素注射后, 母代蜂对寄主幼虫的寄生率、羽化率以及子代蜂翅型分化情况差异显著,说明注射到寄主体内的外源激素成功处理到了子代蜂。
结果表明,在Twl下早熟素Ⅱ仍能诱导寄生蜂产出有翅率更高的子代,具有明显的具翅化效应;3种不同浓度的蜕皮激素对该蜂的翅型分化均无明显影响。此结果和对白背飞虱Sogatella furcifera[22]及褐飞虱Nilaparvata lugens[38]的研究结果相吻合,也有研究证明蜕皮激素参与调控麦长管蚜Sitobion avenae 的翅型分化[39]。本文白蜡吉丁肿腿蜂在试验浓度的早熟素Ⅱ处理后子代翅型分化明显,然而提高后的子代有翅率仅为15.62%,这可能是因为低温弱化了激素对翅型的调控作用,但是二者如何关联作用于该蜂的翅型分化仍不清楚,也有可能是因为只对寄主幼虫进行一次性注射,导致部分活性物质被寄主分解所致。也有学者认为决定外源激素对昆虫翅型分化影响大小的关键因素是处理时的虫龄,和浓度关系并不密切[40]。总之,昆虫体内激素调控下的生理生化过程在变态期间是十分复杂的,本文研究结果为白蜡吉丁肿腿蜂对外源激素处理下翅型应答机制提供了初步参考依据,进一步研究将关注不同浓度早熟素处理对翅型分化的作用机制、不同龄期处理对翅型结果的影响以及激素处理后置于不同温度下寄生蜂的翅型分化情况。
参考文献
[1] MISOF B, LIU S, MEUSEMANN K, et al. Phylogenomics resolves the timing and pattern of insect evolution [J]. Science, 2014, 346(6210): 763767.
[2] 朱道弘. 昆虫翅型分化的调控及翅多型性的进化[J]. 生态学报, 2009, 46(1): 1116.
[3] 王小艺, 杨忠岐, 魏可, 等. 昆虫翅型分化的表型可塑性机制[J]. 生态学报, 2015, 35(12): 39883999.
[4] ROFF D A. The evolution of wing dimorphism in insects [J]. Evolution, 1986, 40(5): 10091020.
[5] ZERA A J, DENNO R F. Physiology and ecology of dispersal polymorphism in insects [J]. Annual Review of Entomology, 1997, 42(1): 207230.
[6] SASAKI R, NAKASUJI F, FUJISAKI K. Environmental factors determining wing form in the lygaeid bug, Dimorphopterus japonicas (Heteroptera: Lygaeidae) [J]. Applied Entomology and Zoology, 2002, 37(2): 329333.
[7] 曾杨, 朱道弘, 赵吕权. 环境因素对长颚斗蟋翅型分化的影响[J]. 生态学报, 2009, 30(21): 60016008.
[8] 朱化平, 杨晓岩, 季兴龙, 等. 环境因子对葡萄根瘤蚜翅型分化的影响[J]. 山东农业科学, 2015, 47(2): 100103.
[9] WEI Y J. Wing polymorphism in Nysius huttoni White (Hemiptera: Orsillidae) [J]. New Zealand Journal of Zoology, 2011, 38(1): 114.
[10]WALKER T J. Wing dimorphism in Gryllus rubens (Orthoptera: Gryllidae) [J]. Annals of the Entomological Society of America, 1987, 80(5): 547560.
[11]刘佳妮, 桂富荣, 李正跃. 影响白背飞虱翅型分化的相关因子[J]. 植物保护学报, 2010, 37(6): 511516.
[12]邹运鼎, 杨义和, 胡雨娟, 等. 影响两种麦蚜成蚜有翅率的因子分析[J]. 应用生态学报, 1997, 8(2): 189193.
[13]LIN Xinda, XU Yili, JIANG Jianru, et al. Host quality induces phenotypic plasticity in a wing polyphenic insect [J]. Proceedings of the National Academy of Sciences, 2018, 115(29): 75637568.
[14]KISIMOTO R. Effect of crowding during the larval period on the determination of the wing-form of an adult plant-hopper [J]. Nature, 1956, 178: 641642.
[15]LAMB K P, WHITE D. Effect of temperature, starvation and crowding on production of alate young by the cabbage aphid (Brevicoryne brassicae) [J]. Entomologia Experimentalis et Applicata, 2011, 9(2): 179184.
[16]刘树生, 吴晓晶. 温度对桃蚜和萝卜蚜翅型分化的影响[J]. 昆虫学报, 1994, 37(3): 292297.
[17]王旭娜, 黄新动, 白晓拴. 温度对巨膜长蝽翅型分化的影响初探[J]. 内蒙古民族大学学报(自然科学版), 2018, 33(2): 152154.
[18]HARDIE J. Juvenile hormone mimics the photoperiodic apterization of the alate gynopara of aphid, Aphis fabae [J]. Nature, 1980, 286(5773): 602604.
[19]IWANAGA K, TOJO S. Hormonal control of wing polymorphism in the brown planthopper, Nilaparvata lugens [J]. Kyushu Plant Protection Research, 1985, 31: 8488.
[20]ZERA A J, TIEBEL K C. Brachypterizing effect of group rearing, juvenile hormone III and methoprene in the wing-dimorphic cricket, Gryllus rubens [J]. Journal of Insect Physiology, 1988, 34(6): 497498.
[21]BERTUSO A G, MOROOKA S, TOJO S. Sensitive periods for wing development and precocious metamorphosis after precocene treatment of the brown planthopper, Nilaparvata lugens [J]. Journal of Insect Physiology, 2002, 48(2): 221229.
[22]张琼秀, 孙梓暄, 李广宏, 等. 三种外源激素类物质对白背飞虱(Sogatella furcifera)翅型分化的影响[J]. 生态学报, 2008, 28(12): 59945998
[23]YANG Zhongqi, WANG Xiaoyi, YAO Yanxia, et al. A new species of Sclerodermus (Hymenoptera: Bethylidae) parasitizing Agrilus planipennis (Coleoptera: Buprestidae) from China, with a key to Chinese species in the genus [J]. Annals of the Entomological Society of America, 2012, 105(5): 619627.
[24]王小艺, 杨忠岐, 唐艳龙, 等. 白蜡吉丁肿腿蜂对栗山天牛低龄幼虫的寄生作用[J]. 昆虫学报, 2010, 53(6): 675682.
[25]杨忠岐, 王小艺, 张翌楠, 等. 以生物防治为主的综合控制我国重大林木病虫害研究进展[J]. 中国生物防治學报, 2018, 34(2): 163183.
[26]KHAFAGI W E. Effects of juvenile hormone I, precocene I and precocene Ⅱ on the progeny of Microplitis rufiventris Kok. female when administered via its host, Spodoptera littoralis (Boisd.) [J]. Journal of Applied Entomology, 2004, 128(1): 610.
[27]SASAKI R, NAKASUJI F, FUJISAKI K. Seasonal changes in wing dimorphism of the lygaeid bug Dimorphopterus japonicus (Heteroptera: Lygaeidae) in relation to environmental factors [J]. Entomological Science, 2003, 6(2): 6370.
[28]SHEARER P W, WEST J D, WALTON V M, et al. Seasonal cues induce phenotypic plasticity of Drosophila suzukiito enhance winter survival [J]. BMC Ecology, 2016, 16(1): 1118.
[29]SAKASHITA T F, FUJISAKI K. Seasonal variation in wing polymorphism of the pyrrhocorid bug, Pyrrhocoris sibiricus (Heteroptera: Pyrrhocoridae) [J]. Applied Entomology & Zoology, 1998, 33(2): 243246.
[30]SHIMIZU T, MASAKI S. Injury causes microptery in the ground cricket, Dianemobius fascipes [J]. Journal of Insect Physiology, 1993, 39(12): 10211027.
[31]武輝, 王小艺, 李孟楼, 等. 白蜡吉丁肿腿蜂的生物学和生态学特性及繁殖技术研究[J]. 昆虫学报, 2008, 51(1): 4654.
[32]WANG Xiaoyi, WEI Ke, YANG Zhongqi, et al. Effects of biotic and abiotic factors on phenotypic partitioning of wing morphology and development in Sclerodermus pupariae (Hymenoptera: Bethylidae) [J]. Scientific Reports, 2016, 6: 26408.
[33]马巨法, 唐健, 胡国文, 等. 稻褐飞虱成虫翅二型现象[J].昆虫知识, 1995, 32(3): 174178.
[34]戴华国, 吴晓毅, 武淑文. 褐飞虱体内保幼激素滴度变化及其与翅型分化的关系[J]. 昆虫学报, 2001, 44(1): 2732.
[35]NIJHOUT H F. Insect hormones[M]. Princeton: Princeton University Press, 1994.
[36]HARDIE J, HONDA K I, TIBOR Timár, et al. Effects of 2, 2-dimethylchromene derivatives on wing determination and metamorphosis in the pea aphid, Acyrthosiphon pisum [J]. Archives of Insect Biochemistry & Physiology, 1995, 30(1): 2540.
[37]FASHING N J, SAGAN H. Effect of the juvenile hormone analog methoprene on Nasonia vitripennis when administered via a host, Sarcophaga bullata [J]. Environmental Entomology, 1979, 8(5):816818.
[38]张琼秀, 孙梓暄, 李广宏, 等. 外源激素对褐飞虱翅型分化的影响[J]. 中山大学学报(自然科学版), 2007, 46(3): 7678.
[39]魏长平, 杨超霞, 李祥瑞, 等. 蜕皮激素参与蚜虫跨代翅型分化调控研究[C]∥中国植物保护学会学术年会论文集, 北京: 中国农业科学技术出版社, 2018: 105.
[40]王健, 吴振廷, 张一九, 等. 外源激素对褐飞虱翅型分化的影响[J]. 昆虫学报, 1998, 41(4): 371375.
(责任编辑:田 喆)
收稿日期: 20190227 修订日期: 20190326
基金项目:
林业公益性行业科研专项(201404403);国家自然科学基金(31370654)
致 谢: 参加本试验部分工作的还有江代礼、谭翰杰、张能和纪烨斌等同学,特此一并致谢。
通信作者 E-mail:xywang@caf.ac.cn
#为并列第一作者
