若尔盖退化高寒湿地土壤理化性质、酶活性及微生物群落的季节动态
2020-06-03孙飞达陈文业苟文龙周冀琼
孙飞达,李 飞,陈文业,朱 灿,周 俗,苟文龙,刘 琳,周冀琼
1 四川农业大学动物科技学院, 成都 611130 2 甘肃省林业科学研究院, 兰州 730020 3 四川省草原科学研究院, 成都 611731
若尔盖地处青藏高原东缘,是长江、黄河的重要水源地区,也是典型的生态脆弱区和气候变化警示区,天然草地面积为8.08×105hm2,其中可利用草地面积占80%以上,畜牧业产值约占全县农业总产值的95%[1- 2]。由于自然和人为双重因素使若尔盖地区草地生产力急速下降、鼠害频发,并在局部地区形成了严重退化的鼠荒地。草原鼠荒地(Rodent Damaged Rangeland)是草地退化的一种表现形式,通常是在草地逆向演替进程中,产生了适宜于啮齿类动物栖息生活的小环境,鼠类的入侵、啃食、掘洞、刈割、贮草行为进而引起草原退化加剧的一类退化草地,主要有沙化型鼠荒地、沼泽退化型鼠荒地等类型[3]。鼠荒地上优质牧草比例大幅降低,草地生产、生活功能受到严重影响,生态服务价值和服务水平也明显降低,土壤裸露、植被稀少、鼠类入侵使得草地土壤的理化性质和生物学特征发生改变[4- 5]。草地“三生功能”衰退不仅影响当地的社会经济发展,甚至对下游川西平原、川中丘陵地区产生了深远的影响[6]。
高原鼢鼠(Myospalaxbaileyi)是若尔盖地区主要害鼠之一,主要分布在高寒草甸生态系统,常常栖息于土质疏松、植被退化、杂类草丰富的斑块地,主要采食粗蛋白和水溶性糖含量高、粗纤维含量低的杂类草[7- 9]。高原鼢鼠掘土造丘过程中,大量下层土壤被翻至地面形成土丘,土丘数量往往能指示该区域高原鼢鼠种群大小,植被返青期5月和枯黄期10月新土丘与高原鼢鼠种群数量呈显著正相关,利用新土丘密度来预测高原鼢鼠的种群密度最为准确[10- 11]。高原鼢鼠被誉为“生态系统的工程师”,种群密度维持在较低密度时,作为生态系统的重要组成部分,能够维持草原生态系统的平衡和稳定[12- 14]。高原鼢鼠挖掘形成的洞穴系统和采食行为通过影响土壤理化性质、酶活性以及微生物群落,进而对草地植被群落的组成和产草量造成影响,一定程度上有利于退化草地的植被恢复重建[15- 17]。但高原鼢鼠密度一旦超过阈值会引起草地急速退化,随着土丘密度增加,草地植物群落优势种迅速由莎草科和禾本科转变为杂类草,地上、地下生物量显著降低,土壤有机质、全氮、速效氮和速效磷等养分含量显著降低,土壤酶活性和微生物群落随土壤养分变化而变化[18- 20]。
目前研究集中在不同退化演替阶段高原鼢鼠活动对草地植被群落、土丘土壤理化性质的影响,多采用空间代替时间序列的方法,对于高原鼢鼠鼠荒地非土丘土壤理化性质、酶活性及微生物群落的季节动态研究尚鲜见报道[21]。本研究于2015—2017三年间连续监测严重退化高原鼢鼠鼠荒地上鼠丘数量、不同土层深度土壤理化性质及土壤生物学特征的季节动态并分析其相关性,以期能揭示不同植被物候期,高原鼢鼠活动对鼠荒地草地植被-土壤-微生物生态系统的影响。
1 试验地概况
试验地位于若尔盖县麦溪乡,地理位置33°56′—33°58′N,102°11′—102°18′E之间,海拔3439 m,距若尔盖县城约97 km,临近甘肃省玛曲县,2015年至2017年各月均温及月降水量如图1(布瑞克农业气象数据库),相对湿度约为70%,年均日照时数约为2000 h,全乡天然草原面积7.25×104hm2,是一个以畜牧业为主要支柱产业的纯牧业乡。

图1 若尔盖地区三年间(2015—2017)月均温和月降水量的平均值Fig.1 Average of monthly mean temperature and monthly precipitation in Zoige from 2015—2017
试验地面积约100 hm2,20世纪50年代曾为高寒沼泽湿地区,由于历史上排水垦田的政策失配,此区域逐渐干涸,再加上长期的超载过牧,草场退化严重,鼠害加剧,从2010年以来,该区域设为禁牧区并长年围封。试验地内高原鼢鼠为唯一造丘鼠类且种群密度较高,未见高原鼠兔(Ochotonacurzoniae)及其他鼠类活动踪迹,根据刘荣堂等[22]对高原鼢鼠危害等级的划分属于高原鼢鼠重度危害地区,鼠丘密度变化如表1。植被以西伯利亚蓼(Polygonumsibiricum)、鹅绒委陵菜(Potentillaanserina)、四川嵩草(Kobresiasetchwanensis)为主,植被盖度和草层高度均较低。

表1 高寒鼠荒地不同植被物候期高原鼢鼠土丘密度变化情况
不同小写字母表示植被物候期间差异显著(P<0.05)
2 研究方法
2.1 试验设计
自2015年起连续三年于每年草地返青期(5—6月)、草盛期 (7—8月)、枯黄期(10—11月)在试验地采用鼠丘计数法选择 9个半径28.2 m的样地(样地间距离50 m以上)进行鼠丘数量统计(新旧土丘分别计数)及土壤取样[23- 24]。土壤采样使用土钻沿一条直线在临近土丘(非土丘区域)分别钻取0—10、10—20、20—30 cm土层土壤,5次重复,各土层土壤混合后,一部分带回实验室自然风干后研磨过筛用于测定土壤化学性质及酶活性,一部分用冰盒保存带回实验室迅速进行微生物指标测定。
2.2 指标测定
土壤样品测定方法参照《土壤农业化学分析方法》和《土壤微生物研究原理与方法》[25],土壤pH值采用酸度计法,容重采用环刀法,有机质采用高温外热重铬酸钾氧化法,全氮采用凯式定氮法,碱解氮采用碱解扩散法,有效磷采用钼锑抗比色法,速效钾采用火焰光度法;脲酶采用苯酚钠-次氯酸钠比色法,碱性磷酸酶采用磷酸苯二钠比色法,蔗糖酶采用3,5-二硝基水杨酸比色法;微生物数量采用稀释平板计数法。
2.3 数据分析
应用Excel 2010、SPSS 19.0和 Canoco 4.5进行数据统计整理及分析作图。高原鼢鼠鼠丘密度、土壤理化性质及生物学特征指标均采用单因素Anova分析,数据表示为平均值±标准差,多重比较采用Duncan法;使用多因素方差分析进行物候期和土层深度对土壤酶活性和微生物数量影响的统计分析;土壤生物学特征(土壤脲酶、碱性磷酸酶、蔗糖酶、细菌、真菌、放线菌、微生物总量)共7个指标指标作为Canoco软件中的species部分,高原鼢鼠土丘密度(新土丘、旧土丘和总土丘密度)、土壤理化性质(pH值、含水量、容重、有机质、全氮、碱解氮、有效磷、速效钾、碳氮比)共12个指标作为Canoco软件中的environment部分进行冗余分析(Redundancy analysis,RDA)和主成分(Principal component analysis,PCA)分析并采用Cano Drw 4作图。
3 结果与分析
3.1 土壤理化性质
不同土层深度土壤理化性质差异明显(表2)。随着土层深度增加,高寒鼠荒地各植被物候期土壤pH值、容重均显著升高(P<0.05),土壤含水量、有机质、全氮、碱解氮、有效磷和速效钾均显著降低(P<0.05);各植被物候期土壤碳氮比20—30 cm土层均显著大于10—20 cm土层(P<0.05),0—10 cm土层在返青期和草盛期显著小于20—30 cm土层(P<0.05),枯黄期则显著大于20—30 cm土层(P<0.05)。高寒鼠荒地不同植被物候期土壤理化性质呈现一定规律(表2)。从植被返青期至枯黄期,各土层土壤pH值均呈先降低后升高趋势,枯黄期各土层土壤pH值均显著高于草盛期和返青期(P<0.05);枯黄期各土层土壤容重均显著大于草盛期和返青期(P<0.05);0—10 cm土层土壤土壤有机质枯黄期显著大于草盛期和返青期(P<0.05),而10—20 cm和20—30 cm土层却显著小于草盛期和返青期(P<0.05);返青期鼠荒地0—10、10—20、20—30 cm土层分别占总有机质的46%、28%和26%,草盛期分别占到39%、34%和27%,枯黄期分别占到51%、29%、20%(图2);全氮、碱解氮、碳氮比、有效磷、速效钾也呈现类似的规律。
3.2 土壤酶活性及微生物数量
不同土层深度对高寒鼠荒地土壤酶活性和微生物数量指标均有显著影响(P<0.05),不同植被物候期对土壤酶活性和土壤微生物数量指标(除真菌外)也有显著影响(P<0.05),土层深度和植被物候期交互作用对脲酶、碱性磷酸酶、放线菌和微生物总数指标有显著影响(P<0.05)(表3)。表明土层深度和植被物候期变化均对土壤酶活性和微生物数量指标产生影响,且有明显的交互效应。
随着土层深度增加,高寒鼠荒地各植被物候期土壤脲酶、碱性磷酸酶和蔗糖酶指标含量均显著降低(P<0.05)。从返青期至枯黄期,各土层脲酶、蔗糖酶指标含量逐渐升高,草盛期和枯黄期显著高于返青期(P<0.05);0—10 cm和10—20 cm土层碱性磷酸酶指标含量逐渐升高,枯黄期显著高于草盛期和返青期,20—30 cm各物候期间差异不显著(P<0.05)(图3)。
表2 高寒鼠荒地不同植被物候期和不同土层深度土壤的理化性质
Table 2 Soil physicochemical properties within three soil depths at different vegetation phenological phase in rodent-damaged alpine rangeland
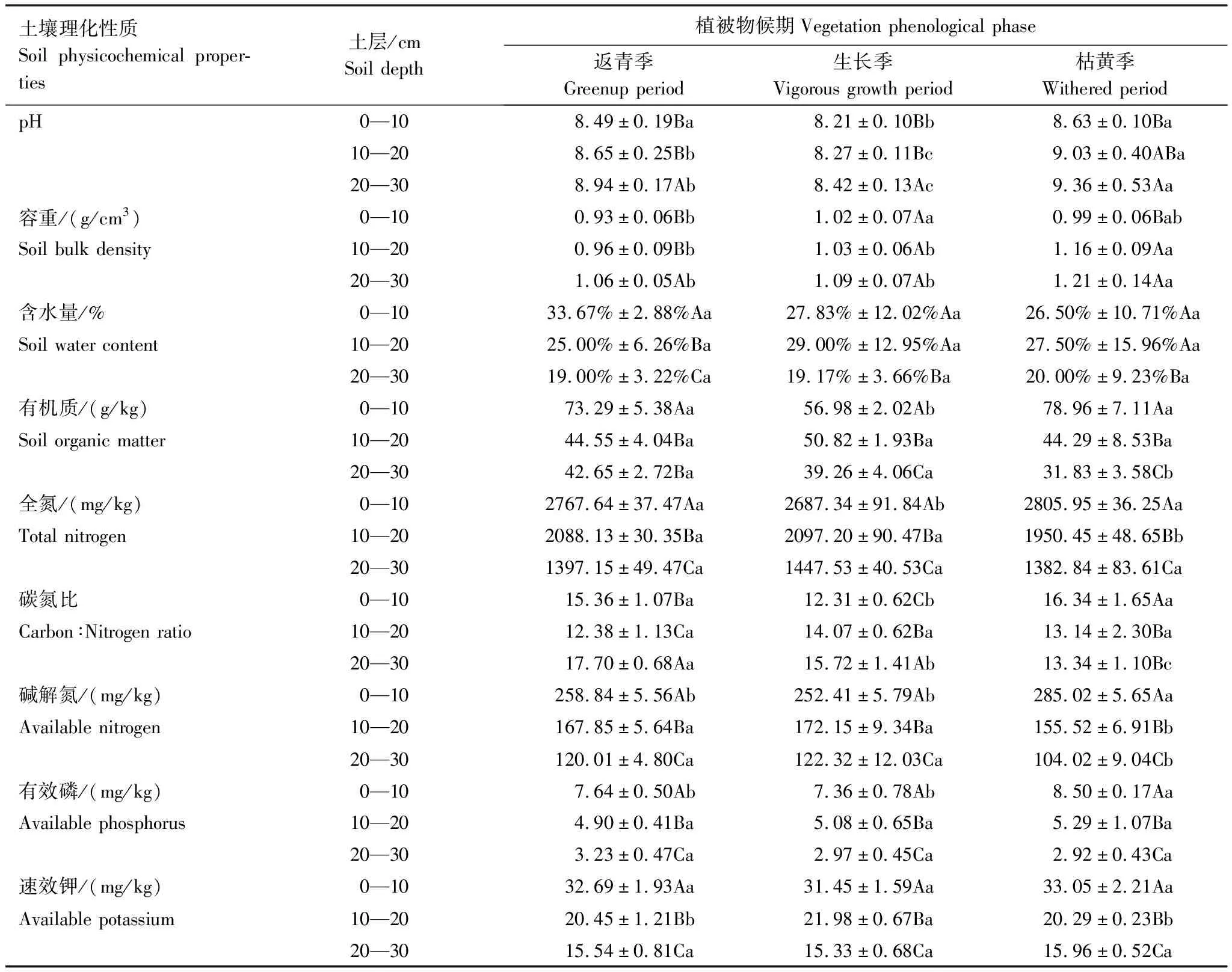
土壤理化性质Soil physicochemical proper-ties土层/cmSoil depth植被物候期Vegetation phenological phase返青季Greenup period生长季Vigorous growth period枯黄季Withered periodpH0—108.49±0.19Ba8.21±0.10Bb8.63±0.10Ba10—208.65±0.25Bb8.27±0.11Bc9.03±0.40ABa20—308.94±0.17Ab8.42±0.13Ac9.36±0.53Aa容重/(g/cm3)0—100.93±0.06Bb1.02±0.07Aa0.99±0.06BabSoil bulk density10—200.96±0.09Bb1.03±0.06Ab1.16±0.09Aa20—301.06±0.05Ab1.09±0.07Ab1.21±0.14Aa含水量/%0—1033.67%±2.88%Aa27.83%±12.02%Aa26.50%±10.71%AaSoil water content10—2025.00%±6.26%Ba29.00%±12.95%Aa27.50%±15.96%Aa20—3019.00%±3.22%Ca19.17%±3.66%Ba20.00%±9.23%Ba有机质/(g/kg)0—1073.29±5.38Aa56.98±2.02Ab78.96±7.11AaSoil organic matter10—2044.55±4.04Ba50.82±1.93Ba44.29±8.53Ba20—3042.65±2.72Ba39.26±4.06Ca31.83±3.58Cb全氮/(mg/kg)0—102767.64±37.47Aa2687.34±91.84Ab2805.95±36.25AaTotal nitrogen10—202088.13±30.35Ba2097.20±90.47Ba1950.45±48.65Bb20—301397.15±49.47Ca1447.53±40.53Ca1382.84±83.61Ca碳氮比0—1015.36±1.07Ba12.31±0.62Cb16.34±1.65AaCarbon∶Nitrogen ratio10—2012.38±1.13Ca14.07±0.62Ba13.14±2.30Ba20—3017.70±0.68Aa15.72±1.41Ab13.34±1.10Bc碱解氮/(mg/kg) 0—10258.84±5.56Ab252.41±5.79Ab285.02±5.65AaAvailable nitrogen10—20167.85±5.64Ba172.15±9.34Ba155.52±6.91Bb20—30120.01±4.80Ca122.32±12.03Ca104.02±9.04Cb有效磷/(mg/kg) 0—107.64±0.50Ab7.36±0.78Ab8.50±0.17AaAvailable phosphorus10—204.90±0.41Ba5.08±0.65Ba5.29±1.07Ba20—303.23±0.47Ca2.97±0.45Ca2.92±0.43Ca速效钾/(mg/kg)0—1032.69±1.93Aa31.45±1.59Aa33.05±2.21Aa Available potassium10—2020.45±1.21Bb21.98±0.67Ba20.29±0.23Bb20—3015.54±0.81Ca15.33±0.68Ca15.96±0.52Ca
不同大写字母表示不同土层间差异显著,不同小写字母表示月份间差异显著(P<0.05)
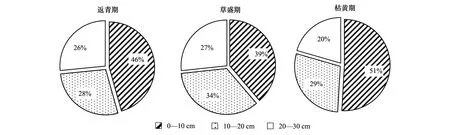
图2 高寒鼠荒地不同物候期土壤有机质在不同土层的分配比例Fig.2 Distribution ratio of soil organic maters within three soil depths at different vegetation phenological phase in rodent-damaged alpine rangeland
表3 高寒鼠荒地不同草地物候期和不同土层土壤生物学特征的方差分析
Table 3 Variance analysis of soil biological characteristics factors within three soil depths at different vegetation phenological phase in rodent-damaged alpine rangeland
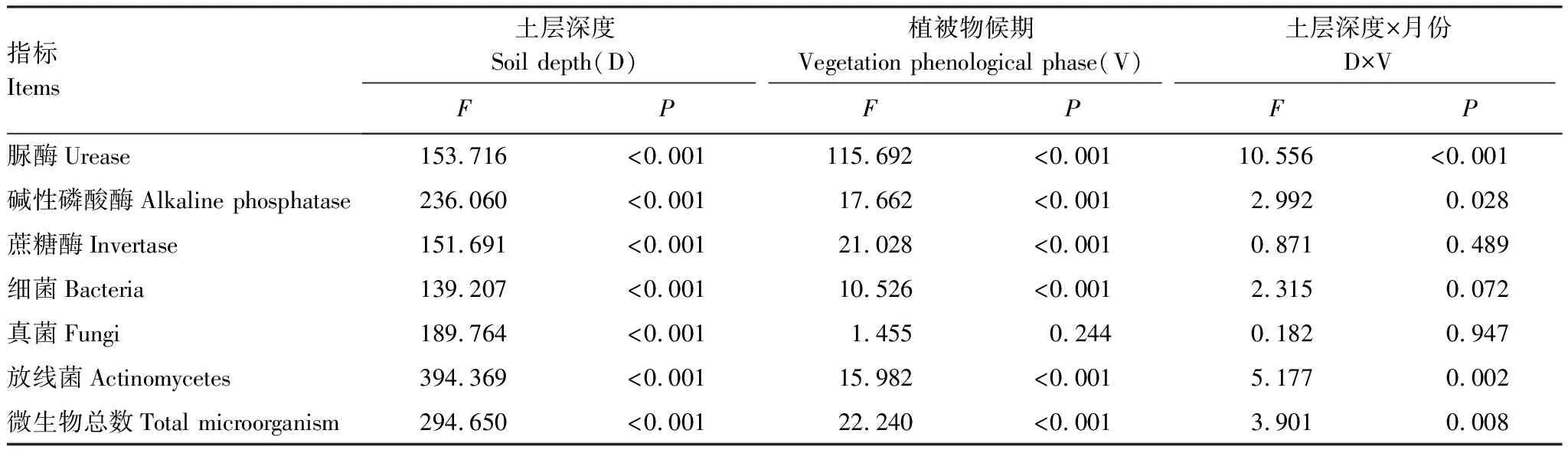
指标Items土层深度Soil depth(D)植被物候期Vegetation phenological phase(V)土层深度×月份D×VFPFPFP脲酶Urease153.716<0.001115.692<0.00110.556<0.001碱性磷酸酶Alkaline phosphatase236.060<0.00117.662<0.0012.9920.028蔗糖酶Invertase151.691<0.00121.028<0.0010.8710.489细菌Bacteria139.207<0.00110.526<0.0012.3150.072真菌Fungi189.764<0.0011.4550.2440.1820.947放线菌Actinomycetes394.369<0.00115.982<0.0015.1770.002微生物总数Total microorganism294.650<0.00122.240<0.0013.9010.008
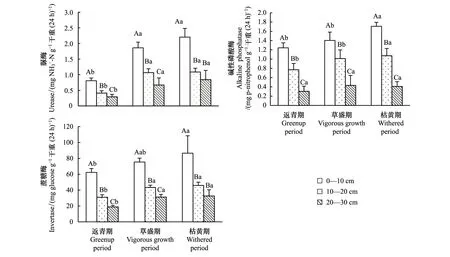
图3 高寒鼠荒地不同植被物候期和不同土壤深度下土壤酶活性含量的季节变化Fig.3 Seasonal dynamics of soil enzyme activities within three soil depths at different vegetation phenological phase in rodent-damaged alpine rangeland
高寒鼠荒地土壤细菌、真菌、放线菌及微生物总数含量均随着土层深度增加显著降低(P<0.05)。从植被返青期至枯黄期,0—10 cm和10—20 cm土层细菌含量逐渐升高,枯黄期显著高于草盛期和返青期(P<0.05),20—30 cm土层植被草盛期显著低于返青期和枯黄期(P<0.05);0—10 cm和10—20 cm土层真菌含量差异不显著(P<0.05),20—30 cm土层草盛期显著高于于返青期和枯黄期;放线菌含量除0—10 cm土层植被返青期显著低于草盛期和枯黄期(P<0.05),其他土层各植被物候期间差异不显著(P<0.05);微生物总数含量逐渐升高,植被返青期显著低于草盛期和枯黄期(P<0.05)(图4)。

图4 高寒鼠荒地不同植被物候期和不同土壤深度下土壤微生物群落含量的季节变化Fig.4 Seasonal dynamics of the content of soil microbial communities within three soil depths at different vegetation phenological phase in rodent-damaged alpine rangeland
3.3 鼠丘密度、土壤理化性质与土壤生物学特征的相关性分析
如图5,将各物候期土壤脲酶、碱性磷酸酶、蔗糖酶、细菌、真菌、放线菌、微生物总数共7个指标指标作为Canoco软件中的species部分,高原鼢鼠新土丘数、旧土丘数、总土丘数、土壤pH值、含水量、容重、有机质、全氮、碱解氮、有效磷、速效钾、碳氮比共12个指标作为Canoco软件中的environment部分,首先用species数据进行去趋势对应分析(Detrended correspondence analysis,DCA)分析,发现返青期、草盛期、枯黄期各排序轴中梯度长度的最大值分别为0.302,0.266和0.270,均小于3,故采用线性模型RDA分析鼠丘密度、土壤理化性质及土壤生物学特征指标的相关性较为合适。结果发现,不同植被物候期土壤生物学特征指标与土壤理化性质间均有显著的相关性(P<0.05),与鼠丘密度无显著相关性(P<0.05)。返青期,土壤微生物特征指标受控于土壤有机质、全氮、碱解氮、有效磷、速效钾土壤性状,二者呈正相关关系,其排序轴特征值为92.2%;土壤容重、pH值、碳氮比对土壤生物学特征具有微弱的拟制作用,其排序轴特征值为3.3%;草盛期与返青期相关性关系类似,第1排序轴和第2排序轴特征值分别为90.3%和2.1%;枯黄期第1排序轴和第2排序轴特征值分别为90.7%和2.2%。
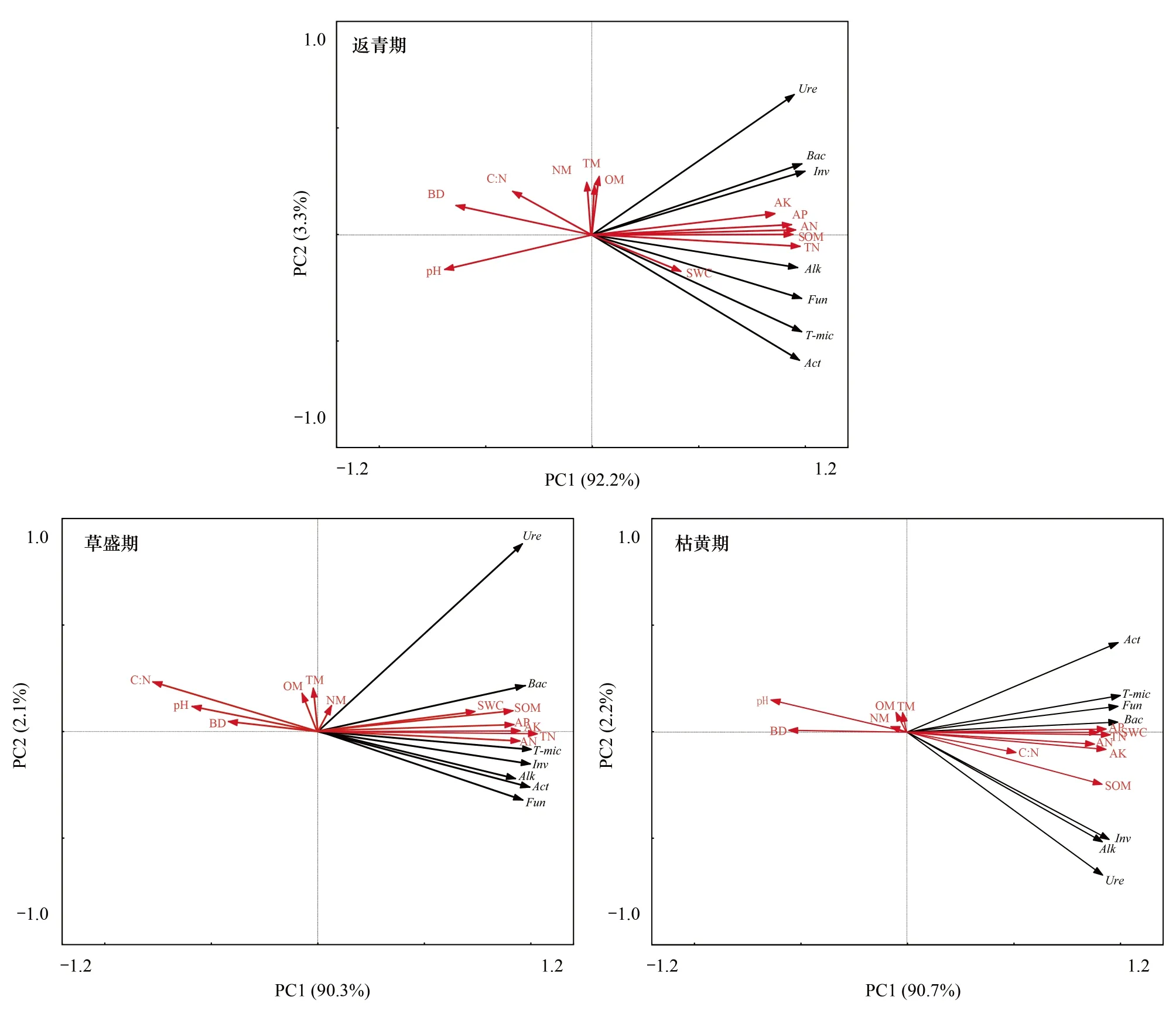
图5 高寒鼠荒地不同植被物候期高原鼢鼠鼠丘数量、土壤理化特性和土壤生物学特征的冗余分析Fig.5 Redundancy analysis on the plateau zoker, soil physicochemical properties and biological characteristics at different vegetation phenological phase in rodent-damaged alpine rangelandpH:pH值;BD:容重; C:N:碳氮比;NM:新土丘数 ; TM:总土丘数;OM:旧土丘数; Ure:脲酶;Bac:细菌;Inv:蔗糖酶;AK:速效钾; AP:有效磷;AN:碱解氮 ; SOM:土壤有机质; TN:全氮;SWC:土壤含水量;Alk:碱性磷酸酶; Fun:真菌; Act:放线菌;T-mic:微生物总数;
对不同土层深度(0—10、10—20、20—30 cm)不同植被物候期的土壤理化性质指标和土壤生物学特征指标的相关性矩阵进行PCA分析(图6)。结果发现高寒鼠荒地不同土层深度下土壤理化性质指标与土壤生物学特征指标在各植被物候期均呈现明显的聚类分布,0—10 cm和10—20 cm土层各指标数量特征在第一轴水平方向上排序为枯黄期>返青期>草盛期,20—30 cm土层各指标数量特征在第一轴水平方向上枯黄期大于草盛期和返青期,但返青期和草盛期间差距不大。说明高寒鼠荒地不同土层深度土壤理化性质指标与生物学特征指标呈现明显的季节性分布。
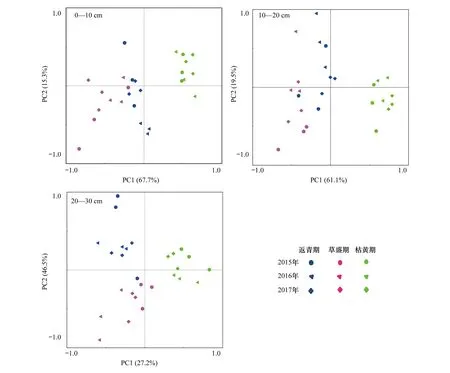
图6 高寒鼠荒地不同土层深度土壤理化性质与土壤生物学特征的主成分分析Fig. 6 Principal component analysis of soil physicochemical properties and biological characteristics at three soil depths in rodent-damaged alpine rangeland
4 讨论
高原鼢鼠掘土造丘将大量下层土壤翻至了地面,使得草地景观、植被群落演替进程发生改变[10,26]。土丘密度往往是指示高原鼢鼠种群数量的重要依据,何俊龄等[27]研究发现以当年草地草盛期(8月)前形成的新土丘密度能较好地表征高原鼢鼠的相对密度。王权业和樊乃昌[10]在青海省门源县研究发现,高原鼢鼠一年中两个活动高峰期分别为4—5月和9—11月,与物候、温度、降雨等均有密切关系。王大伟等[28]在川西北地区的研究发现秋季高原鼢鼠的活动强度显著高于春季,且各个季节鼠丘密度均能较好地反映出高原鼢鼠种群大小。本研究发现,严重退化的高寒鼠荒地上新土丘、旧土丘和总土丘密度植被枯黄期(10月)均大于返青期(5月)和草盛期(8月),草盛期土丘密度最低(新土丘(116±28)个/hm2、旧土丘(1892±208)个/hm2、总土丘(2008±209)个/hm2)(表1)。植被草盛期(8月)平均月降雨量59.1 mm,为3个植被物候期中最低,平均月均温13.36 ℃,为3个植被物候期中最高(图1),可见不同植被物候期的降雨量和温度对高原鼢鼠的鼠丘密度产生了一定的影响,降雨量与鼠丘密度正相关,温度与鼠丘密度负相关[10]。
鼠类造丘活动可以改变草地土壤的空间分布格局,进而改变土壤的理化性质、酶活性和微生物群落特征[29]。Viall等[30]和Wang等[19]研究均发现地下鼢鼠干扰可以增加土壤含水量和有机质含量,从而改善土壤质量,促进土壤的物质能量循环。但高原鼢鼠种群密度超过阈值时,土壤理化性质、酶活性和微生物会发生不可逆转的改变,影响草地的可持续发展[20]。Li等[31]和胡雷等[16]对不同恢复年限土丘土壤采样分析发现高原鼢鼠掘丘和采食行为会对表土性质和土壤有机质产生消极的影响,造成草地土壤有机质、全氮、速效养分等含量显著降低。本研究从鼠丘周围的非土丘区域采样分析发现,高寒鼠荒地不同植被生长季各土层的营养物质分配比例有所变化,植被返青期鼠荒地0—10、10—20、20—30 cm土层分别占总有机质的46%、28%和26%,草盛期分别占到39%、34%和27%,枯黄期分别占到51%、29%、20%(图2),其他指标如全氮、碱解氮、有效磷和速效钾也有类似的规律。由此可见,不同植被生长季高原鼢鼠活动不仅影响了土丘上土壤养分含量的垂直分配[15- 16,30],对非土丘区域土壤养分在鼠荒地的垂直分配也产生了显著的影响[32]。
土壤生物学特征与土壤有机质等营养源的丰富程度、土壤通气状况、水热状况有着密切关系[33]。本研究发现随着土层加深,土壤脲酶、碱性磷酸酶、蔗糖酶、细菌、放线菌、真菌数量以及总微生物数含量均逐渐降低,与孙飞达等[34]在若尔盖花湖景区的研究结果一致。从植被返青期至枯黄期,土壤表层(0—10 cm)各生物学特征指标含量显著提高,下层土壤(10—20 cm和20—30 cm)生物学特征含量的变化不太显著(表2、图3、图4),原因可能是返青期5月平均月均温为6.64 ℃(图1),土壤还未完全解冻,导致土壤生物学特征含量均处在较低水平;草盛期8月份虽然温度较高,但鼢鼠活动强度下降,土壤通气状况较差,降雨量也较低,仅59.10 mm,为3个植被物候期中最低,土壤生物学特征含量均处相较5月份持续升高;枯黄期10月份,鼢鼠活动强度上升,土壤温度、湿度较高,导致土壤脲酶、碱性磷酸酶、蔗糖酶、细菌、放线菌、真菌数量以及微生物总数均处在高位,这些因素对土壤表层生物学特征含量的影响更大[32,35]。
土壤生物学特征对生态系统改变非常敏感,与环境因子关系密切[36]。RDA分析发现各植被物候期土壤理化性质和土壤生物学特征间均有显著的相关性,土壤pH值、容重与土壤生物学特征指标呈显著负相关,土壤有机质、全氮、碱解氮、有效磷、速效钾与土壤生物学特征含量呈显著正相关(图5),与胡雷等[34]在三江源地区的研究一致。土壤碳氮比在植被返青期和草盛期与土壤生物学特征含量呈负相关,枯黄期与土壤生物学特征含量负相关,原因主要是枯黄季土壤有机质及全氮的垂直分配变化导致[32]。PCA分析发现,各土层土壤在第一轴水平方向上呈明显的聚类分布(图6),表明虽然植被物候期改变了土壤理化性质和生物学特征含量的垂直分配,但不同土层指标含量呈现明显的季节性分布。0—10 cm和10—20 cm土层各指标数量特征在第一轴水平方向上排序为:枯黄期>返青期>草盛期,20—30 cm土层各指标数量特征在第一轴水平方向上枯黄期大于草盛期和返青期,但返青期和草盛期间差距不大。可见,植被枯黄期由于高原鼢鼠活动加剧对非土丘区域土壤表层的理化性质和生物学特征指标均产生了显著的影响。
