瞬时遗传转化长白落叶松NAC基因植株抗旱性的研究
2020-06-03熊欢欢赵佳丽张含国
张 磊 熊欢欢 曹 庆 赵佳丽 张含国
(东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040)
由于全球气候的波动变化,各种极端气候频发,我国春旱灾害时有发生,严重限制了各地林业生产的发展。随着生物技术尤其是基因工程技术的发展,物种间的遗传界限逐步被打破,使得各类育种工作能够通过基因工程的手段精准、便捷、快速地实现育种目标。利用生物技术或基因工程对林木进行遗传改良的基础是遗传转化体系及对基因功能的了解[1~4],这对开展落叶松分子育种十分必要。
NAC类转录因子是植物中特有的一类转录因子,多数研究表明NAC在植物抗逆和防卫反应中起到了重要作用[5~6]。长白落叶松作为我国东北地区主要针叶造林树种,其基因功能研究较少,通过长白落叶松瞬时转化NAC基因后对干旱胁迫生理指标的变化可以推断基因在抗逆方面是否具有一定的功能,为后期培育落叶松抗旱新品系提供优良的基因,为加速落叶松分子育种进程奠定基础。
1 材料与方法
1.1 试验材料
选取饱满的长白落叶松种子,用去离子水浸泡4~5 d,期间换水3~4次,播种于营养土基质(粘土、蛭石、珍珠岩比例为5∶3∶2)中,1.0~1.5个月后,取针叶未完全展开的落叶松幼苗利用构建好的pCAMBIA1301植物表达载体进行瞬时侵染,获得LoNAC18瞬时遗传转化的长白落叶松植株用于胁迫处理。
1.2 材料处理
配置30%(w/v)的PEG溶液,在瞬时遗传转化植株栽植在营养土中12 h后(记为T0)开始浇灌,同时以瞬时转化空载体植株作为对照,分别于胁迫后0 h(T0),24 h(T24),48 h(T48)取整株材料作为试验样品,置于去离子水中洗去表面杂质,吸干水分,液氮速冻后于-80℃冰箱中保存,提取RNA待用。
1.3 序列分析
将得到的基因序列利用NCBI进行分析,确定保守结构域,然后利用ExPASy提供的在线软件ProtParam(http://web.expasy.org/protparam/)对氨基酸序列进行一级结构预测,包括氨基酸长度、分子质量、理论等电点,利用网页在线工具GOR4和SwissModle(网址为https://www.swissmodel.expasy.org/)分别预测其二级结构与三级结构。
1.4 RT-PCR
以长白落叶松α-tubulin基因为内参基因,根据基因序列设计定量引物序列为(5′—3′)为AAGCTGCAACTGCCCAAACT和TGCTGCTGTTTCCATTACCA。荧光定量PCR反应体系为2×TransStart Top Green qPCR SuperMix 10.0 μL,10 μmol·L-1F和R引物各1.0 μL,Passive ReferenceDye(50×)(optinal)1.0 μL ,cDNA模板1.0 μL,加水补至20.0 μL。采用2-ΔΔCt方法进行数据分析。
1.5 生理指标测定
分别于胁迫后0 h(T0),24 h(T24),48 h(T48)取整株材料作为试验样品,进行研磨,参照可溶性糖含量、丙二醛含量(malondiadehyde,MDA)、可溶性蛋白含量、过氧化物酶(Peroxidase,POD)活性、超氧化物歧化酶(Superoxide Dismutase,SOD)活性测定试剂盒(苏州科铭生物技术有限公司)进行测定。
2 结果与分析
2.1 NAC基因序列分析
根据前期长白落叶松在干旱胁迫条件下的转录组数据分析获得该条NAC全长基因序列,命名为LoNAC18,对该基因进行基本分析结果表明:全长基因序列1 101 bp,具有NAC基因家族的NAM保守结构域。与拟南芥的NAC基因家族序列比对结果表明,相似度最高的是AtNAC018和AtNAC056。
利用Protparam蛋白的理化性质及亲水性进行预测分析发现LoNAC18为不稳定的亲水性蛋白,其二级结构表明蛋白以随机卷曲为主,含有17.21%的α-螺旋,21.31%的延伸链以及61.48%的无规则卷曲(如图3所示)。利用同源建模法预测的三级结构(如图4所示)。
2.2 瞬时转化植株的验证
经过RT-PCR检验基因表达量发现,瞬时遗传转化的长白落叶松过表达幼苗的NAC基因表达量有显著性增加。说明通过瞬时遗传转化后基因的表达量发生变化,可用于基因功能的初步研究。在PEG模拟干旱胁迫的条件下,NAC基因表达量从胁迫0 h到胁迫48 h呈上升趋势,且在胁迫48 h达到最大值。转化植株在PEG胁迫条件下的基因表达量较未胁迫的同一时期的植株发生上调,说明NAC基因可以被PEG模拟的干旱胁迫诱导表达,NAC基因可能参与调控长白落叶松对干旱胁迫的应答反应。

表1 LoNAC18氨基酸序列理化性质分析

图1 NAC基因的保守结构域分析Fig.1 NAC conserved domain database
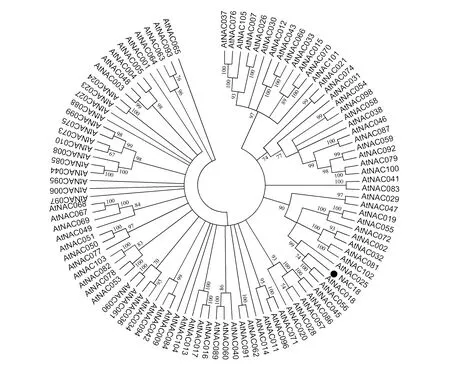
图2 长白落叶松NAC基因与拟南芥NAC基因家族的序列比对Fig.2 NAC Sequence alignment between Larix olgensis and Arabidopsis thaliana family
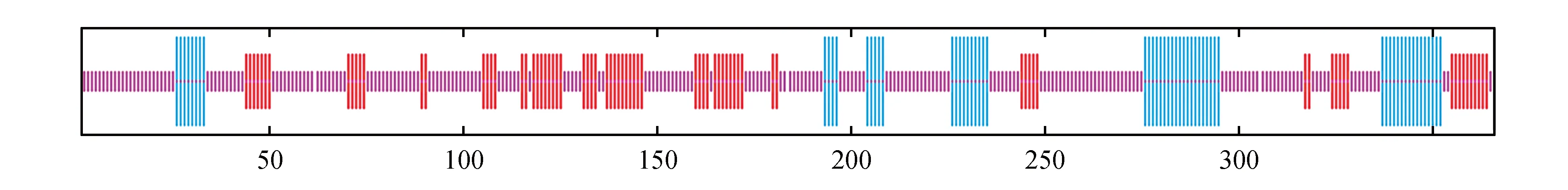
图3 长白落叶松蛋白质的二级结构 竖长线(蓝色)代表螺旋;竖中线(红色)代表折叠;竖短线(紫色)代表卷曲Fig.3 Secondary structure of larch protein Vertical line(blue) represents spiral; Vertical midline(red) represents folding; Short vertical lines(purple) represent curls

图4 长白落叶松蛋白质的三级结构Fig.4 Tertiary structure of larch protein
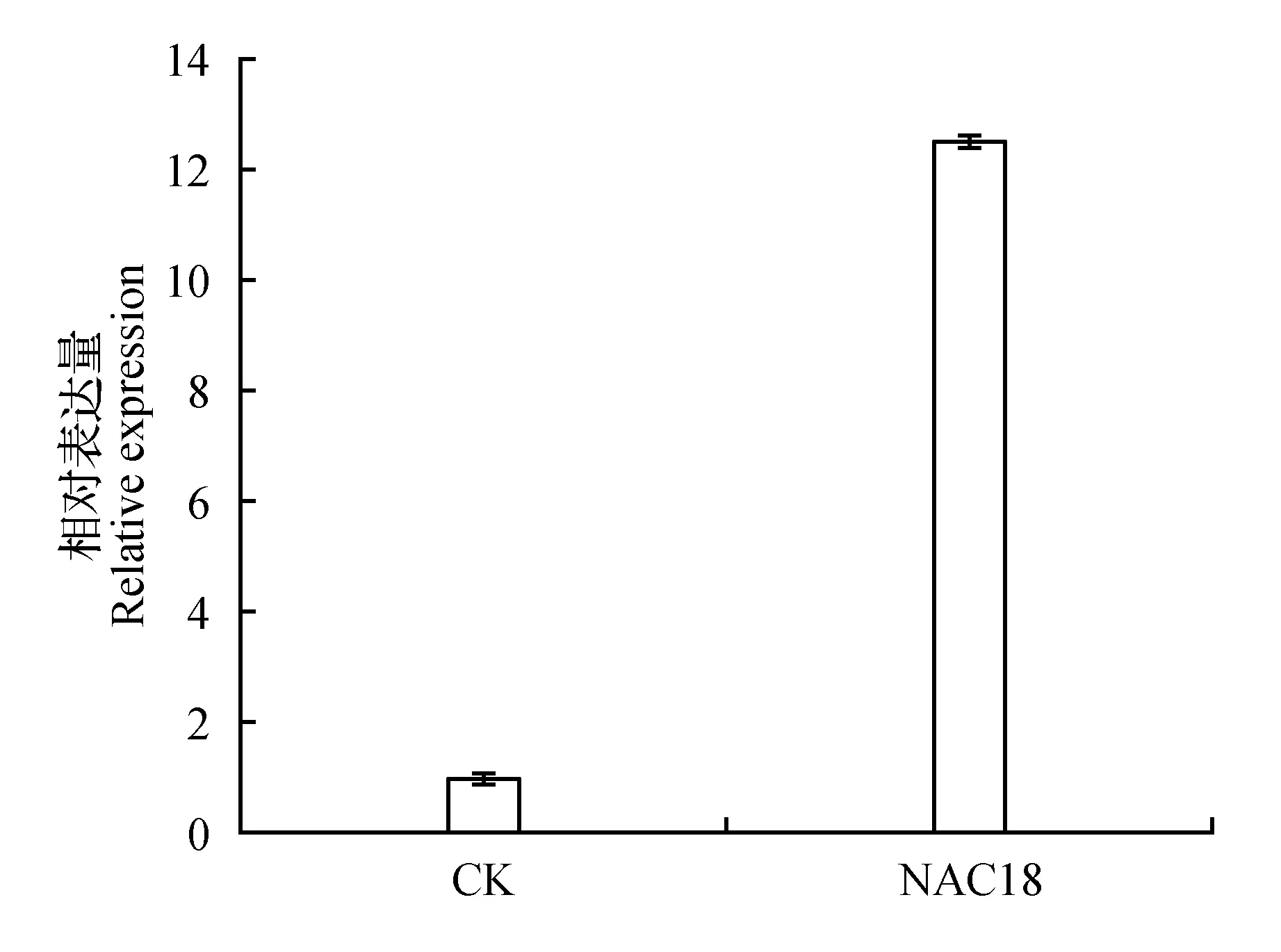
图5 瞬时转化植株NAC基因表达量分析Fig.5 Analysis of NAC gene expression in transient transformed plants
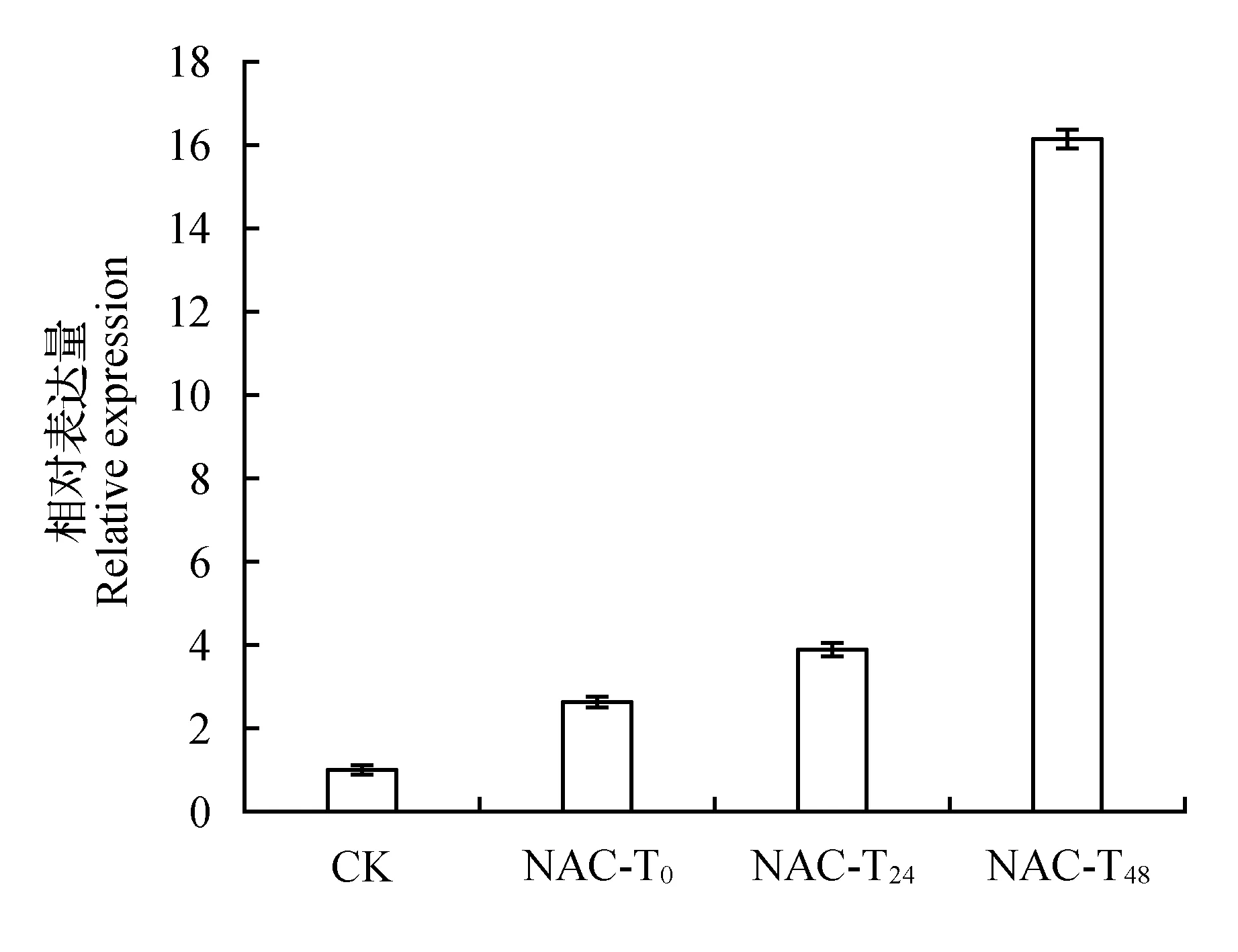
图6 PEG模拟干旱胁迫瞬时转化植株NAC基因表达量分析Fig.6 NAC gene expression of transformed plants under PEG stress
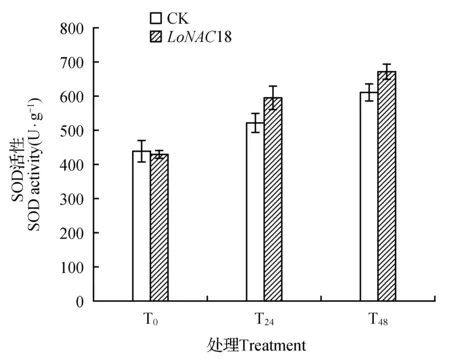
图7 瞬时转化植株在干旱胁迫下SOD活性的变化Fig.7 SOD activity in transient transformed plants under drought stress
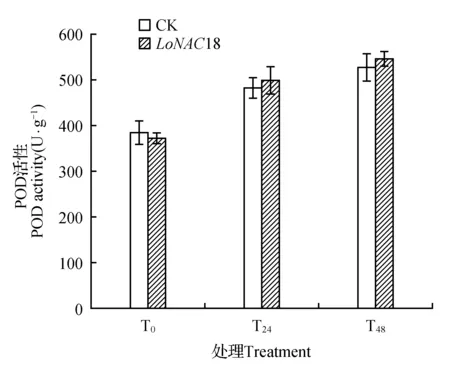
图8 瞬时转化植株在干旱胁迫下POD活性的变化Fig.8 POD activity in transient transformed plants under drought stress
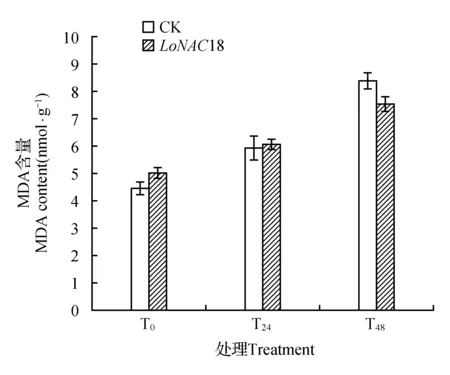
图9 瞬时转化植株在干旱胁迫下MDA含量的变化Fig.9 MDA content in transient transformed plants under drought stress
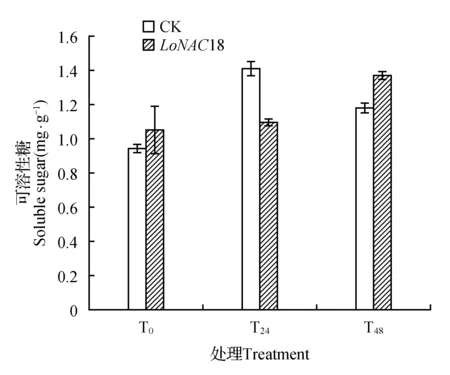
图10 瞬时转化植株在干旱胁迫下可溶性糖含量的变化Fig.10 Soluble sugar content in transient transformed plants under drought stress
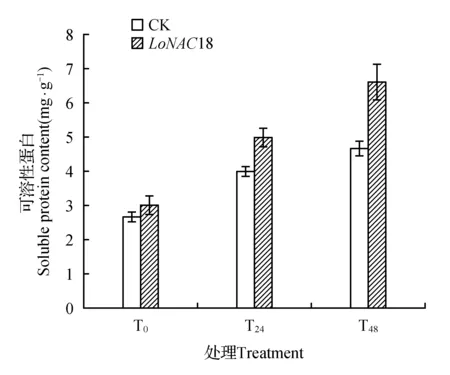
图11 瞬时转化植株在干旱胁迫下可溶性蛋白含量的变化Fig.11 Soluble protein content in transient transformed plants under drought stress
2.3 瞬转植株在PEG胁迫下的生理指标变化
2.3.1 超氧化物歧化酶(SOD)活性
当植物受到生物或者非生物性逆境胁迫时,植物中的抗氧化保护酶系统过度表达。其中SOD可以调节细胞膜膜质之中的氧化反应,与POD在植物的抗逆性生长发育中起着重要的作用,可以提高植物的抗逆性。
长白落叶松植株瞬时转化NAC基因后,在干旱胁迫条件下SOD酶活性呈现上升趋势(如图7所示)。
与对照植株中的活性相比,在干旱胁迫开始后瞬时转化植株的SOD酶活性均高于对照,且在48时达到最大值。瞬时转化NAC基因的长白落叶松植株SOD活性与未胁迫时相比,在24 h增加38.46%,48 h增加56.33%。T24和T48胁迫后的转基因植株与同时期对照相比,SOD活性分别为13.85%和10.37%。
2.3.2 过氧化物酶(POD)活性
POD是过氧化物酶,可以催化氢氧化物和过氧化氢反应转化为醌类且可以促进植物木质化。干旱胁迫之下,长白落叶松植株之中的POD活性随着胁迫时间的增长而逐渐增加(如图8所示),且增加幅度均大于对照,但差异不明显。
瞬转NAC基因的长白落叶松植株POD活性在胁迫48 h后达到最大,与未胁迫时相比,T48比T0和T24样品的POD活性分别提高46.95%和9.62%。而胁迫24和48 h的瞬时转化植株与对照相比分别只提高了3.31%和3.27%。
2.3.3 丙二醛(MDA)含量
MDA是植物在遭遇逆境胁迫下产生的,其在植物体内的含量随逆境胁迫时间和胁迫程度的变化而变化。因此,MDA也可以作为研究植物生理和生态变化的一项重要生理指标。
从胁迫开始,对照植株与瞬转植株的MDA含量均呈现上升趋势(如图9所示),表明所有植株都受到干旱胁迫的影响,体内的MDA含量逐渐增加。
在胁迫开始时和胁迫24 h时,瞬时转化NAC的落叶松植株的MDA含量大致相同,没有出现显著差异。在胁迫开始后,对照植株与瞬转植株的MDA含量均大幅度上升,在48 h到达最大值,但瞬时转化植株MDA上升幅度小于对照植株。对照植株的胁迫48 h与胁迫24 h的MDA含量与未胁迫时相比分别增加了87.62%和33.56%,而同时期瞬转NAC植株的MDA含量与未胁迫相比增加了50.12%和20.28%。所以瞬转植株累计的MDA含量明显小于对照植株,说明其受到胁迫影响较小。
2.3.4 可溶性糖含量
可溶性糖储存能量并为植物的生长发育做能量准备。植物在遭受干旱、高温等逆境胁迫时,其体内的可溶性糖含量会增加,植物的抗逆机制将被触发,以防止植物受到更多的损害。
在干旱胁迫下,对照植株的可溶性糖含量呈现先上升后下降的趋势,而瞬转植株的可溶性糖含量在胁迫24 h之内几乎没有变化,在胁迫48 h时显著增加(如图10所示)。
在胁迫24 h,对照的可溶性糖含量分别比对照和胁迫48 h高出49.95%和19.82%。胁迫48 h的瞬转植株可溶性糖含量与T0和T24相比分别高出30.82%和25.03%,相比同时期对照可溶性糖含量高出16.65%。
2.3.5 可溶性蛋白含量
可溶性蛋白作为植物细胞中重要的渗透调节物质和营养物质,其含量的增加和积累能提高细胞的保水能力,对细胞的生命物质及生物膜起到保护作用。因此,可溶性蛋白的含量也可以作为测定植物抗逆能力的一项重要指标。
在干旱胁迫下,瞬转植株与对照植株的可溶性蛋白质含量随着胁迫时间的增长而不断上升(如图11所示)。与对照相比,瞬转植株的可溶性蛋白含量上升趋势高于对照。瞬转植株胁迫24和48 h的可溶性蛋白含量与未胁迫时相比分别提高66.89%和120.81%,与对照同时期相比分别高出9.2%和30.3%。而瞬转LoNAC8的植株胁迫24和48 h的可溶性蛋白含量与未胁迫时相比分别提高72.3%和147.6%,与对照同时期相比分别高出25.14%和41.51%。
3 讨论
当植物受到干旱胁迫时,一般会通过减少细胞内的水分含量、缩小细胞的体积、增加细胞内可溶解物质的含量等途径,降低细胞渗透势,防止细胞过度失水,从而维持植物的正常生命活动[7~8]。NAC类转录因子是植物中所特有的,有研究指出NAC转录因子参与了植物的抗逆和防卫反应[6]。比如在柑橘[9]、毛竹等均发现了能够响应干旱胁迫的NAC转录因子家族[10]。可溶性糖含量、可溶性蛋白含量、MDA含量、SOD活性、POD活性均呈现上升趋势是植物在受到干旱胁迫后的普遍变化[11~13]。
Tran等[14]证明拟南芥的ANAC019,ANAC055和ANAC072基因的表达受干旱、高盐和ABA诱导,超表达这3个基因能够增强转基因植株的耐旱能力。Hu等[15]发现水稻中的SNAC1基因能够被干旱和盐胁迫所诱导,将该基因在水稻中超表达可以明显提高转基因植株对干旱、高盐的耐受性。还有大量的研究证实柑橘中的NAC83[9]、毛竹中的PeNAC1~PeNAC7等均能够响应干旱胁迫[10],在胁迫后被诱导高表达时基因参与调控的显著特征之一。有研究表明拟南芥NAC018蛋白及其编码基因具备耐干旱和耐盐性及延缓衰老的作用[16],LoNAC18与拟南芥NAC018基因序列相似度最高,推测其具有类似的功能或作用。
为初步验证长白落叶松NAC基因是否参与调控长白落叶松对干旱胁迫的应答反应,对瞬时转化后的植株进行PEG干旱胁迫,分析转化后植株在不同时间的基因表达量变化,结合转化后植株的可溶性糖、可溶性蛋白、SOD、POD、MDA等生理生化指标分析。分析结果可知,瞬时转化NAC基因的长白落叶松在PEG模拟干旱胁迫条件下,NAC基因表达量上调,与对照相比,SOD和POD活性都有所增加,MDA含量累计程度低于对照,可溶性蛋白含量增加,可溶性糖含量在胁迫48 h时高于对照,说明NAC基因在一定程度上参与调控了长白落叶松应答PEG模拟干旱胁迫的过程,其参与调控的代谢通路有待进一步研究。
