抚育择伐对马尾松林凋落叶分解速率和养分释放的短期影响*
2020-06-02王晓荣曾立雄肖文发
王晓荣 雷 蕾 付 甜 潘 磊 曾立雄 肖文发
(1.中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室 北京 100091;2.南方现代林业协同创新中心 南京 210037;3.湖北省林业科学研究院 武汉 430075)
凋落物分解是森林生态系统物质循环和能量流动的重要环节,是维持生态系统功能的主要过程之一(刘瑞强等,2015;林成芳等,2017),在维持林地土壤肥力、提供植物生长发育的必要养分、促进全球养分循环等方面具有举足轻重的作用(Kunhamuetal., 2009;Chenetal., 2014)。凋落物分解过程受基质质量、分解者种类和数量、物理环境等因素的综合影响(Navarroetal., 2013;葛晓改等,2015),一直以来都是生态学研究的热点。
抚育择伐是常见的森林经营管理方式(Navarroetal., 2013),往往会使林分结构、树种组成、土壤环境、微生物种类和活性、凋落物数量和质量发生改变(王意锟等,2011;梁晶等,2015),并对林内光有效性、大气温度、土壤温湿度等微环境产生影响(林娜等,2010;Chenetal., 2014),进而共同影响凋落物分解速率和周转以及养分矿化速率(Kunhamuetal., 2009;Navarroetal., 2013)。已有研究发现,适度择伐可改变凋落物层温度、酶活性、木质素/N、C/N等,加速凋落物分解,促进养分归还给土壤;而过度择伐则会抑制凋落物分解,阻滞养分的回归和利用(李国雷等,2008;肖文娅等,2016)。然而,研究区域、森林类型、择伐时间、择伐方式和强度不同(Titusetal., 2006; Kunhamuetal., 2009;De Longetal., 2016),往往会造成不同营林方式和干扰程度对凋落物分解的巨大差异(肖文娅等,2018)。有报道指出,凋落物基质质量与生物交互作用会导致分解速率和养分周转更加复杂(Schmidtetal., 2011),而抚育择伐会直接改变林分结构和生物环境,加剧凋落物分解和生态系统养分循环的复杂程度。以往关于抚育择伐对凋落物分解的影响主要基于抚育择伐强度来开展,而对选择抚育择伐对象考虑较少,此方面研究也鲜见报道。
马尾松(Pinusmassioniana)是我国亚热带地区重要的造林树种之一,分布广、适应性强,以马尾松为优势种的针叶林是三峡库区分布面积最大的森林类型(雷蕾等,2016),在库区生态安全中发挥着重要作用。目前,由于长期受当地农民采伐大径材马尾松、砍伐杂木以及林下植被抚育等粗放经营影响,库区马尾松林林分质量、稳定性和生态系统功能较差(Leietal., 2018),如何采取科学合理的营林措施提升低效马尾松林林分质量和生态系统功能,已成为库区森林可持续发展的核心问题。而研究不同植被功能群择伐后的林分结构变化对凋落物分解的影响并确定主要影响因子,对完善森林抚育间伐理论和指导现实生产具有重要意义。鉴于此,本研究以三峡库区马尾松林为对象,设置未择伐、除灌、伐除非马尾松和伐除优势马尾松4种抚育择伐处理,分析不同抚育择伐处理对马尾松林凋落叶分解速率和养分释放的影响,以期为马尾松生态公益林可持续经营提供科学依据。
1 研究区概况
研究区位于湖北省秭归县九岭头林场(110°45′—110°48′E,30°59′—31°05′N),海拔156~2 030.6 m。属亚热带大陆性季风气候,年均气温17.9 ℃,年降水量1 000~1 250 mm,多集中在4—9月,年日照时数1 619.6 h,年无霜期260天。土壤类型以黄壤、黄棕壤为主(王娜等,2017a)。该区马尾松林为20世纪70年代飞播造林,长期以来人为干扰极少,以马尾松为优势种,郁闭度约0.7,伴生树种包括光皮桦(Betulaluminifera)、漆树(Toxicodendronvernicifluum)、茅栗(Castaneaseguinii)和盐肤木(Rhuschinensis)等。灌木主要有绿叶胡枝子(Lespedezabuergeri)、桦叶荚蒾(Viburnumbetulifolium)和宜昌荚蒾(V.erosum)等,草本以中日金星蕨(Parathelypterisnipponica)、狗脊(Cibotiumbarometz)、鱼腥草(Heartleafhouttuynia)、野青茅(Deyeuxiaarundinacea)和细梗苔草(Carexteinogyna)为主。
2 研究方法
2.1 试验设计
2013年,在九岭头林场选择同一坡面生长且林相完整的马尾松林,随机布设16块20 m×20 m样地。设置未择伐(保持林分现状,无人为干扰)、除灌(清除林内灌木层和阔叶更新层,保留马尾松更新小树)、伐除非马尾松(伐除胸径≥4 cm的非马尾松树种,择伐强度为15%)和伐除优势马尾松(伐除树木平均胸径以上的马尾松个体,择伐强度为70%)4种抚育择伐处理,每处理4次重复。2016年,对样地林分内胸径≥4 cm的木本植物进行统计,包括物种名、胸径、数量及郁闭度等指标。样地基本情况如表1所示。
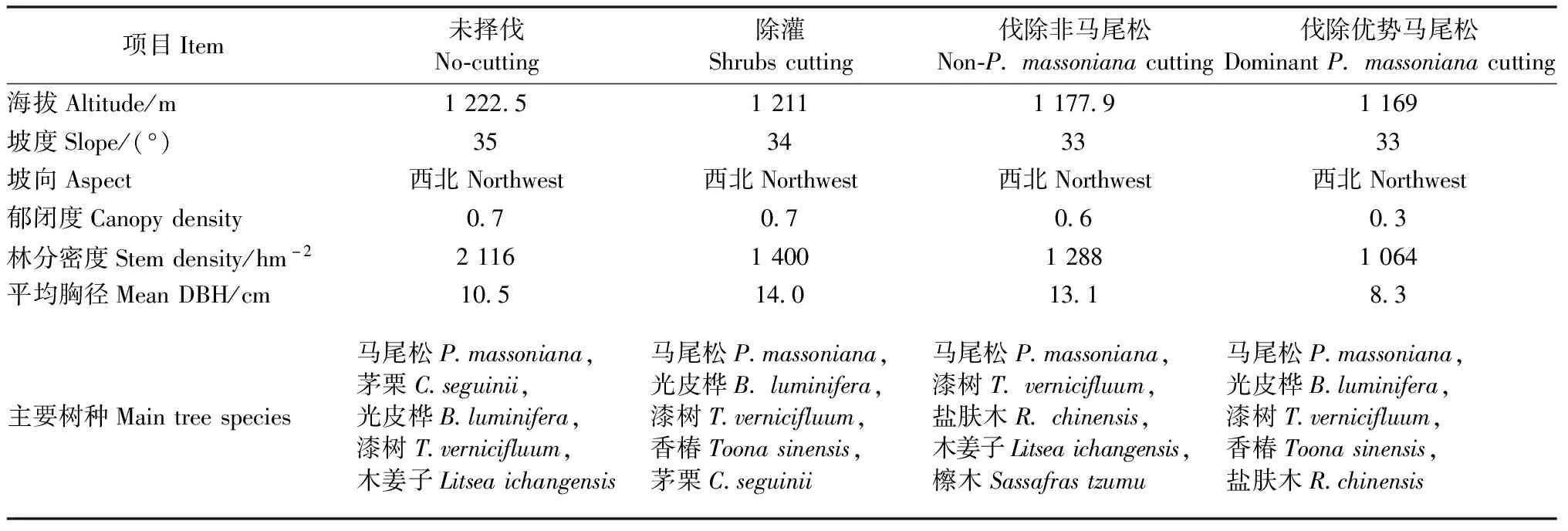
表1 样地2016年的基本情况
2016年11月初,原位收集各处理样地内新近自然凋落的马尾松叶,带回实验室65 ℃烘干至恒质量,将同一处理样地内马尾松林凋落叶充分混合,称量10 g装入规格为20 cm×20 cm、孔径1 mm的尼龙网袋,封口并标记。随后,将每块20 m×20 m样地划分为4个10 m×10 m分解小区,在每个分解小区内随机各放置4个尼龙网袋,每处理共计64次重复,4种处理共放置256袋(4处理×4重复×4个小区×4个尼龙袋)。将土壤表面凋落物去掉,分解袋紧贴地表放置,并用木钉固定,以模拟自然状态凋落物分解,自放置之日起,每隔3个月从每个分解小区随机收集1袋,每次共收集64袋,共收集4次,收集日期分别为2017年2月15日、5月15日、8月15日和11月15日。凋落叶袋取回后小心清除袋内的土壤和杂质,65 ℃下烘干至恒质量后称干质量,以计算凋落叶残留率。
在凋落叶样品装入分解袋前,对充分混合好的新近自然凋落的马尾松叶,每处理随机挑选4份作为凋落叶初始养分含量的分析样品。在分解过程中,将每块20 m×20 m样地的4个分解样混合为1份,共计16个样品。样品粉碎并过0.25 mm筛,待化学分析。C含量用重铬酸钾氧化法测定;N含量用H2SO4-H2O2法消煮-靛酚蓝比色法测定;P含量用H2SO4-H2O2法消煮-钼锑抗比色法测定。利用HOBO微型气象监测系统记录各处理样地林内距地表1.3 m高处空气温度和湿度以及5 cm深处土壤温度和水分含量(土壤体积含水量),记录间隔时间1 h,每块样地安装1套微型气象监测系统。
2.2 数据分析
凋落叶分解速率采用Olson指数衰减模型计算,公式如下:
Xt/X0=ae-kt。
(1)
式中:Xt表示分解时间t后的凋落叶干质量(g);X0表示凋落叶初始样品干质量(g);k为凋落叶年分解速率(g·g-1a-1);t为分解时间(a);a为拟合参数。
凋落叶分解50%和95%所需时间计算公式如下:
t0.5=ln0.5/(-k);
(2)
t0.95=ln0.05/(-k)。
(3)
式中:t0.5表示凋落物分解50%所需时间(a);t0.95表示凋落叶分解95%所需时间(a)。
凋落叶分解过程中养分释放率Et公式为:
Et=(X0C0-XtCt)/X0C0×100%。
(4)
式中:Et为分解过程中凋落叶C、N、P释放率(%);X0为凋落叶初始质量(g);Xt为分解t时刻剩余凋落叶质量(g);C0为凋落叶初始元素含量;Ct为分解t时刻剩余凋落叶元素含量。
采用SPSS19.0和Excell13.0软件对数据进行统计分析。凋落叶分解速率、养分含量的差异性采用单因素方程分析(one-way ANOVA),且进行LSD多重比较,以检验数据之间的差异显著性(P≤0.05)。为进一步了解不同抚育择伐处理中影响马尾松林凋落叶分解的主导因素,利用多元逐步回归分析方法构建分解速率与凋落叶初始C、N、P、C/N、C/P、N/P及5 cm深处土壤温度和水分含量等因子的模拟关系式。采用Sigmplot10.0软件进行作图。
3 结果与分析
3.1 空气温湿度及5 cm深处土壤温度和水分含量变化
由图1可知,不同抚育择伐处理间林内空气温度和湿度均不存在显著差异(林内空气温度:F=0.204,P=0.894;林内空气湿度:F=0.527,P=0.664);5 cm深处土壤温度也无显著差异(F=0.257,P=0.856);但5 cm深处土壤水分含量存在极显著差异(F=19.129,P<0.001),表现为未择伐>除灌>伐除非马尾松>伐除优势马尾松,除灌、伐除非马尾松和伐除优势马尾松土壤水分含量分别较未择伐降低0.80%、5.98%和7.57%。土壤温度和土壤水分含量具有相反的变化趋势。
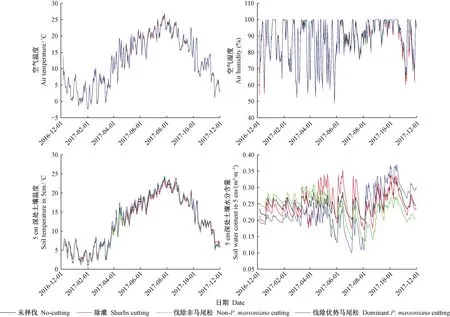
图1 不同抚育择伐处理下林内空气温湿度及5 cm深处土壤温度和水分含量
3.2 抚育择伐对马尾松林凋落叶初始养分含量的影响
由表2可知,不同抚育择伐处理间初始C、N含量和C/N存在极显著差异(C含量:F=7.114,P=0.005;N含量:F=7.831,P=0.004;C/N:F=13.008,P=0.001),N/P存在显著性差异(F=4.070,P=0.033),而P含量和C/P无显著差异(P含量:F=2.296,P=0.130;C/P:F=2.047,P=0.161)。初始C含量表现为伐除优势马尾松>除灌>未择伐>伐除非马尾松,N含量表现为未择伐>除灌>伐除非马尾松>伐除优势马尾松,C/N表现为伐除优势马尾松>伐除非马尾松>除灌>未择伐,N/P表现为除灌>伐除非马尾松>未择伐>伐除优势马尾松。由此可知,不同抚育择伐对马尾松林凋落叶初始养分含量具有显著影响,其中除伐除非马尾松处理C含量降低外,其余处理C含量均明显增加,同时抚育择伐降低了凋落叶初始N、P含量,增加了C/N和N/P。
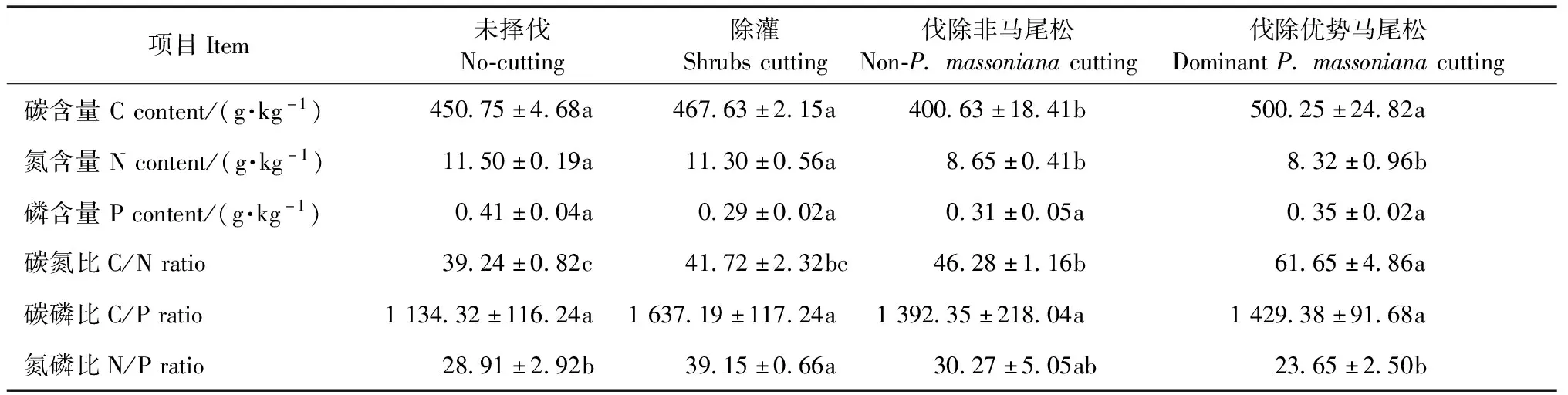
表2 不同抚育择伐处理下马尾松林凋落叶初始养分含量①
①同行不同小写字母表示差异显著(P<0.05)。Different letters in the same row indicated significant difference at 0.05 level.
3.3 凋落叶分解动态
不同抚育择伐处理马尾松林凋落叶残留率均随分解时间增加呈逐渐降低趋势(图2),且处理和分解时间对马尾松林凋落叶分解速率均存在极显著影响(P<0.01)(表3)。分解12个月后,不同抚育择伐处理凋落叶残留率表现为未择伐(73.06%±1.05%)>伐除优势马尾松(72.59%±0.69%)>伐除非马尾松(71.92±1.40%)>除灌(66.41%±1.13%),均可促进凋落叶分解,且以除灌处理的凋落叶分解最快。采用Olson指数衰减模型拟合分解速率,各处理拟合方程模拟精度均达到显著水平(表4),不同抚育择伐处理马尾松林凋落叶年分解速率表现为除灌(0.397 g·g-1a-1)>伐除优势马尾松(0.315 g·g-1a-1)>伐除非马尾松(0.311 g·g-1a-1)>未择伐(0.304 g·g-1a-1)。未择伐、除灌、伐除非马尾松和伐除优势马尾松处理的t0.5分别为2.28、1.74、2.23和2.20年,t0.95分别为9.85、7.55、9.63和9.51年。可见,抚育择伐处理均在一定程度上提高了马尾松林凋落叶的分解速率,但各处理间存在明显差异。
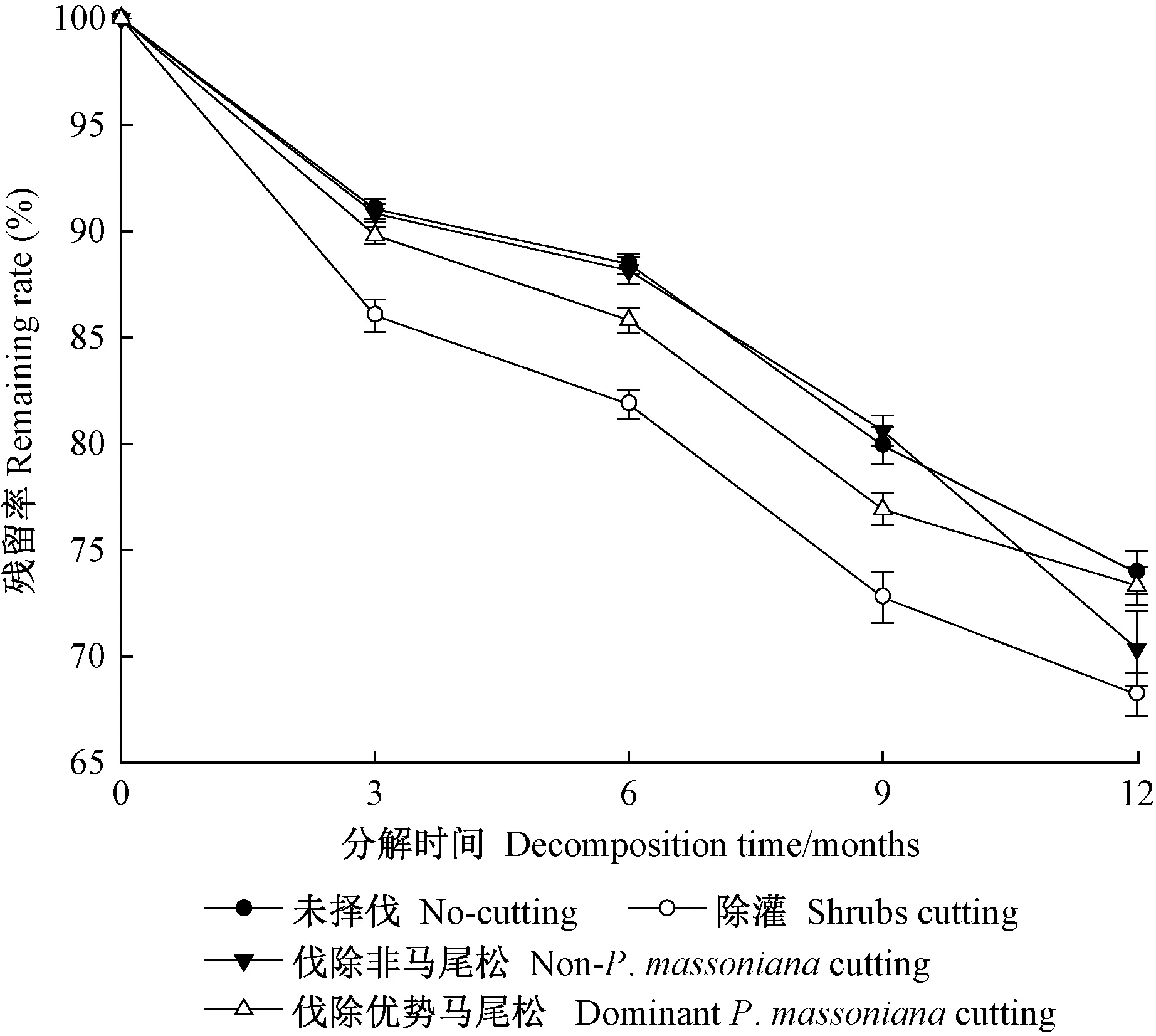
图2 不同抚育择伐处理下马尾松林凋落叶分解动态

表3 分解速率和养分释放率的双因素方差分析(F值)①
①* :P<0.05;**:P<0.01.
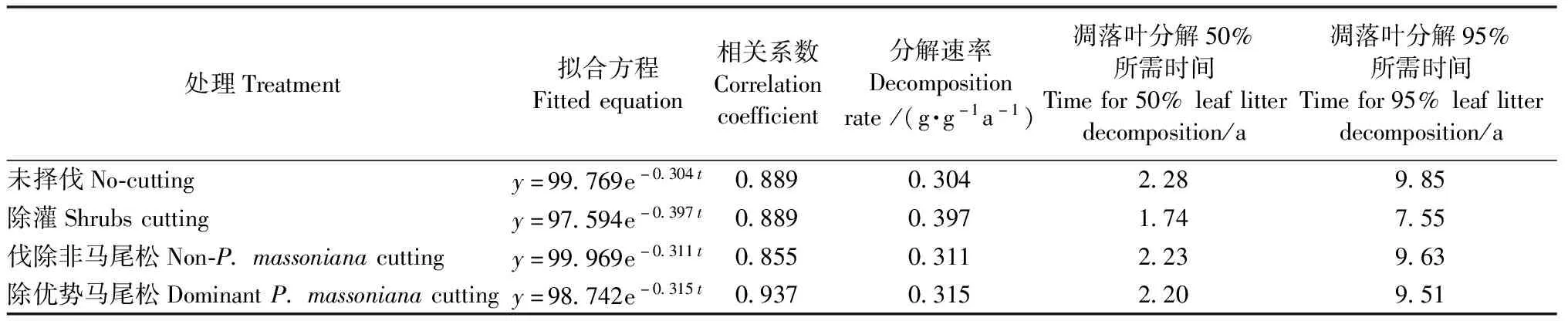
表4 马尾松林凋落叶分解速率方程及分解参数

图3 不同抚育择伐处理下马尾松林凋落叶分解过程中C、N、P含量和C/N、C/P、N/P变化
3.4 凋落叶分解过程养分释放动态
由表3可知,在不同处理和分解时间之间,凋落叶C和N释放率存在极显著差异(P<0.001),但P释放率则无显著差异(P>0.05)。图3表明,不同抚育择伐处理马尾松林凋落叶C含量均先逐渐降低后升高再降低;分解12个月后,未择伐、除灌、伐除非马尾松和伐除优势马尾松处理C含量较各自初始C含量降低50.50%、52.53%、43.67%和51.35%;各处理N含量均在前3个月呈减少状态,随后呈持续增加状态,分解结束后较初始N含量分别增加140.61%、144.33%、92.82%和188.34%;各处理P含量在前3个月呈减少状态,而后9个月内变化趋势各不相同,分解12个月后未择伐、除灌和伐除优势马尾松处理凋落叶P含量分别较初始P含量降低31.71%、44.83%和48.57%,而伐除非马尾松处理则较初始P含量增加了45.13%;各处理凋落叶C/N在分解期内均呈先升高后持续降低趋势,分解结束时C/N分别较初始C/N降低了77.77%、77.52%、69.71%和54.94%,且择伐强度越大下降越明显;各处理凋落叶C/P仅在分解初期表现为增加,后9个月则无规律性变化,可能与生长季中雨水对P元素的淋洗造成波动有关;N/P在分解前6个月内变化很小,与初始比率相差无几,在随后6个月内除伐除非马尾松处理仍没有显著变化外,其余处理均呈增加趋势。
3.6 影响凋落叶分解的因素分析
马尾松林凋落叶分解速率与初始养分含量以及分解期间5 cm深处土壤温度和土壤水分含量的相关性分析表明,分解速率与初始N含量和N/P显著正相关(P<0.05),而与其他初始养分含量和土壤温湿度均无显著相关关系(P>0.05)。多元逐步回归分析拟合方程为k=0.222+0.004x1(R2=0.324,P=0.021),式中k为分解速率,x1为N/P。从方程系数可知,凋落叶初始N/P是影响马尾松林凋落叶分解速率的关键性因素。
4 讨论
4.1 不同抚育择伐处理对马尾松林凋落叶分解速率的影响
采取适度抚育择伐,可通过改变林分结构、物种组成、凋落物基质质量、土壤温湿度条件、微生物群落活性和结构等因素,加快凋落物分解,缓解过分积累问题(李国雷等,2008;林娜等,2010;肖文娅等,2016;2018;Titusetal., 2006;梁晶等,2015)。本研究发现,除灌、伐除非马尾松和伐除优势马尾松处理均在一定程度上促进了马尾松林凋落叶分解,这与以下两方面原因有关。一方面,抚育择伐可改变林分密度和林分覆盖度,增加林地地表光照、蒸散增强,导致地表温度的同时降低了土壤含水量,从而直接影响微生物的结构、功能和酶活性,进而显著影响凋落物分解速率(Laihoetal., 2004;刘华等,2017)。本研究中,相较未择伐,除灌、伐除非马尾松和伐除优势马尾松导致林内5 cm深处土壤温度分别升高0.11、0.20和0.39 ℃,土壤水分含量分别下降0.01、0.06和0.08 m3·m-3,且土壤温湿度变化随抚育择伐强度增大而增加,但凋落物分解速率与土壤温湿度间相关性并不显著,说明对于局域尺度上人为抚育择伐产生的林内水热条件的小幅度变化还不足以成为影响凋落物分解速率的决定性因素(李国雷等,2008),土壤温湿度对凋落物分解的影响可能仅在大尺度上或者温湿度变化较大的情况下才会存在显著作用(王卫霞等,2016)。另一方面,初始凋落物质量作为影响分解最重要的功能特征,在林分尺度上被认为是影响凋落物分解的最主要因素(唐仕姗等,2014),本研究结果也支持此结论。除灌、伐除非马尾松和伐除优势马尾松处理明显改变了马尾松林凋落叶的初始基质质量,其中凋落物初始N含量分别降低1.74%、24.52%和27.65%,P含量分别降低29.27%、24.39%和14.63%,主要是因为抚育择伐后保留木的资源与养分竞争压力减小,生长条件相对变好,树叶生长周期可能加长,为老叶中的养分回流或淋失提供了更多时间,从而保留木凋落叶的N、P含量降低(Kunhamuetal., 2009)。多元逐步回归结果发现,凋落叶分解速率与初始N含量和N/P均存在显著正相关,特别是N/P成为抚育择伐下影响凋落叶分解速率的关键因素,这是因为N和P均是微生物参与分解过程的重要因素(林娜等,2010;Huangetal., 2007),凋落叶N/P决定了微生物的稳定性和适应方式,进而影响凋落物分解(周正虎等,2016)。有研究发现,当N/P<14时凋落物分解受N限制,N/P>16时受P限制(Güsewelletal., 2009),本研究结果与此观点相吻合。
本研究中,不同抚育择伐处理下马尾松林凋落叶分解速率存在较大差异,除灌、伐除非马尾松和伐除优势马尾松伐育择伐处理分别较未择伐提高了30.59%、2.30%和3.62%,以除灌处理影响最大。进一步分析其原因,可能与各处理影响林分结构和生态过程的差异有关。本研究采取的抚育择伐主要是针对林分不同树种和不同结构层次来开展的,在不同抚育择伐处理中,一方面除灌对林分结构影响属轻度干扰,凋落叶初始N含量显著高于2种处理,N/P最大;另一方面择伐后林分结构改变会提高林下植被生物多样性,增加微生物可利用养分和资源,改变微生物种类和数量,促使分解过程发生显著改变(Prescottetal., 2013)。野外调查发现,除灌处理可显著促进草本植物生长发育,草本易分解、周转快,可为微生物提供更多易利用的底物,增强了土壤微生物的活性(Gurleviketal., 2004)。伐除非马尾松和伐除优势马尾松处理虽然显著降低了凋落物初始N含量,但由于对上层林木的采伐会造成林冠郁闭度下降,光照增加,土壤温度升高,湿度降低,有利于凋落物分解(Chenetal., 2014)。伐除非马尾松处理导致原有林内马尾松树种比重增加、阔叶树种减少、凋落物组成更加单一,而伐除优势马尾松处理的采伐强度达到70%,林分组成结构已发生剧烈变化,短期内灌木层迅速发育,生物多样性显著提升,不同凋落物种类之间的营养交换能够促进土壤动物和微生物群落的活动(Zhaoetal., 2013),使得伐除优势马尾松处理的凋落叶分解速率高于伐除非马尾松处理。在相同地点开展的抚育择伐处理对马尾松林土壤微生物群落结构影响的研究(高尚坤等,2018)中发现,剔除非马尾松乔木会导致土壤微生物种系多度和谱系多样性显著降低,而除灌和伐除优势马尾松处理会使林下层植被大量更新发育,微生物生态位得到恢复,这可能是马尾松林凋落叶分解速率在伐除非马尾松处理中低于除灌和伐除优势马尾松处理的主要原因。因此,抚育择伐层次不同造成的林分结构和种类组成差异,会产生物种间生物学和生态学上的差异和互补性,造成凋落物功能特性及分解过程随森林结构变化而变化(Prescottetal., 2013)。
4.2 不同抚育择伐处理对马尾松林凋落叶养分释放的影响
一般而言,凋落物养分释放受凋落物初始质量和降雨等环境因素的影响(王娜等,2017b),养分元素迁移模式主要包括淋溶—富集—释放、富集—释放、直接释放等(李国雷等,2008;陈栎霖等,2015)。本研究中,不同抚育择伐处理下凋落物C元素具有相同的释放模式,表现为前期持续大幅度降低和后期轻微波动变化,主要是因为C是构成凋落物的主要元素,所含碳水化合物极易分解,前期释放快与降雨淋溶作用有较大关系,而后期受外界环境和微生物活动共同作用(Prescottetal., 2000)。理论上,凋落物分解过程中某种养分元素浓度相对较高能满足微生物的拓殖需求,则会发生释放,反之容易发生富集(王娜等,2017b)。
本研究中,各处理N释放模式均表现为初期短期释放后持续累积的趋势,与许多研究结果存在差异。如李国雷等(2008)研究间伐强度对油松(Pinustabulaeformis)凋落叶的影响时发现,间伐导致凋落物N呈释放—富集多次波动变化,而Chen等(2014)却发现油松凋落物分解N含量先升高后逐渐降低,可能与不同地区树种不同分解阶段所处的环境、凋落物自身特性以及干扰程度不同有关。但是,本研究结果与葛晓改等(2015)对三峡库区马尾松叶分解N的释放模式基本相似,原因可能是初期释放与降雨淋溶、物理破碎等非生物因素造成的凋落物质量损失有关,而后期凋落物中微生物生长和繁殖的需求导致N含量存在短暂累积(陈栎霖等,2015)。除灌、伐除非马尾松和伐除优势马尾松处理导致马尾松林凋落叶初始N含量分别降低1.74%、24.78%和27.65%,在整个分解过程中凋落物初始N含量始终影响其释放速率,N释放模式呈一致变化,表现为初始N含量越高后期凋落物基质N含量也越高。
不同抚育择伐处理和分解时间均未对P含量造成显著影响,但仍可看出分解初期凋落叶P含量呈一致性释放,随后在生长季内呈无规律变化且组内变异较大,可能与P容易受到雨水冲刷影响有关,还可能与抚育择伐对象和层次的差异造成林分冠层、树木分布等空间格局的异质性有关,因为P元素受控于物理因素、生物因素和化学因素,且以生物因素的影响较强(Laskowskietal., 1995),从而导致凋落物P的释放呈无规律变化,与微生物群落变化及对元素的固定有关(陈金玲等,2010)。
5 结论
不同抚育择伐处理均可使马尾松林凋落叶初始质量和林下环境发生明显改变,提高凋落叶分解速率,凋落叶年分解速率表现为除灌>伐除优势马尾松>伐除非马尾松>未择伐。影响马尾松林凋落叶分解速率的主要因素是凋落叶初始N含量和N/P,而不是林内空气温湿度和土壤温湿度等环境因素变化。抚育择伐能够明显促进C释放和N累积,但对P释放没有显著影响。由于择伐对象不同,造成林分组成和结构、凋落物基质数量和质量、林分水热条件发生改变,促使土壤微生物组成和结构变化,进一步对凋落物分解产生综合影响。未来开展马尾松林经营时,建议选择各林分层次的轻中度择伐,以保证合理的林分结构,更利于促进针叶人工林养分循环。
