‘冷白玉’枣果实发育过程中内源激素变化与胚败育的关系*
2020-06-02任海燕王永康赵爱玲薛晓芳弓桂花杜学梅李登科杜俊杰
任海燕 王永康 赵爱玲 薛晓芳 弓桂花 杜学梅 李登科 杜俊杰
(1.山西省农业科学院果树研究所 果树种质创制与利用山西省重点实验室 太谷 030815; 2.山西农业大学园艺学院 太谷 030801)
枣(Ziziphusjujuba)胚早期败育现象非常严重,阻滞了枣杂交育种。祁业凤等(2004)统计了《中国果树志·枣卷》中471个枣种仁发育情况,败育率高于50%的品种占58%。刘孟军等(2009)于2003—2005年连续3年调查国家枣资源圃300个枣品种的胚败育状况,发现花后1个月胚败育率高于90%的品种占33.8%。生产中枣农为获得较高的坐果率,长期运用花期开甲技术,并在花期多次喷施GA,取得了较高的经济效益(何业华等,1993;刘孟军等,2015)。但是,随着年份、气候条件的改变(金竹萍,2003),加之生产中GA的大剂量使用,出现了高比例的胚败育现象(梁春莉等,2010)。这种高比例的枣胚败育现象,严重制约了枣常规杂交育种的进程,枣果实发生胚败育的原因有待深入研究。
植物激素与胚败育有密切的关系,目前果树中已有葡萄(Vitisvinifera)(Wangetal., 2012;Reynoldsetal.,2016;Ebadietal.,2016)、荔枝(Litchichinensis)(陈伟等,2000)、芒果(Mangifera)(贺军虎等,2012;Shabanetal., 2009)等树种开展了相关研究。关于枣胚发育与激素的关系也有一些报道:胡芳名等(1998)在长枣(Z.jujuba‘Changzao’)上研究认为,幼果果肉中低含量的ABA和IAA可导致枣胚早期败育;祁业凤等(2004)比较了低度败育型冬枣(Z.jujuba‘Dongzao’)和高度败育型梨枣(Z.jujuba‘Lizao’)果实发育过程中激素水平变化情况,幼胚的正常发育需要高水平的ZT和IAA,且果肉中高含量的生长类物质不利于胚发育;王玖瑞等(2008)研究了枣雄性不育种质果实发育期间内源激素含量变化,认为生长抑制类激素和生长类激素的比值与胚胎发育有关。但上述研究主要采用了不同败育类型的枣品种作为试材。因高败育枣品种的败育胚难以取材,无法将败育胚与正常胚进行对比研究,这就给枣胚败育机理的研究带来了极大的困难。
选用具有正常胚和败育胚的同一枣品种为试材,可避免枣品种间胚发育时期不一致、败育胚取材难的问题。目前针对基因型一致的枣品种果实发育过程中内源激素含量变化的研究还未见报道。本试验以中等败育品种‘冷白玉’枣为试验材料, 从研究果实前期的发育及内源激素变化规律入手,初步探索枣胚败育机理。
1 材料与方法
1.1 试验材料
中等败育品种‘冷白玉’枣采自山西省农业科学院果树研究所国家枣资源圃,该品种盛花期为6月上旬,以黄蕾期(蜜盘刚出蜜)为自然授粉时期,定花定果,采集不同胚龄的幼果。
在前期研究的基础上,确认‘冷白玉’枣20、30、40、50天左右胚龄的幼胚依次正值原胚、小球形胚、球形胚、鱼雷形胚,且40天正值败育高峰期(李登科等, 2016; Renetal., 2019)。采集20、30、40、50天的幼果,每个时期采集50个,观测果发育状况,重复3次。每个时期采集幼果200个,重复3次,于冰块上剥开枣果,观察胚胎是否败育,并分为正常胚、败育胚、胚正常果实的果肉、胚败育果实的果肉共4组;收集材料要快速完成,用锡纸包裹,液氮速冻后-80 ℃保存,用于生长素(IAA)、赤霉素(GA4)、顺式脱落酸(cABA)和反式脱落酸(tABA)含量的测定。
判断胚是否败育的方法为:剖开果实观察,胚珠瘦小、部分组织褐化、变黑、软化、明显萎缩皱缩、干瘪坏死或胚完全消失为胚败育果实,反之则为胚正常果实。
1.2 试验方法
用电子天平称量果实质量,游标卡尺测量纵、横径,取平均值。剥开胚,观察胚珠发育状况。
称取果肉5.0 g、胚1.0 g进行激素含量的测定。每个时期重复3次。果实内源激素含量用氘代同位素内标定量计算法,激素组分用液相色谱-质谱(HPLC-ESI-MSn)定量分析检测(Liuetal., 2013),测试在北京林业大学生物工程重点实验室完成。
1.3 统计分析
各项指标均重复3次以上,取平均值。试验数据采用Excel 2003处理,采用SAS8.0进行Duncan’s多重比较(P<0.05)统计分析。
2 结果与分析
2.1 枣幼果及幼胚发育状况观察
‘冷白玉’枣花后10~50天,果实体积逐渐增大。在20~50天,随着果实逐渐增大,胚珠外表皮由绿白色变为淡黄色,种核逐渐木质化,胚乳由透明状变为乳白色固体状,胚起初由不可见逐渐微见绿色。在花后20天,胚珠外表皮白色微绿,胚乳透明晶亮(图1A);30天胚珠外表皮发育为白色(图1B);40天后胚珠外表皮呈黄白色,有韧性,胚乳呈乳白色(图1C);50天胚珠外表皮呈黄白色,核硬化,枣果皮颜色由深绿色逐渐变浅。解剖发现,正常胚体饱满而大,而败育胚不饱满,且胚珠表面局部或全部褐化(图1D)。花后40天,枣胚褐变发生败育,胚体变黑且失去活性。

图1 ‘冷白玉’枣果及胚胎发育情况
2.2 枣果实生长发育动态
由图2可知,枣果在花后20天,开始快速生长,起初单果质量增加较慢,花后30天快速增加;纵径生长比横径生长开始早,速度快,花后30天以后,纵径生长速度逐渐减慢,而横径生长速度增加。50天时纵、横径分别达到成熟果的60%~70%,质量为成熟果的58.83%。

图2 ‘冷白玉’枣果实的发育

图3 ‘冷白玉’枣果中GA4、IAA、cABA及tABA含量的变化
2.3 不同发育时期‘冷白玉’枣幼果中内源激素含量的动态变化
2.3.1 GA4含量的变化 由图3A可知,胚中GA4的含量明显低于果肉,并一直处于较低的水平。正常胚中GA4含量低于败育胚。花后30天,败育胚GA4含量开始升高;花后50天,败育胚含量达到最大,为0.302 ng·mg-1,且明显高于正常胚中GA4含量 0.011 ng·mg-1。
在果肉中,胚发育正常的果肉和败育果肉中的GA4含量变化规律基本相似,表现为先升高后降低的趋势(图3A)。然而,胚正常果肉中,GA4含量在花后40天达到最大值,为2.560 ng·mg-1;败育胚果肉中,GA4含量则在花后30天达到最大,随后降低,且低于幼胚果肉。
2.3.2 IAA含量的变化 胚中IAA的含量明显高于果肉。正常胚中IAA含量呈逐渐升高趋势,花后50天时达到最大,为15.895 ng·mg-1。而败育胚中的IAA含量则呈逐渐下降趋势(图3B)。
在果肉中,胚败育的果肉中IAA含量花后30天下降,之后逐渐上升并超过胚正常的果肉,而胚正常的果肉中IAA含量变化呈先上升后下降趋势,并在花后30天达到最大值,其后逐渐下降,40天后低于胚败育果肉中含量。胚正常果肉中IAA含量呈倒V字形,而胚败育果肉中呈正V字形(图3B)。
胚败育果的胚和果肉中IAA含量均呈逐渐下降趋势,而正常胚中其含量仍保持较高水平,且远远高于果肉(图3B)。这与祁业凤(2004)对梨枣的研究结果一致,梨枣果肉中IAA含量20天后呈逐渐下降趋势,说明幼胚中高水平的IAA有利于胚的正常发育。
2.3.3 ABA含量的变化 cABA在果肉和胚发育过程中变化较为一致,均表现为逐渐降低趋势。在胚正常枣果中,胚中cABA含量大于果肉中的含量;在胚败育枣果中,则表现为果肉中cABA含量大于胚中的含量(图3C)。
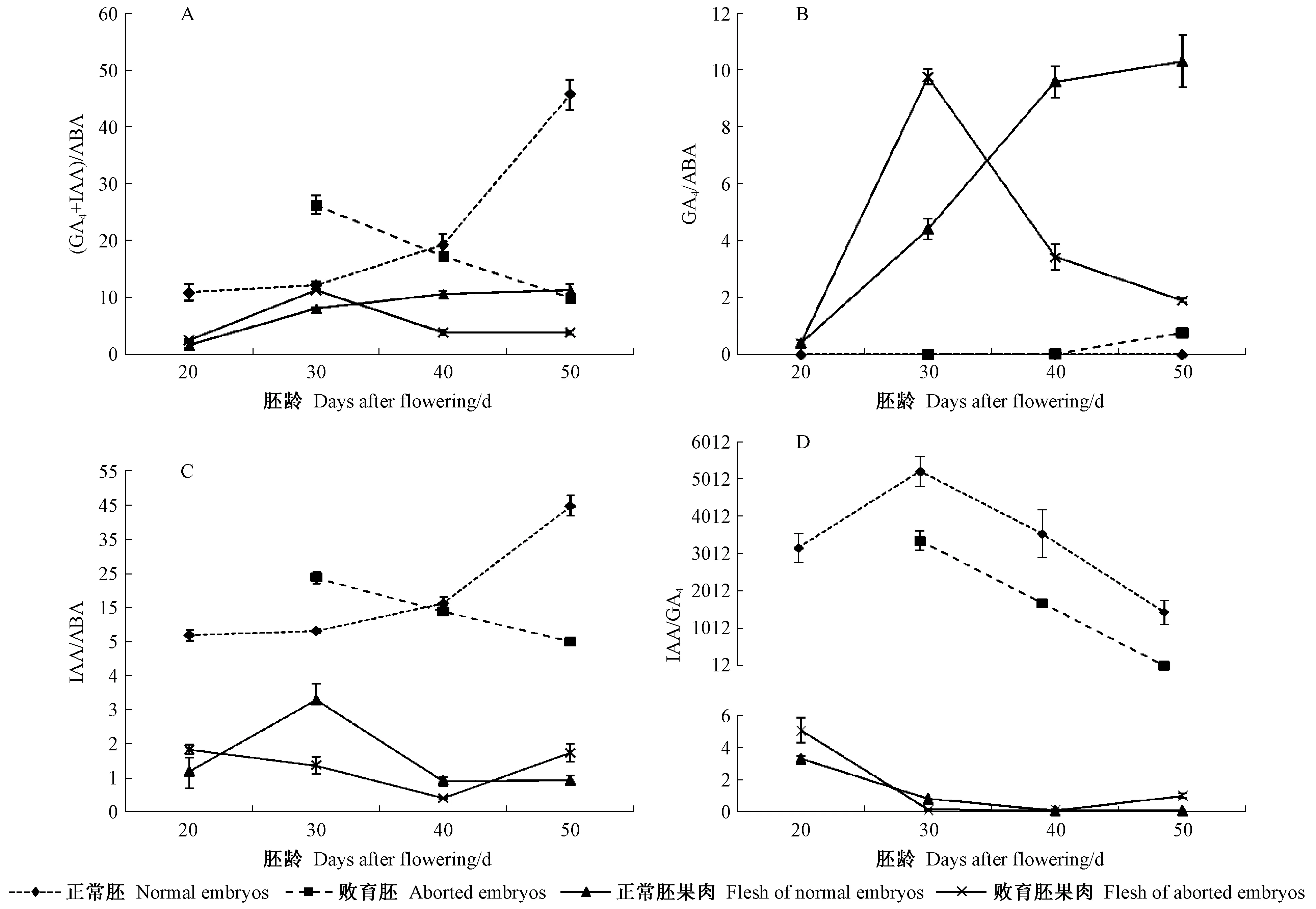
图4 ‘冷白玉’枣果中内源激素的平衡关系
胚中tABA的含量高于果肉。正常胚中tABA的含量在胚发育初期呈下降趋势,并在50天时达最低值;而在败育胚中tABA呈逐渐上升趋势,并在后期高于正常胚(图3D)。
胚正常发育的果实果肉中和胚中的tABA含量变化表现一致,呈先下降之后缓慢升高的趋势。而在败育胚的果肉中呈先下降30天后又上升,在40天出现一个小峰值,之后又下降,并高于胚正常的含量(图3D)。
2.3.4 内源激素平衡关系对果实发育的影响 从图4A得知,在胚中,花后20~50天,正常胚中(GA4+IAA)/ABA比值逐渐上升,而败育胚中迅速下降,二者差异显著。在果肉中,花后20~40天,胚正常果中(GA4+IAA)/ABA的比值呈逐渐上升趋势,在花后40天,达到峰值,之后逐渐下降;胚败育果中(GA4+IAA)/ABA的比值在30天达到最大,之后迅速下降,在40天后下降趋势逐渐缓慢。在30~40天,正常胚的果实中果肉的(GA4+IAA)/ABA比值逐渐上升,而胚败育的果肉中其比值迅速下降,而此时正值果实迅速膨大期,胚正常果肉中生长类的激素含量超过抑制类的激素并迅速上升,果实生长加快,而胚败育的果肉中抑制类激素超过了生长类激素,果实生长缓慢。
由图4B中GA4/ABA值看出,果肉中GA4/ABA大于胚中。在果肉中,胚正常果肉中GA4/ABA呈逐渐增高趋势;胚败育果肉中GA4/ABA在30天时达到峰值,之后降低。而在胚中GA4/ABA较低,正常胚中其变化呈逐渐降低,败育胚中其变化呈上升趋势。可见在胚中,较低的GA4/ABA利于胚的正常发育,而果肉中较高的GA4/ABA利于枣果的正常生长。
由图4C可知,胚中 IAA/ABA高于果肉。正常胚中,IAA/ABA不断增加,败育胚中,则呈下降趋势。胚败育果肉中,IAA/ABA呈先降低后升高趋势,在胚正常果肉中,在30天出现一个小高峰后逐渐降低,花后40天后,其比值保持平稳。可见,果肉中较高的生长促进类激素,增加了果肉争夺养分能力,使得胚获得养分减少,可能是导致枣胚败育的重要原因。
从图4D中IAA/GA4值的变化看出,胚中IAA/GA4远远大于果肉。在果实发育过程中,果肉中,其比值呈先降低后升高趋势,而在胚中表现为先升高后降低趋势,且胚正常型果肉大于胚败育型果肉,因此初步认为,胚中高比值的IAA/GA4利于幼胚的正常发育。
3 讨论
3.1 IAA含量变化与胚发育的关系
生长素在生物合成、极性运输和信号转导等多个层次控制着胚胎的发育过程(宋丽珍等,2013;翟开恩等,2015)。IAA是胚胎发育期间主要作用的生长素类物质,在胚胎发育过程中IAA含量的变化被认为是胚胎发生的早期信号,高水平的IAA可以维持较强的胚胎细胞活性,并通过细胞伸长来实现胚的膨大伸长,促进胚发育(Kongetal., 1997;梁艳等,2016)。本研究发现,正常胚IAA含量在20~30天即原胚和小球形胚发育时期缓慢增加,在40天(球形胚)后迅速增加,这可能与分生组织开始发育前多的IAA来维持更强的胚细胞活性有关;而败育胚中IAA含量变化整体呈下降趋势,起初缓慢下降,在败育高峰期花后40天球形期后急剧下降。Cheng等(2007)认为,在IAA调控信号途径中,低浓度IAA使yuc1yuc4yuc10yuc11四突变体在拟南芥(Arabidopsisthaliana)球形期胚发生阶段受到抑制,从而诱发胚发育缺陷。可见,IAA在枣胚发育过程中起重要作用。本研究结果中正常胚IAA含量逐渐增高趋势与在冬枣中的研究结果(祁业凤等,2004)一致,但不同于冬枣胚胎坐果后40天时IAA含量达到高峰之后维持较高水平的结果,这可能与不同品种间胚胎发育时期不同有关。
果实中的植物激素来源于种子,种子与果肉中的植物激素相互影响(Kondoetal.,2001)。在本研究中,果肉中IAA含量一直处于较低水平,远远低于胚中的含量,这与陈伟等(2000)、祁业凤等(2004)的研究结果相似,而不同于杜维等(2018)沙棘(Hippophaerhamnoides)果中IAA一直保持较高水平的研究结果。本研究中IAA在枣胚正常果肉中呈先上升后下降,这种变化趋势与葡萄果肉(Gouthuetal., 2015;高江曼等,2017)中及栗(Castaneamollissima)叶片中(Wangetal.,2012)IAA变化趋势相似。而在枣败育胚果肉中IAA含量变化呈正V形,相应的败育胚中则呈逐渐下降趋势。这很好地说明败育胚果肉中生长类激素的增加,使得果肉争夺能力增强,导致种子获得养分减少,发生败育。
3.2 GA4含量变化与胚发育的关系
GA被广泛用于葡萄、草莓(Fragaria×ananassa)、番茄(Lycopersiconesculentum)等果实的单性结实。目前发现赤霉素136种,其中GA3在促进细胞增长、形成无核果实等方面有重要的作用(Mesejoetal., 2016)。GA4较GA3的生物活性低,但对于胚轴生长和提高坐果率方面起着重要作用(Curry,2012;杜维等,2018)。对番茄的研究表明,GA4含量的增加受CcGa20oxl基因的调控,可导致单性结实(张映等,2018;Garcia-Hurtadoetal., 2012)。在褪黑素诱导‘红茄’梨(Pyruscommunis‘Starkrimson’)单性结实的研究中发现,外施褪黑素上调了子房内PbGA20ox并下调PbGA2ox基因的表达,使GA3和GA4含量增加,从而诱发单性结实的形成(Liuetal., 2018)。关于GA4影响枣胚发育的研究较少。从本研究‘冷白玉’枣果实发育过程中GA4的含量变化来看,果肉中GA4的含量明显高于胚,胚中GA4含量则一直保持较低水平。祁业凤等(2004)研究也得出枣果肉中GA3含量高于胚中含量的结果,但在变化趋势上有所不同,果肉含量变化表现为逐渐降低的趋势,不同于本研究中GA4含量呈先升高后降低的趋势。果肉中高含量的GA可能是多数枣败育型品种能继续膨大生长的一个原因,同样的结果也出现在Adam等(1999)的研究中。王玖瑞等(2008)研究表明,枣雄性不育种质胚中GA3含量高于果肉的含量,可能与不同品种间激素水平的差异有关。关于枣果的GA4、GA3含量变化在促进胚发育方面哪个发挥着更重要作用,其作用及其机理还需要进一步研究。
3.3 ABA含量变化与胚发育的关系
ABA在促进种子成熟期的贮藏蛋白合成方面起着重要的作用,此外还作为胚性状态保持剂有维持胚的正常生长发育等方面的作用(Quatrano, 1987)。研究表明,ABA 在红松(Pinuskoraiensis)胚胎发育过程中尤其是中后期的胚胎形态建成及早期阶段起着关键性的作用(梁艳等,2016)。在发育中后期胚中ABA的急剧上升是导致胚胎败育的一个重要因素(祁业凤等,2004;王玖瑞等,2008)。本研究中花后20~50天枣果发育阶段,胚正常果肉和胚败育果肉中的总ABA含量均呈下降趋势,其中胚败育果肉中总ABA含量高于胚正常果肉,而在胚中,正常胚总ABA含量逐渐下降,败育胚含量则逐渐增加。这也证实了在胚发育中后期总ABA含量的增加与胚败育有密切的关系。
天然存在的ABA是一个15碳的倍半萜羧酸,顺式脱落酸cABA和反式脱落酸tABA互为脱落酸的顺反异构体,其生理活性取决于C-2处的双键是顺式,即cABA是具有生理活性的,而tABA则相对比较稳定,在光中异构化后才有活性(Bhatlaetal., 2018)。依据ABA的作用方式划分为涉及核酸蛋白合成的“慢反应”和主要作用在质膜上的“快反应”。幼嫩的器官中所含的ABA能促进依赖于K+的H+/糖的卸载,而在成熟器官中ABA的主要功能可能是与启动特殊酶和蛋白质的合成及膜的泄漏有关(Qinetal., 1985)。在发育早期稻(Oryzasativa)胚中研究推测,cABA可能参与了“慢反应”,是总ABA库的钝化形态(覃章铮等,1990)。本研究中,果肉和胚中所含cABA含量随胚的发育逐渐降低,而tABA含量在中后期有所升高,由此推测在枣胚发育的中后期,具有活性的cABA可能部分转化为较为稳定的tABA,使得tABA在ABA影响胚发育的过程中起主导作用。胚败育和胚正常果肉中cABA含量变化趋势一致,均呈下降趋势,而胚中tABA含量高于果肉,在败育胚中,tABA呈持续上升的趋势,在40天后含量高于正常胚,且胚败育果肉中其含量也高于胚正常果肉。由此推测tABA可能与幼胚早期败育有关。
3.4 激素间动态平衡对幼胚发育的影响
胚胎发育不仅与单一内源激素含量有关,还与各类激素之间的平衡,尤其是促进生长类激素与抑制生长类激素之间的比例及平衡有关(许智宏等,2012)。ABA与GA之间存在拮抗作用,它们的相对含量及敏感性在胚发育过程中起重要作用(杨荣超等,2012;伍静辉等,2018)。IAA与GA之间相互协同,GA的合成需要IAA存在,IAA应答因子作为GA信号途径的激活剂,调控其表达(Mcateeetal., 2013)。本研究中,枣胚中(IAA+GA4)/ABA比值随胚的发育呈现逐渐增加的趋势,尤其在花后40~50天,即球形胚末期到鱼雷形胚快速增加,此时果实快速膨大,说明这个时期需要更高的生长促进激素;反之,(IAA+GA4)/ABA降低,不能满足幼胚生长所需,导致了败育的发生。此外,从(GA4+IAA)/ABA、IAA/ABA、GA4/ABA比值的变化趋势看,趋势较为一致,正常胚及其果肉均呈上升趋势,而败育胚及胚败育果肉中均呈下降趋势。这就意味着GA4、IAA、ABA影响胚胎发育可能是通过生长抑制激素与生长促进激素的平衡,这与在其他果树上研究的结果(陈伟等,2000;张新友等,2013)一致。
另外,胚的发育是由合子经过一系列细胞分裂而形成一个完整成熟胚的过程。作为刺激细胞分裂和生长作用的细胞分裂素,在胚的发育过程中起着重要的作用。陶建敏等(2006)认为,生长素和细胞分裂素的平衡影响胚的发育。并且较高的ZT、GA和IAA含量能促进子房的正常发育(Zangetal., 2011)。但也有研究认为,植物体内的GA、IAA、ABA激素变化是由乙烯信号诱导而成的,乙烯是胚败育的诱导因素(张凤路等,1999)。关于植物激素之间的关系、胚与果肉中各激素如何相互影响的机制较为复杂,仍待进一步探讨。
4 结论
‘冷白玉’枣胚中IAA的含量显著高于果肉,果肉中GA4的含量高于胚。胚中低水平的GA4、较高水平的IAA及后期低含量的tABA和cABA、高IAA/GA4比值有利于幼胚的正常发育。胚发育过程中IAA下降、tABA增加及(GA4+IAA)/ABA比值下降是导致胚败育的重要因素。枣幼胚发育前期,需要较高的生长类激素促进生长,而在发育后期则需要一定量的ABA,其中tABA与胚胎早期败育有关。
