不同光照强度下香溪河浮游植物演替过程研究
2020-06-01贡丹丹刘德富张佳磊严广寒
龚 川 , 贡丹丹,2, 刘德富, 张佳磊*, 严广寒,3
1.湖北工业大学土木建筑与环境学院, 河湖生态修复与浮游植物利用湖北省重点实验室, 湖北 武汉 430068
2.长江科学院流域水环境研究所, 流域水资源与生态环境科学湖北省重点实验室, 湖北 武汉 430010
3.中国环境科学研究院, 湖泊水污染治理与生态修复技术国家工程实验室, 国家环境保护饮用水水源地保护重点实验室, 北京 100012
基于2004—2015年水华事件跟踪监测结果,自2003年6月三峡水库一期试验性蓄水后,共有26条支流发生过水华,水华暴发主要集中在香溪河、大宁河、小江、梅溪河、汤溪河、磨刀溪和长滩河等7条距大坝相对较近的支流,其浮游植物群落由原来硅藻、甲藻为优势物种的河流型逐渐向蓝藻、绿藻为优势物种的湖泊型演替,严重影响了库区水生态系统和人民生产生活,引起国内外学者的广泛关注[1-5]. 从生态学角度来看,水华暴发的本质是藻种之间运动、竞争、死亡、被掠食等一系列生理生态过程的结果,即浮游植物种群由平衡状态演替为顶级状态[6]. 如果能采取某种措施将浮游植物群落稳定在平衡状态,从而维持其生物多样性,就能有效防控水华的暴发. 因此,确定影响三峡水库支流浮游植物演替的关键因素,筛选维持其群落平衡状态的主控因素阈值范围,为三峡水库生态调度方案的制定提供基础数据和理论依据,以及对河道型水库浮游植物演替机制的构建具有重要意义.
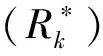
基于浅水湖泊或室内恒化器控制试验所得结论[18,20],光照强度是否是控制三峡水库支流回水区浮游植物演替的主控因素有待验证,同时维持其平衡状态的光照阈值有待筛选. 该研究基于前期三峡水库支流回水区的光照强度监测数据[21-22],设置室内控制不同光照强度,采用三峡水库一级支流香溪河回水区源水[23],验证光照强度是否是影响浮游植物演替的关键要素,并筛选维持其平衡状态的关键光强,以期为三峡水库生态调度方案的制订提供理论依据,为构建河道型水库浮游植物演替机制提供理论支撑.
1 材料与方法
1.1 藻种来源
依据三峡水库一级支流香溪河前期水华事件统计结果,收集香溪河水华易发区(31°08′02.70″N、110°46′42.20″E)的源水[1],利用Algacount S300浮游植物智能鉴定计数仪(杭州迅数科技有限公司)进行鉴定和计数[24-25]. 源水中主要浮游植物为绿藻、蓝藻和硅藻[26],依据该浮游植物生理生态特征,选取BG-11 为培养基,通过驯化、净化和饥饿培养得到试验用藻[27-28]. 各试验组藻种种类均来自同一源水,其起始浓度均为155.8×106cells/L,试验用藻共鉴定出5门17属,各门类种属及占比如表1所示.
1.2 试验装置
试验装置如图1所示,将外径为60 mm、内径为55 mm、高为90 mm的圆底玻璃管,置于长40 cm×宽25 cm×高60 cm的有机玻璃箱中,用水浴加热,并在水箱中放入微型水泵以扰动水体,使之受热均匀[29]. 插入高度为1 m、内径为5 mm的空心玻璃管,外接硅胶软管通过空气压缩机通入空气;进气前端设有孔径为0.2 μm的空气过滤器用以除尘除菌,另设置空气开关阀用以控制进气量大小[29]. 试验期间采取半连续培养方式,水温用可调式绝缘加热棒控制在(23±1)℃;培养液成分与BG-11培养基相同. 研究[30]表明,藻类生长适宜的ρ(DTN)、ρ(DTP)分别为2.214、0.038 4 mgL. 该试验选取ρ(DTN)、ρ(DTP)分别为5、0.5 mgL,保证其营养盐充足,且每天采用持续滴灌的方法补充高浓度培养液以维持试验装置中营养盐浓度稳定. 光源由白色日光灯组连续均匀地侧向供应,用不同灯管数调控光照强度,各处理组间用遮光布隔成单独小室,防止相邻光照之间的影响[29].

表1 试验用藻种属及占比
1.3 试验方法
依据前期香溪河水华易发区现场跟踪监测的水下光照强度数据,其全年光照强度范围在500~17 000 lx之间[21-22]. 因此,该实验根据前期监测数据及恒定光照下混合浮游植物竞争试验研究结果[29-30],设置0、1 500、3 000、4 500、6 000、8 000、10 000 和 13 000 lx八个梯度的恒定光照试验组,各梯度均设置两个平行样. 为保证完整准确地反映浮游植物的演替规律,按照浮游植物世代的3~4倍设置试验周期,试验时间为2015年10月29日—11月10日(共13 d),监测频率为1次d.

图1 试验装置示意Fig.1 Schematic diagram of the experimental device
1.4 测定指标及方法
试验开始后,于每天09:30取样. 光照强度和水温分别用TES-1339照度计(台湾泰仕电子工业股份有限公司)和0~100 ℃的玻棒煤油温度计(河北武强县光华仪表厂)测定[31];DTN、DTP和Chla浓度利用UV-1700紫外分光光度计(上海美析仪器有限公司)参考文献[25]进行室内分析;浮游植物的现场采样和室内分析参考文献[25]进行进行分析测定;藻密度和种类鉴定利用Algacount S300浮游植物智能鉴定计数仪(杭州迅数科技有限公司)参考文献[24]进行鉴定和计数. 藻种统计至属即可完整反映浮游植物群落结构的改变和演替过程[32]. 淡水水体中浮游植物世代周期一般为3~4 d[33],故浮游植物鉴定时间间隔为3 d次. 通过查阅文献[12,34]对所检出的藻种(属)进行功能组分类.
1.5 数据处理
1.5.1比增长率
绝大多数浮游植物种群增长符合指数增长模型,比增长率是指在某一时间间隔内浮游植物的生长速率,计算公式[35]:
μ=(lnN1-lnN0)/(t1-t0)
(1)
式中:μ为培养时间内的比增长率,d-1;N1、N0分别为t1、t0时藻细胞密度,cellsL.
1.5.2多样性指数
为避免使用单一多样性指数造成的多样性评价分析偏差,同时运用Margalef指数(S)、Shannon-Wiener指数(H)、Simpson指数(D)和Pielou指数(J)对浮游植物群落特征进行分析,计算公式[34-37]:
S=(M-1)/lnN
(2)
(3)
(4)
J=H/Hmax=H/lnM
(5)
式中:M为群落中物种总数;N为观察到的藻种总数;Ni为第i种藻的个数;Pi为第i种藻的频率,Pi=NiN;Hmax为Shannon-Wiener指数的最大值.
1.5.3演替速率
演替速率是群落结构改变潜势的客观尺度,Lewis[38]修正了群落演替速率计算方法,并在浮游植物群落研究中广泛应用,计算公式:
3.提出高度关注海洋、太空、网络空间安全。积极运筹和平时期军事力量运用,不断拓展和深化军事斗争准备,提高以打赢信息化条件下局部战争能力为核心的完成多样化军事任务能力。
(6)
式中:σs为演替率,d-1;S为物种总数;B2、B1分别为群落演替过程中2个时间状态(t2、t1)的物种丰度总和;b2i、b1i分别为群落演替过程中2个时间状态(t2、t1)群落中第i个物种的丰度.
2 结果与分析
2.1 不同光照强度下浮游植物生物量和群落多样性特征
2.1.1不同光照强度下浮游植物ρ(Chla)的变化特性
由图2可见:光照强度为 1 500 和 3 000 lx试验组的浮游植物生长周期分为延滞期(1~3 d)、对数增长期(3~10 d)和衰亡期(10~13 d)3个阶段,其他光照强度下源水中混合浮游植物的生长周期均符合浮游植物生长动力学曲线,均经过了延滞期(1~3 d)、对数增长期(3~10 d)和稳定增长期(10~13 d)3个阶段. 不同光照强度下ρ(Chla)存在显著性差异(p<0.05),恒化器试验装置中,其他环境条件适宜情况下,0~4 500 lx光照强度范围内,ρ(Chla)随着光照强度的增加而增加;4 500 lx光照强度下,ρ(Chla)达峰值(3 394.1 μgL);4 500~13 000 lx光照强度范围内,ρ(Chla)随光照强度的增加而减少,4 500 lx是最适宜香溪河源水中混合浮游植物生长的光照条件.

图2 不同光照强度下ρ(Chla)的变化特性Fig.2 Characteristics for concentration change of chlorophyll a under different light intensity
2.1.2不同光照强度下浮游植物多样性指数
研究期间,Margalef指数、Shannon-Wiener指数、Simpson指数和Pielou指数随光照强度的变化特征如图3所示. 由图3可见,Margalef指数在不同光照强度下差异不显著(p>0.05),Shannon-Wiener指数、Simpson指数和Pielou指数在不同光照强度下均差异显著(p<0.01). 光照强度为0~1 500 lx时,Shannon-Wiener指数、Simpson指数和Pielou指数均随光照强度的增加呈明显下降的趋势;光照强度为 1 500~4 500 lx时,Shannon-Wiener指数、Simpson指数和Pielou指数均随光照强度的增加呈显著上升的趋势;光照强度为 4 500~13 000 lx时,Shannon-Wiener指数、Simpson指数和Pielou指数均随光照强度的增加基本维持稳定. 因此,4 500 lx是在其他生境条件适宜时浮游植物多样性维持的最适光照强度.
2.2 不同光照强度下浮游植物的群落演替过程
2.2.1不同光照强度下浮游植物的演替速率
由图4可见:浮游植物的演替速率在不同光照强度下差异不显著(p>0.05). 无光照强度(0 lx)时浮游植物因无光照,无法满足生长需求而处于休眠状态,故演替速率始终较低,且变化不大;在演替的中期(4~10 d),光照强度为 6 000 lx的演替速率最高(0.263 d-1),达到顶级群落时各组的演替速率相差不大. 浮游植物群落在不同生长周期的演替速率存在显著差异(p<0.01),各光照强度处理组的演替速率在延滞期时相对缓慢,在对数增长期较快,在稳定增长期降至最低. 综上,光照因素对香溪河源水中混合藻的演替速率无显著影响,演替速率的时间变化规律符合浮游植物世代周期演替规律.
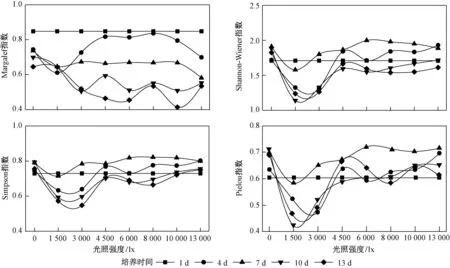
图3 不同光照强度下浮游植物的多样性分析Fig.3 Analysis of the phytoplankton diversity under different light intensity

图4 不同光照梯度下浮游植物的演替速率Fig.4 Succession rate of phytoplankton under different illumination levels
2.2.2不同种类浮游植物的最适光强

图5 香溪河源水中主要优势种的比增长率随光照强度的变化特性Fig.5 Specific growth rate variation of main dominant species in source water of Xiangxi River along with light intensity
2.2.3不同光照梯度下浮游植物群落生存策略演变模式
研究期间,浮游植物室内镜检共检出5门(绿藻门、蓝藻门、硅藻门、隐藻门和裸藻门)23属,最为丰富的是绿藻门(17属),其次是蓝藻门(3属),硅藻门、隐藻门和裸藻门各1属. 源水中起始群落结构由绿藻、蓝藻、硅藻、隐藻和裸藻组成,优势藻种为小球藻(46.34%),主要藻种为衣藻(17.46%),其余藻种含量均较低. 在试验末期(浮游植物稳定增长期衰亡期),不同光照强度下浮游植物群落结构存在显著差别: ①无光照强度时,该试验组里的藻种由初始的小球藻、衣藻演替为小球藻、衣藻和席藻,因小球藻为低光强耐受藻,因此可以在0 lx这一光限制条件下占优势. ②1 500 lx光照强度下,该试验组里的优势藻种在演替初期(1~4 d)和末期(10~13 d)均为小球藻、衣藻;而在 3 000 lx光照强度下,该试验组里的藻种由初始的小球藻、衣藻演替为衣藻,小球藻的最适光强为 3 000 lx,而衣藻最适光强为 8 000 lx,因此随着光照强度的增加,衣藻替代小球藻占优势. ③4 500 lx光照强度下,该试验组里的藻种由初始的小球藻、衣藻演替为衣藻和肾形藻,小球藻的最适光强为 3 000 lx,而衣藻和肾形藻最适光强分别为 8 000 和 6 000 lx,因此随着光照强度增加,衣藻和肾形藻替代小球藻占优势. ④6 000 lx光照强度下,该试验组里的藻种由初始的小球藻、衣藻演替为衣藻、肾形藻、微囊藻和栅藻,小球藻的最适光强为 3 000 lx,低于衣藻(8 000 lx)、肾形藻(6 000 lx)和微囊藻(6 000 lx)的最适光强,因此 6 000 lx是肾形藻和微囊藻的最适光照条件,衣藻、肾形藻和微囊藻替代小球藻占优势. ⑤8 000 和 10 000 lx光照强度下,该试验组里的藻种由初始的小球藻、衣藻演替为衣藻和栅藻,小球藻的最适光强为 3 000 lx,而衣藻和栅藻的最适光强分别为8 000和13 000 lx,因此光照强度更靠近衣藻和栅藻的最适光强,此时衣藻和栅藻替代小球藻占优势. ⑥13 000 lx光照强度下,该试验组里的藻种由初始的小球藻、衣藻演替为衣藻、栅藻和隐藻,衣藻、栅藻和隐藻的最适光强分别为 8 000、13 000 和 13 000 lx,远高于小球藻的最适光强,因此光照强度更靠近衣藻、栅藻和隐藻最适光强,此时衣藻和栅藻替代小球藻占优势. 综上,光照条件是决定香溪河源水中混合浮游植物植物群落演替方向的关键要素,其演替方向由此时光照条件与该群落中藻种最适光强的匹配程度决定.
根据不同藻种的生理生态特征,CSR理论将不同藻种生长特性及其环境适应机制划分为3种类型,分别为竞争者(Competitors,简称“C型”)、杂生者(Ruderals,简称“R型”)、环境胁迫耐受者(Stress-tolerators,简称“S型”). 由表2可见: ①不同光照强度下优势藻种演替趋势基本一致,初始时C型生存策略的小球藻的相对丰度为46.3%,为优势藻种,在演替中期时C型生存策略衣藻取代小球藻,并在末期保持绝对竞争优势,而小球藻则由优势藻种逐渐降为次优或更低. ②无光照强度时,初始期、延滞期、对数增长期和稳定增长期生存策略演变模式为C型(小球藻、衣藻)→C型(小球藻、衣藻)、S型(微囊藻)、R型(席藻)→C型(小球藻、衣藻)、S型(微囊藻)、R型(席藻)→C型(小球藻、衣藻)、R型(席藻),并未出现光限制效应使源水中浮游植物向S型生存策略浮游植物演变的趋势. ③1 500 和 3 000 lx光照强度下,初始期、延滞期、对数增长期和稳定增长期生存策略演变模式为C型(小球藻、衣藻)→C型(小球藻、衣藻)→C型(小球藻、衣藻)、S型(色球藻)、R型(肾形藻)→C型(小球藻、衣藻),表明 1 500~3 000 lx光照强度范围比较适宜C型生存策略的浮游植物生存; 同时,当光照强度由 1 500 lx升至 3 000 lx时,在对数增长期生存策略为S型和R型的浮游植物增多,表明光强的升高有利于S型和R型浮游植物的生存. ④4 500 lx光照强度下,初始期、延滞期、对数增长期和稳定增长期生存策略演变模式为C型(小球藻、衣藻)→C型(小球藻、衣藻)、S型(微囊藻)→C型(小球藻、衣藻)、R型(肾形藻)、S型(色球藻)→C型(衣藻)、R型(肾形藻). ⑤6 000、8 000 和 10 000 xl光照强度下,初始期、延滞期、对数增长期和稳定增长期生存策略演变模式为C型(小球藻、衣藻)→C型(小球藻、衣藻)、S型(微囊藻)→C型(小球藻、衣藻、隐藻)、S型(微囊藻、色球藻)、R型(肾形藻)→C型(衣藻、栅藻)、R型(肾形藻)、S型(微囊藻). ⑥13 000 lx光照强度下,初始期、延滞期、对数增长期和稳定增长期生存策略演变模式为C型(小球藻、衣藻)→C型(小球藻、衣藻)、S型(微囊藻、色球藻)→C型(小球藻、衣藻、隐藻、栅藻)、S型(微囊藻、色球藻)→C型(衣藻、栅藻、隐藻),未出现光限制效应使源水中浮游植物向S型生存策略浮游植物演变的趋势. 总体上,CSR理论的核心是认为浮游植物演替是由能量、物质协同作用的结果,因此并不能准确解释不同梯度恒定光照控制条件下浮游植物的演替规律.

表2 试验期间浮游植物群落演替序列
3 讨论
光照通过影响浮游植物光合活性、放氧速率、酶活性等影响其光合效率,进而影响浮游植物的生长过程[40-41]. 研究[42-43]发现,光照是控制春季第一场硅藻水华的关键因子,且对藻类的连续生长和水华暴发时间起到了驱动作用. 浮游植物均存在光饱和强度,光照不足或光照过饱和都会对浮游植物光合作用产生不利影响[44-46]. 该研究拟通过探究香溪河源水中混合藻种生长的最适光强,使其在藻种快速增长时能维持其生物多样性,避免向单一水华藻种的顶级状态发展,从而达到抑制水华的目的.
目前,对单一藻种适宜光强范围的探索是研究光照强度影响藻类生长特性和演替规律的重点. 前期单一藻种室内控制试验结果[47-49]表明,小球藻、栅藻、微囊藻、小环藻、隐藻、色球藻和衣藻的最适光强分别为 4 500、5 300、2 000~5 000、3 000~6 000、7 500、2 000~5 000 和 5 000 lx. 根据CSR理论,小球藻、衣藻、栅藻、隐藻均属于C型藻种,其生长所需光照条件相对较低,可迅速吸收营养盐快速繁殖,获得竞争优势;微囊藻、色球藻属于S型藻种,其生长增殖速率相对较慢,对资源的发掘、获取能力强,通常能在能量或物质供给相对短缺的条件下,有效调整需求应对有限的资源,对环境条件的耐受性较强;小环藻属于R型藻种,其可调和、适应环境,但也受到一定限制,能长期在很低的光照条件下生长,也耐受于高光照条件,能够抓住机会将资源转化为自己生长所需能量.
前期香溪河围格试验及现场监测结果显示:光照强度的变化是导致香溪河浮游植物初级生产力变化的主要原因[21];光照强度改变导致水体中光混比发生变化,当混合层与真光层比值为0.5时对蓝藻增殖影响最大[23];小球藻和栅藻的最适光照强度范围为625~ 1 125 lx[22]. 该试验设定ρ(DTN)、ρ(DTP)分别为5、0.5 mgL,远高于藻类生长所需营养盐阈值[30],保证试验期间营养盐充足;为藻类生长提供适宜的物质条件,水温控制在(23±1)℃,为大多数藻类生长适宜温度;光照强度设置0、1 500、3 000、4 500、6 000、8 000、10 000 和 13 000 lx八个梯度,且在试验过程中维持各组光强稳定,最大误差为±200 lx;此外,试验所用藻种及各藻种初始比例均是香溪河源水在自然条件下驯化后所得,仅将其非稳态条件改为恒定光照条件. 结果表明:不同梯度光照强度条件下优势藻种演替趋势基本一致,初始时C型生存策略的小球藻为优势藻种,占比为46.3%;演替中期时,C型生存策略的衣藻取代小球藻,并在末期保持绝对竞争优势,而小球藻则由优势藻种逐渐降为次优或更低的层次. 在高光照(13 000 lx)和低光照(0 lx)试验组中,藻类均未出现光限制效应使源水中浮游植物向S型浮游植物演变的趋势. 结果表明,在浮游植物其他生境条件适宜,仅光照条件不同的条件下,小球藻、衣藻、栅藻、肾形藻、微囊藻、色球藻、隐藻和小环藻最适光强分别为 3 000、8 000、13 000、6 000、6 000、13 000、13 000 和 6 000 lx,其中,小球藻属于低光强耐受藻,栅藻、色球藻和隐藻则属于高光强耐受藻.
对比发现,实验室恒化器中单一藻种和现场围格试验中藻种的最适光强与该研究结果有较大区别,推测其原因是现场围格试验期间,受降雨等的影响,整体光照强度较低,且放入围格中的试验藻种初始比例相同,直至试验结束,围格中的藻密度也一直处于相对较低的水平,小球藻、栅藻等在现场围格试验条件下占优也可能是因为其本身属于竞争型藻种,生长所需光照条件相对较低所致;而实验室恒化器试验也是基于单一藻种或藻种初始比例相同的情况下得出的藻种最适光强. 对比发现,各藻种本身的生存策略并不能准确解释该试验结果,在混合藻种的香溪河源水光限制试验中,各藻种初始比例不同,所以各藻种的生长演替除了受生境条件的影响外,还受内部竞争的影响,且该研究结果表明其演替方向取决于藻种本身的生存策略及其在单一藻种试验中所得到的最适光强的匹配程度,所以单一藻种与香溪河源水中混合藻种培养试验中所得出的藻种的最适光强有较大的区别;同时,在其他浮游植物环境条件适宜的情况下,光照强度为 4 500 lx时,香溪河源水中混合藻种的生物量和多样性均达到最高.
该研究表明,光照条件是决定香溪河源水中混合浮游植物群落演替方向的关键要素,其演替方向由此时光照条件与该群落中藻种关键光强的匹配程度决定. 前期三峡水库香溪河水华事件研究[21-23,26]表明,香溪河冬末春初多暴发硅藻水华,春末夏初多暴发绿藻水华,夏季多暴发蓝藻水华. 区别于太湖、东湖等浅水湖泊的蓝藻水华,三峡水库支流回水区暴发多藻种优势水华的原因是由于三峡水库支流回水区的光照强度除了受到太阳辐射强度的影响外,还受到三峡水库调度所带来泥沙含量变化的影响,因此在冬末春初光照强度较低时,最适光强较低的硅藻演替占优形成硅藻水华;而春末夏初光强逐渐回升,最适光强相对较高的绿藻演替占优形成绿藻水华;夏季光强达最强,由于蓝藻的最适光强相对其他藻种较高,所以蓝藻在此时演替占优形成蓝藻水华.
研究[13,17,21,47]表明,改变水域的光照条件是控制水库水华的重要手段. 日本琵琶湖通过遮光控制技术有效抑制了琵琶湖水华的暴发,显著改善了琵琶湖的水质状况[50]. 孙扬才等[51]遮光技术示范工程表明,该技术有效降低了水下光照强度,大幅削减了藻类生物量,显著抑制了藻类光合产氧速率,促使藻类衰亡. 三峡水库支流回水区由于水域面积较大,同时水华暴发带有一定的不确定性,因此遮光控藻很难实现[52]. 前期三峡水库调度机制研究[4,14,28]表明:基于水华成因的研究结果和中度扰动理论,可以通过水库调度改变水动力条件来调整其水体层化结构,影响水体中的垂向掺混模式及真光层、混合层深度,以及影响藻类的光合作用效率,从而抑制浮游植物的生长;同时,通过水库调度使水环境处于适当范围的变化,从而保证物种的多样性. 因此,可以基于该研究筛选出的维持浮游植物群落平衡状态的光照强度阈值,通过生态调度将水体中的真光层与混合层的比例及扰动程度控制在适宜范围,再加上前期所构建的水动力—光场—浮游植物响应关系,制定相应的调度方案. 前期整流幕在三峡水库的数值模拟研究[53-54]显示,整流幕能较好的防控水华,水库上游来流与整流幕接触而发生冲击,形成较强扰动的水体环境,使水体透明度降低,真光层深度减小,且在垂向掺混作用下,使藻类光照条件发生改变. 因此,可以综合该研究所筛选出的维持浮游植物群落平衡状态的光照强度阈值与整流幕数值模拟结果,制定相应的防控方案.
4 结论
a) 在浮游植物其他生境条件适宜,仅设置不同恒定光照条件时,小球藻、衣藻、栅藻、肾形藻、微囊藻、色球藻、隐藻和小环藻最适光强分别为 3 000、8 000、13 000、6 000、6 000、13 000、13 000 和 6 000 lx,小球藻属于低光强耐受藻,栅藻、色球藻和隐藻均属于高光强耐受藻.
b) 在其他浮游植物环境条件适宜的情况下,4 500 lx是最适宜香溪河源水中混合浮游植物生长的光照条件,同时也是维持其多样性的适宜光照条件.
c) CSR理论的核心是认为浮游植物演替是由能量、物质协同作用的结果,并不能准确解释不同梯度恒定光照控制条件下浮游植物的演替规律. 光照条件是影响香溪河源水中混合浮游植物群落演替方向的关键要素,其演替方向由群落发生演替时光照条件与该群落中藻种最适光强的匹配程度决定.
