柔海胆科(Echinothuriidae)3个属叉棘超微结构的研究*
2020-05-25郑婉瑞安建梅
郑婉瑞 肖 宁 安建梅
柔海胆科(Echinothuriidae)3个属叉棘超微结构的研究*
郑婉瑞1肖 宁2①安建梅1①
(1. 山西师范大学生命科学学院 临汾 041000; 2. 中国科学院海洋研究所 海洋生物分类与系统演化实验室 青岛 266071)
海胆的叉棘是海胆御敌、清除体表异物的重要器官, 其形状在海胆的分类上具有重要意义。本文通过扫描电子显微镜对西太平洋海山区柔海胆科软海胆属兜海胆属和革海胆属的叉棘形态与结构特征进行研究, 并提供了柔海胆科属的检索表。结果表明: 柔海胆有三种常见的叉棘形式, 三叉叉棘、三叶叉棘和指状叉棘。三叉叉棘和三叶叉棘在3个属中均存在, 指状叉棘只在软海胆属中存在, 三叶叉棘和指状叉棘的超微结构变化比较细微, 三叉叉棘的超微结构特征具有明显的种属间差异。建议将叉棘的种类和结构作为柔海胆科的分类依据。
深海; 柔海胆; 叉棘; 形态学; 扫描电镜
叉棘是棘皮动物海胆纲和海星纲特有的一种微小的棘, 尤以海胆纲最为发达, 是海胆御敌、清除体表异物的重要器官, 在海胆壳的棘间、围口部和围肛部最为丰富(Tamori, 2018)。叉棘的头部通常有3个瓣, 但偶尔有2个、4个或5个, 连接在长度不同的柄上, 头部和柄之间相接可伸缩的颈部(连接部)(Ramalingam, 1975)。叉棘的头部和柄部为钙质组成, 表面覆盖很厚的肌肉组织(吴古远等, 2017), 连接部也由肌肉组织构成, 头部肌肉具有开放或关闭瓣的作用。根据其头部结构的不同可将叉棘分为4种基本形式: 三叉叉棘、三叶叉棘、蛇首叉棘和球形叉棘(廖玉麟, 1982), 每种叉棘都有不同的大小和形状的变化, 其形状是海胆的分类上的重要特征(由香莉, 2003)。Brosseau等(2003)利用扫描电镜对柄头帕海胆近似种的叉棘结构进行了详细的描述; 吴古远等(2017)通过扫描电镜研究了刻肋海胆科的四种叉棘, 证明叉棘的超微结构在刻肋海胆科中具有明显的种间差异; Tamori等(2018)研究了喇叭毒棘海胆球形叉棘的结构及作用机制。但目前专门针对深海柔海胆叉棘的研究还是较少。
柔海胆科(Echinothuriidae)隶属于棘皮动物门(Echinodermata)海胆纲(Echinoidea)柔海胆目(Echinothurioida)(Kroh, 2010), 包含7属52种。当中除了囊海胆属的种类主要分布在沿岸海域, 其余完全为半深海或深海种。柔海胆类具有柔软的壳体与薄的重叠的骨板很容易与其它海胆区分, 与其它海胆相比, 它们壳体又宽又低, 被捕获时, 壳体通常会完全塌陷变扁平(Anderson, 2013, 2016)。由于水压等原因, 采集的柔海胆标本经常破损, 因此仅凭柔海胆的外部形态及步带、间步带、顶系和管足孔等特征难以较准确的进行分类工作。叉棘在柔海胆中非常丰富, 有着不同的数量、大小和形状(Mortensen, 1935), 且特征明显, 可以为分类学提供更有效的数据。柔海胆叉棘的有四种: 三叉叉棘、三叶叉棘、指状叉棘和蛇首叉棘, 真正的球形叉棘在任何柔海胆中均未发现(Mortensen, 1935), 蛇首叉棘非常罕见, 指状叉棘为软海胆属所特有的(Mortensen, 1903; Mooi, 2004)。目前已有的柔海胆叉棘描述大多为文字描述以及手绘的方式, 难以清楚的展现柔海胆叉棘的细微结构。本研究采用扫描电子显微镜的方式获得柔海胆科软海胆属兜海胆属和革海胆属海胆叉棘的超微结构, 丰富了柔海胆叉棘的研究。
1 材料与方法
1.1 实验材料
实验样品分别采自西太平洋南海蛟龙海山区、雅浦海山区、马里亚纳海山区、卡罗琳海山区, 采集方式为潜水机器人(remotely operated vehicle, ROV), 采集时间从2014年至2019年。海胆采集后用海水冲净表面异物, 用75%的乙醇固定备用。实验样品具体采集信息(见表1)。
1.2 实验方法
在解剖镜下找到完整的海胆叉棘, 用镊子小心取下, 浸泡于95%的乙醇中备用。将取到的叉棘置于培养皿中, 加入10%的次氯酸钠溶液, 没过叉棘, 以分离和移除瓣膜中的肌肉组织。根据不同浸泡效果可适当调节浸泡时间, 叉棘不同浸泡时间不同, 浸泡过程中不时在显微镜下观察, 以免浸泡过度。待叉棘表面的肌肉组织溶解完全后, 蒸馏水清洗2遍酒精清洗1遍, 清洗过程中注意不要损坏叉棘。将清洗后的叉棘置于碳导电双面胶带上(15mm), 干燥, 喷金。在扫描电子显微镜(Hitachi-3400N)下找到目标叉棘, 观察单个瓣以及整体的形态与结构特征, 并拍照记录。
表1 实验海胆的采集信息
Tab.1 The information of the sample collection for the study
2 结果
不同海胆因栖息环境与生活习性的不同, 海胆叉棘的种类和结构存在明显的分类特征。而海胆叉棘的连接部与柄部基本相同, 因此本研究以海胆叉棘的头部为主。
2.1 三叉叉棘
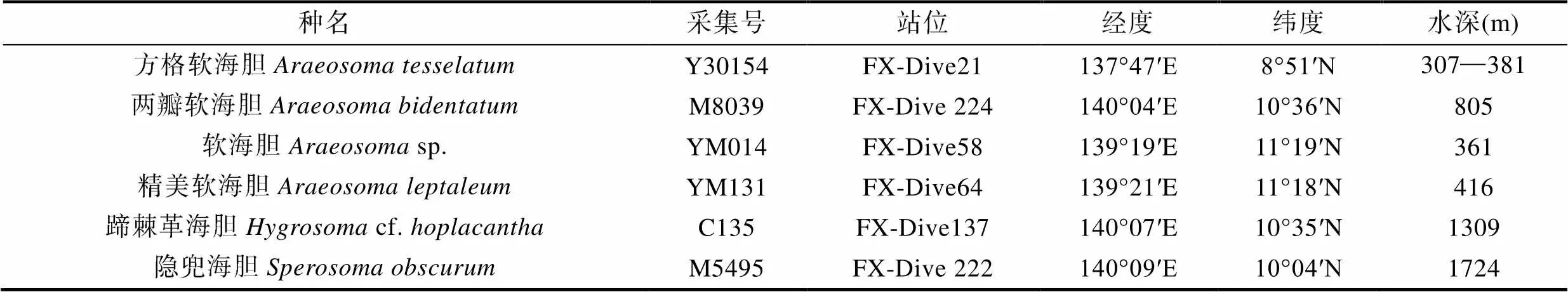
三叉叉棘在柔海胆中较丰富, 并且有许多不同的大小和形状。其基本特征为瓣具有复杂的网状结构, 基部膨大呈三角形, 闭合时整体呈锥形。
隐兜海胆具有一个类型的三叉叉棘, 为简单的叶形, 其基部向上渐窄, 近顶端微宽; 瓣狭长, 不内卷; 闭合时仅末端与1/2的瓣相接, 半封闭(图1a, b), 有的边缘具刺状凸起(图3a), 有的边缘无刺状凸起(图3b)。
两瓣软海胆具有三个类型的三叉叉棘。较大的三叉叉棘瓣具龙骨, 有强烈的内卷; 顶端宽度约为基部的2/3, 边缘深波状; 颈部细长于2/3处向外凸起, 长度约为整体的1/3; 闭合时首尾在铰合部连接, 中间不相连, 内部包被空间(图1c, 图3c)。中等的三叉叉棘瓣狭长, 不内卷边缘无锯齿; 基部向上渐窄, 闭合时瓣全部相接, 仅近基部微开口(图1d, 图3d)。较小的三叉叉棘仅由两瓣组成, 瓣短而粗, 边缘无锯齿, 不内卷; 闭合时瓣全部相接, 仅近基部微开口(图1e)。

图1 三叉叉棘(2-1)
注: a—b: 隐兜海胆; c—e: 两瓣软海胆; f—g: 方格软海胆
方格海胆具有三个类型的三叉叉棘。大的三叉叉棘瓣有强烈的内卷, 中间窄; 顶端边缘深波状, 宽度约为基部一半, 颈长约为整体的1/2; 闭合时, 首尾在铰合部连接, 中间不相连, 内部包被空间(图1f)。中等的三叉叉棘顶端稍宽越为基部的一半, 边缘无锯齿; 颈部短, 约为整体的1/4; 瓣具龙骨, 内卷; 闭合时首尾在铰合部连接, 在中间约有1/3的开口(图1g, 图3e)。小的三叉叉棘无头颈之分, 瓣狭长, 基部向上渐窄, 约1/3处停止; 边缘无锯齿, 不内卷(图3f)。
软海胆sp.具有两个类型的三叉叉棘。大的三叉叉棘瓣强烈内卷; 顶端宽约为基部的1/3, 边缘深波状; 颈狭长, 长度约为整体的1/2; 闭合时首尾在铰合部连接, 中间不相连, 内部包被空间(图2a, 图4a)。小的三叉叉棘瓣宽, 无头颈之分, 具龙骨; 边缘无锯齿, 不内卷; 闭合时瓣全部相接, 仅近基部微开口(图2b, 图4b)。

图2 三叉叉棘(2-2)
注: a—b: 软海胆sp.; c: 精美软海胆; d—e: 蹄棘革海胆cf.
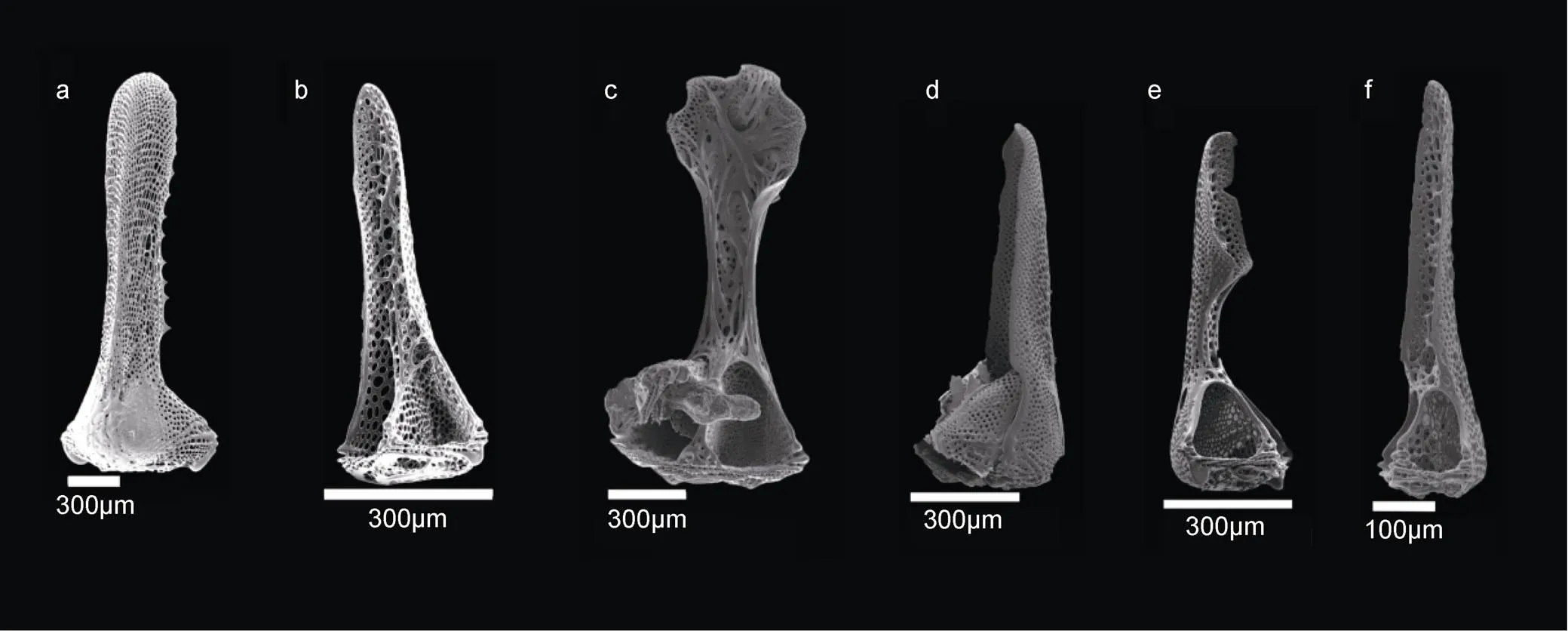
图3 三叉叉棘的瓣(2-1)
注: a—b: 隐兜海胆; c—d: 两瓣软海胆; e—f: 方格软海胆
精美软海胆具有两个类型的三叉叉棘。第一种三叉叉棘具不同大小, 瓣宽, 边缘深波状具龙骨, 不内卷; 基部向上渐窄, 闭合时瓣全部相接, 完全封闭(图2c, 图4c)。第二种三叉叉棘瓣狭长, 基部向上渐窄, 边缘无锯齿, 不内卷(图4d)。
蹄棘革海胆cf.具有两个类型的三叉叉棘。较大的三叉叉棘瓣狭长, 内卷; 顶端宽度约为基部的1/4倍, 边缘无锯齿, 内凹; 颈部狭长约为顶端长的3倍; 闭合时首尾在铰合部连接, 中间不相连, 内部包被空间(图2d, 图4e)。较小的三叉叉棘内卷, 顶端勺子状, 宽度约为基部的1/3倍; 颈部狭长, 约为顶端长的1.5倍; 闭合时首尾在铰合部连接, 中间不相连, 内部包被空间(图2e, 图4f)。

图4 三叉叉棘的瓣(2-2)
Fig4 Tridentate pedicellariae valves (Part 2 of 2)
注: a—b: 软海胆.; c—d: 精美软海胆; e—f: 蹄棘革海胆cf.
2.2 三叶叉棘
三叶叉棘是最小的叉棘, 在柔海胆科中变化不大, 具短而宽的瓣, 每瓣呈叶片状, 顶端弯曲略内凹, 边缘密布规则的栅栏状锯齿, 排列紧密; 瓣充满了极其密集和复杂的网状结构(Agassiz, 1879); 基部窄顶端宽; 闭合时在顶端不相接, 内部包被空间(图5a)。隐兜海胆的三叶叉棘内部有竖向支架呈“Y”字形, “Y”字形横向支架连向瓣两侧边缘, 边缘内凹(图5b)。两瓣软海胆三叶叉棘内部有竖向支架呈“工”字形, 上下同宽, “工”字形横向支架连向瓣两侧边缘, 边缘内卷(5c)。蹄棘革海胆cf.的三叶叉棘内部有竖向支架呈“Y”字形, “Y”字形横向支架连向瓣两侧边缘, 边缘内卷(图5g)。其余物种内部无“工”字形或“Y”字形支架, 瓣两侧连接部较多, 边缘均内卷且有网状结构(图5d, e, f)。
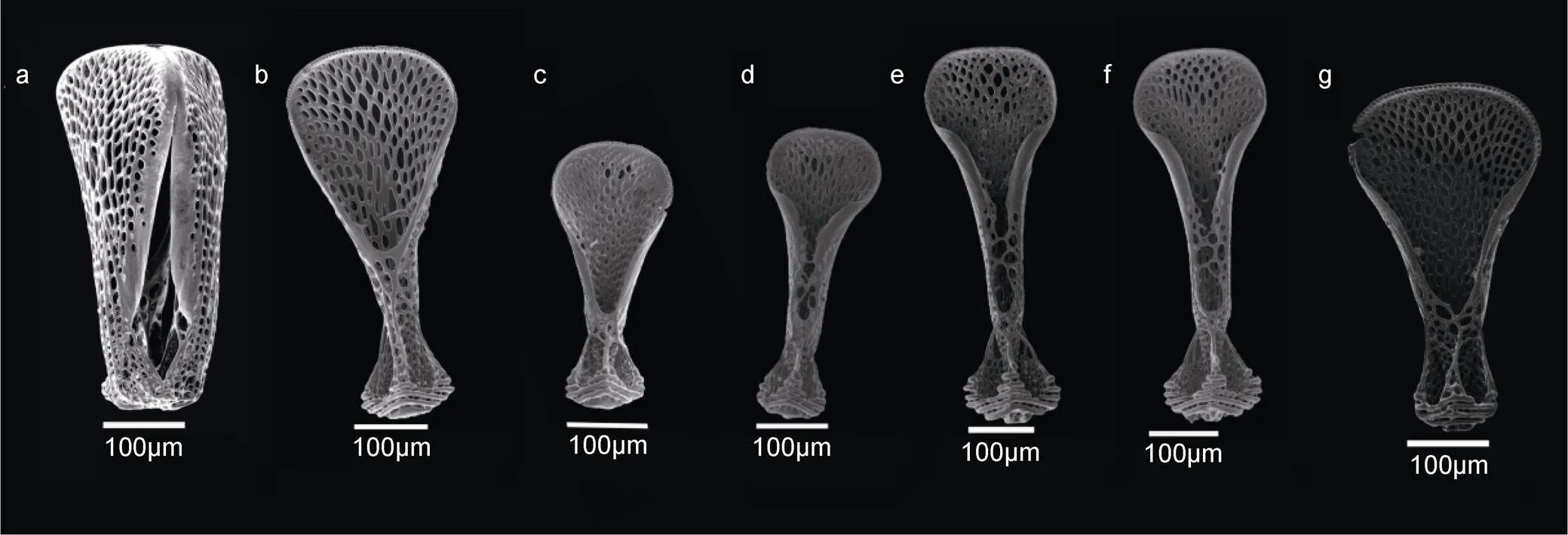
图5 三叶叉棘
注: a—b: 隐兜海胆; c: 两瓣软海胆; d: 方格软海胆; e: 软海胆.; f: 精美软海胆; g: 蹄棘革海胆cf.
2.3 指状叉棘
指状叉棘为软海胆属所特有的(Mortensen, 1903; Mooi, 2004), 瓣发达, 形稍精致, 因具毒囊可能是球形叉棘变形。本研究的样品在采集过程和保存过程中或多或少有些损伤, 所以本实验只在两瓣软海胆中发现了指状叉棘。其具有6个瓣, 瓣精美, 形狭长, 顶端呈匙状, 其边缘类似牙齿的形状(图6)。

图6 两瓣软海胆的指状叉棘
3 讨论
3.1 叉棘在柔海胆科属间的差异比较
见表2。本实验共取得了三种叉棘形式: 三叉叉棘、三叶叉棘和指状叉棘。三叉叉棘和三叶叉棘在3个属所有标本中均取得, 兜海胆属三叉叉棘形状较原始, 为简单的叶形, 边缘无内卷, 瓣上或多或少布满了复杂的网状结构, 有的凸起成刺状, 闭合时瓣全部相接, 仅近基部微开口(图1a, b)。软海胆属三叉叉棘除简单的叶形之外(图1d), 还有的发展成复杂的形式(图1c), 具内卷, 深波状的瓣; 从一个仅基部有点开放和扩大的封闭锥形体结构发展成内部包被空间的锥形体结构。革海胆属较小的三叉叉棘顶端具有典型的勺子状, 高度复杂, 边缘内卷, 闭合时内部包被空间。由此可见, 三叉叉棘的超微结构特征具有明显的属间差异。三叶叉棘典型叶片状(图5), 在柔海胆科中变化较小, 顶端弯曲略内凹, 边缘密布规则的栅栏状锯齿, 排列紧密; 瓣充满了极其密集和复杂的网状结构。指状叉棘是软海胆属的典型叉棘(图6), 比较罕见, 本研究只在两瓣软海胆中取得, 一般具有4或5个瓣(少数具6个瓣), 瓣精美, 形狭长, 顶端呈匙状, 其边缘类似牙齿的形状, 变化细微。
表2 海胆叉棘差异

Tab.2 The differences of the pedicellariae
3.2 三叉叉棘在软海胆属种间的差异比较
从我们的研究中, 可以看出三叉叉棘的种类与结构在软海胆属种间差异明显, 特别是复杂形式的三叉叉棘边缘、顶端与基部宽的比和颈长。如: 两瓣软海胆顶端宽度约为基部的2/3, 边缘深波状, 颈长约为整体的1/3(图1c, 图3c)。方格软海胆较大的三叉叉棘顶端宽度约为基部1/2, 边缘深波状, 颈长约为整体的1/2(图1f)。较小的三叉叉棘顶端宽越为基部的1/2, 边缘无锯齿, 颈长约为整体的1/4(图1g, 图3e)。软海胆sp.顶端宽约为基部的1/3, 边缘深波状; 颈长约为整体的1/2(图2a, 图4a)。精美软海胆仅具简单叶形, 无复杂的三叉叉棘等。
4 结论
本文通过扫描电子显微镜对西太平洋海山区柔海胆科软海胆属、兜海胆属和革海胆属的叉棘形态与结构特征进行了研究。描述了3种叉棘的形态特征, 提供了叉棘整体与单个瓣结构的电镜图, 并比较了叉棘在柔海胆科属间和种间的差异。结果表明: 叉棘的种类和结构可以作为柔海胆科的重要分类依据, 特别是三叉叉棘的超微结构。为今后柔海胆科的分类工作提供了有效数据。由于我们目前只采集到了柔海胆科三个属的标本, 该科另外四个属的叉棘结构仍需进行进一步的比较研究。
致谢 感谢台湾自然科学博物馆生物学组李坤瑄老师在实验过程中的指导以及文章撰写中的建议, 感谢中国科学院实验海洋生物学重点实验室刘伟老师在扫描电镜拍摄中给予的帮助, 谨致谢忱。
附录 柔海胆科属的检索表(改自Smith, 2013)
1. 步带管足孔在口面与反口面均为三列··············································································································· 4
- 步带管足孔在口面形成不规则的一列, 一些在壳顶排列成三列············································································ 2
2. 步带靠外侧的次级板大并且和侧辐缝合线相接。赤道部步带区几乎与步带间区一样宽················································ 3
- 步带次级板小, 并且不与侧辐缝合线相接。赤道部步带区小于间步带区·················· 革海胆属Mortensen, 1903
3. 鳃缺失或退化。步带初级板通常不分裂··························································· 脆海胆属Mortensen, 1903
- 具鳃。步带初级板分裂成有孔区和无孔区···························································· 兜海胆属Koehler, 1897
4. 大疣在口面间步带的侧辐边缘与中央区丰富。步带区具大疣················································································· 5
- 口周围间步带侧辐边缘没有明显的大疣。大疣在步带和间步带的中央区缺失。围口部步带区的孔对单列·······················
············································································································ 脻海胆属Mortensen, 1903
5. 围口部管足孔对成四行排列。生殖孔在生殖板外侧膜隙中···················································································· 6
- 围口部管足孔对成双行排列。生殖孔在生殖板内··················································· 囊海胆属Grube, 1868
6. 具蛇首叉棘。间步带板间的膜隙仅在口面明显·················································· 秃海胆属Mortensen, 1934
- 蛇首叉棘缺失, 间步带板间的膜隙在口面与反口面都明显···································· 软海胆属Mortensen, 1903
由香莉, 2003. 黄、东海海胆分类学研究. 青岛: 中国科学院研究生院(海洋研究所)硕士学位论文, 6
吴古远, 付婉莹, 姚 旺等, 2017. 刻肋海胆科4种海胆叉棘超微结构的研究. 海洋湖沼通报, (4): 134—138
廖玉麟, 1982. 海胆生物学概况. 水产科学, (3): 1—8
Agassiz A, 1879. Preliminary report on the Echini of the Exploring Expedition of H.M.S “Challenger”. Sir C. Wyville Thomson Chief of Civilian Staff. Proceedings of the American Academy of Arts and Sciences,6: 190—212
Anderson O F, 2013. A review of New Zealand and southeast Australian echinothuriinids (Echinodermata: Echinothuriidae) with descriptions of seven new species. Zootaxa, 3609(6): 521—567
Anderson O F, 2016. A review of New Zealand and southeast Australian echinothurioids (Echinodermata: Echinothurioida)— excluding the subfamily Echinothuriinae—with a description of a new species of. Zootaxa, 4092(4): 451—488
Brosseau O, Eléaume M, 2003. Morphometric analysis of pedicellariae in(Philippi, 1845) (Echinodermata, Echinoidea, Cidaridae) using scanning electron microscopy (SEM). In: Echinoderm Research 2001: Proceedings of the 6th European Conference on Echinoderm Research, 137—142
Kroh A, Smith A B, 2010. The phylogeny and classification of post-Palaeozoic echinoids. Journal of Systematic Palaeontology, 8(2): 147—212
Mooi R, Constable H, Lockhart S, 2004. Echinothurioid phylogeny and the phylogenetic significance of(Echinoidea: Echinodermata). Deep Sea Research II: Topical Studies in Oceanography, 51(14—16): 1903—1919
Mortensen T, 1903. The Danish Ingolf-Expedition 1895-1896. Vol.4, No.2. Echinoidea, pt.1. Bianco Luno, Copenhagen, 198pp.page(s): 53, 63—64
Mortensen T, 1935. A Monorgraph of the Echinoidea, 2 (Bothriocidaroida, Melonechinoida, Lepidocentroida and Stirodonta). Copenhagen: Mortensen Th, pp.101—102 text-figs., 89pls
Ramalingam K, Ravindranath M H, Krishnan R, 1975. Histochemical studies of pedicellaria of an echinoid, Salmacis bicolor. Acta Histochemica, 52(2): 222—230
Smith A B, 2013. Key to genera of Echinothuriidae. In: Smith A B, Kroh A eds. The Echinoid Directory. World Wide Web Electronic Publication, available online at http://www. nhm.ac.uk/research-curation/research/projects/echinoid-directory/taxa/taxon.jsp?id=884
Tamori M, Koki J, Motokawa T, 2018. Dumbbell-shaped ossicles discovered in pedicellaria of flower sea urchins. Zoological Science, 35(1): 92—98
STUDY ON ULTRASTRUCTURE OF PEDICELLARIAE OF THREE GENERA IN FAMILY ECHINOTHURIIDAE
ZHENG Wan-Rui1, XIAO Ning2, AN Jian-Mei1
(1. School of Life Science, Shanxi Normal University, Linfen 041000, China; 2. Department of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China)
The pedicellariae of sea urchins can defend the invader and remove foreign matters on the surface. The shape of pedicellariae is one of the key characters for classification. The morphology characteristics of pedicellariae of,andin the western Pacific Ocean were studied by scanning electron microscope. A key to the genus of the Echinothuriidae was provided. Three common forms of pedicellariae in Echinothuriidae were recognized: tridentate pedicellarias, triphyllous pedicellarias, and dactylous pedicellarias. Tridentate pedicellarias and triphyllous pedicellarias occur in all three genera. In addition, dactylous pedicellarias was found inonly. The ultrastructure of triphyllous pedicellarias and dactylous pedicellarias changed slightly. The ultrastructural characteristics of tridentate pedicellarias were obviously different among species and genera. It is suggested that the type and structure of pedicellariae should be taken as the basis for the classification of the Echinothuriidae.
deep sea; echinoids; pedicellariae; morphology; SEM
Q959.268
10.11693/hyhz20191100207
* 中国科学院战略生物资源计划, KFJ-BRP-017-38号; 中国科学院战略性先导A类专项, XDA22050203号。郑婉瑞, 硕士研究生, E-mail: 1670772386@qq.com
肖 宁, 博士, E-mail: xiaoning@qdio.ac.cn; 安建梅, 硕士生导师, 教授, E-mail: anjianmei@sxnu.edu.cn
2019-11-07,
2020-01-27
