底质细化对厦门潮下带文昌鱼栖息地大型底栖动物群落的影响*
2020-05-25陈丙温蔡立哲饶义勇李文君陈昕韡傅素晶彭文晴
陈丙温 蔡立哲, 2 饶义勇 李文君 陈昕韡 傅素晶 彭文晴 郑 斌
底质细化对厦门潮下带文昌鱼栖息地大型底栖动物群落的影响*
陈丙温1蔡立哲1, 2①饶义勇1李文君1陈昕韡1傅素晶1彭文晴1郑 斌1
(1. 厦门大学环境与生态学院 厦门 361102; 2. 厦门大学滨海湿地生态系统教育部重点实验室 厦门 361102)
本文根据2014年至2017年在厦门文昌鱼保护区黄厝、南线十八线、鳄鱼屿和小嶝岛四处潮下带以及参照点白城潮下带获得的大型底栖动物和沉积物粒径数据, 探讨底质细化对厦门潮下带文昌鱼栖息地大型底栖动物群落的影响。聚类分析表明个别黏土含量较高的取样站没有发现文昌鱼且大型底栖动物种数少。单因素方差分析表明, 厦门潮下带部分大型底栖动物群落参数和优势种数量存在显著差异。底质粒径分析表明, 2017年9月, 鳄鱼屿和白城潮下带黏土和粉砂含量高于黄厝、南线十八线和小嶝岛潮下带的黏土和粉砂含量。2016年夏季黄厝、南线十八线潮下带黏土和粉砂含量明显高于2014年夏季的黏土和粉砂含量。相关分析表明, 5个大型底栖动物群落参数如物种数、栖息密度、生物量、多样性指数(′)和丰度指数(), 7个常见种的栖息密度和生物量与粉砂和黏土含量呈显著正相关, 均匀度指数()和4种常见种的数量与粉砂和黏土含量无显著相关, 白氏文昌鱼()的栖息密度和生物量与粉砂和黏土含量呈显著负相关。上述结果证实, 黏土含量增加到15%和粉砂含量增加到60%有利于多数大型底栖动物栖息密度和生物量的增加, 但导致白氏文昌鱼栖息密度和生物量的减少, 而红角沙蚕()、滑指矶沙蚕()、光滑倍棘蛇尾()受底质细化的影响较小。
大型底栖动物; 群落结构; 文昌鱼栖息地; 底质细化; 潮下带; 厦门
海洋底栖生物中许多种类是经济鱼虾蟹的天然饵料, 有些是水产养殖和捕捞对象, 而且底栖生物还是生物监测的敏感指示生物(蔡立哲, 2006)。大型底栖动物是海洋生态系统物质循环和能量流动中积极的参与者(马骏等, 2010)。有关海洋大型底栖动物群落生态学的研究主要涉及群落的物种组成及分布(邓玉娟等, 2016; 王金宝等, 2017)、群落优势种的数量分布及其季节变化(李荣冠等, 1999; 彭松耀等, 2017)。大型底栖动物与其生活的底质沉积生境有密切关系(杜飞雁等, 2008)。台湾海峡西部近海大型底栖生物与沉积物有相当密切的关系, 在不同类型的沉积物中, 种类和数量都有显著的差别(江锦祥等, 1984)。大型底栖动物经常被作为监测海洋污染的生态类群(李荣冠等, 1992)。
1991年厦门市建立了文昌鱼自然保护区, 1999年厦门文昌鱼与中华白海豚、白鹭一起申报“厦门海洋珍稀物种国家级自然保护区”, 并于2000年获得国务院批准。长期以来, 人们认为厦门文昌鱼资源量减少的原因之一是底质变化。早在1953年, 金德祥等(1953)就阐述了文昌鱼分布与底质粒径的关系。周秋麟等(1990)探讨了厦门黄厝海区文昌鱼的分布及其与底质的关系。文昌鱼栖息地底质类型主要是砂质, 但不是纯砂, 而是粗细砂掺杂, 以中粗砂为主(曾国寿等, 1996; 吕小梅等, 1997)。砂含量高值区是适合于文昌鱼生存的区域(吕小梅等, 2005)。随着高集海堤的兴建以及刘五店文昌鱼渔场附近的东坑、策槽等地大面积的滩涂围垦, 改变了海域的水动力条件, 使大量淤泥不断在文昌鱼渔场沉积, 致使文昌鱼赖以生存的砂质沉积物环境逐渐被泥质环境所取代(翁朝红等, 2012)。
综上所述, 无论是大型底栖动物群落生态学研究, 还是文昌鱼种群与底质关系的研究, 均说明大型底栖动物与其生活的底质沉积生境有密切关系(杜飞雁等, 2008), 而具体存在什么样的关系是本文要探讨的内容。本文根据2017年9月在厦门白城、黄厝、南线十八线(简称南线)、鳄鱼屿和小嶝岛5处潮下带(其中后4处潮下带位于文昌鱼保护区)获得的大型底栖动物和沉积物粒径数据, 以及2014年和2016年夏季在黄厝、南线十八线潮下带获得的大型底栖动物和沉积物粒径数据, 探讨底质细化对厦门潮下带文昌鱼栖息地大型底栖动物群落的影响。
1 材料与方法
厦门文昌鱼自然保护区由四块海区组成, 共有58km2, 其中黄厝海区划为核心区, 保护面积10km2; 南线十八线海区划为经管区, 保护面积为35km2; 刘五店附近的鳄鱼屿海区划为实验区, 保护面积为5km2; 小嶝岛-角屿岛海区为核心区, 保护面积为8km2(方少华等, 2002)。
2017年9月, 在厦门文昌鱼保护区的上述四块海区以及白城海区(参照海区)共43个取样站进行了大型底栖动物和沉积物底质粒径调查。为了比较的科学性, 在厦门五处海区分别取6个取样站的数据进行大型底栖动物群落参数、优势种数量和沉积物粒径比较, 所选取的1—6取样站(白城潮下带)、7—12取样站(黄厝潮下带)、20—22和24—26取样站(南线-十八线潮下带)、35—40取样站(鳄鱼屿潮下带)、42—47取样站(小嶝岛潮下带)见图1。大型底栖动物调查采用0.05m2抓斗式采泥器, 每个取样站采集4次。将沉积物倒入塑料桶内, 加水搅拌, 悬浮液用孔径0.5mm的筛网过滤, 重复淘洗直至塑料桶内的水清澈透明, 将截留在筛网上的大型底栖动物及残渣装入样品瓶中, 用5%的甲醛溶液固定后带回实验室。在实验室进行样品再分选, 即带回的样品同样置于孔径0.5mm的网筛内, 小心地用自来水冲洗, 而后把留在筛网上的大型底栖动物连同残渣用自来水转移至大培养皿, 用镊子将大型底栖动物(含文昌鱼)从残渣中挑出, 放入盛有质量分数75%的酒精的塑料透明瓶, 待鉴定。在解剖镜下进行标本的鉴定、记数后用滤纸吸干, 用感量为0.0001g的电子天平称湿重。
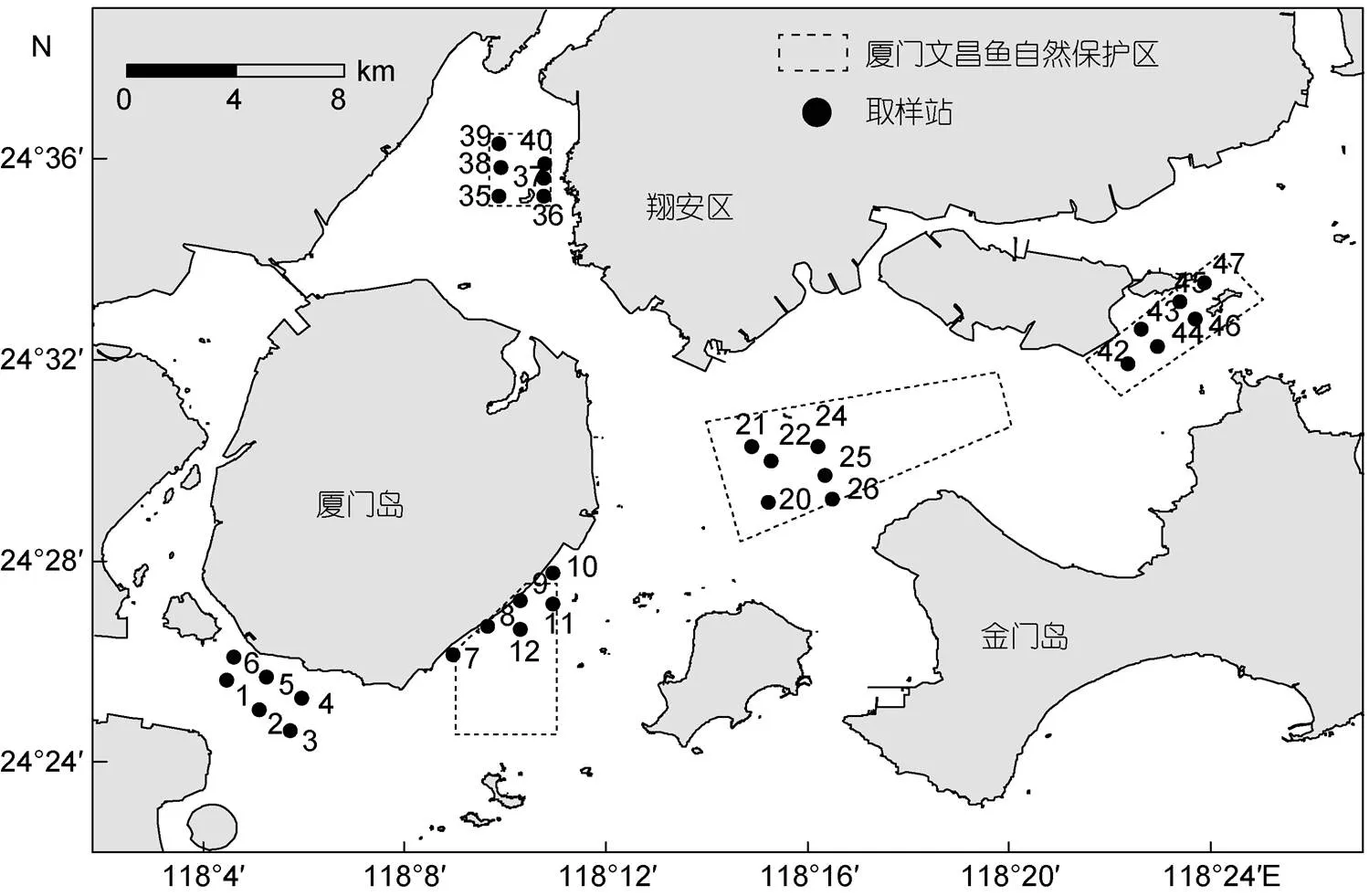
图1 2017年9月厦门潮下带文昌鱼和大型底栖动物取样站位示意图
2014年夏季(2014年8月)和2016年夏季(2016年7月)在厦门文昌鱼自然保护区的黄厝潮下带设置9个取样站(H1—H9)和南线十八线潮下带设置9个取样站(S1—S9)进行大型底栖动物(含文昌鱼)和沉积物粒径调查。大型底栖动物的采集均在水深约1—2m的潮下带进行, 使用面积为32cm×34cm的耙式采样器, 泥沙样品经0.5mm孔径网筛分选后, 留存在筛网上的文昌鱼及其他大型底栖动物以及一些杂质用5%的中性海水甲醛溶液固定在对应的瓶子中带回实验室(饶义勇等, 2015)。在每个取样站取3份表层沉积物样品用于分析沉积物的粒度特征。
运用 Primer 5.0 软件计算大型底栖动物群落种类多样性指数(′)、均匀度指数()、丰富度指数()和群落的聚类分析; 利用SPSS17.0 软件进行群落参数、优势种栖息密度和生物量的方差分析和相关分析。
2 结果
2.1 研究区域沉积物粒径的时空变化
2014年夏季与2016年夏季比较可以看出, 厦门黄厝潮下带黏土和粉砂含量明显增加, 黏土含量平均值从0.10%上升到3.43%, 粉砂含量从0.51%上升到57.45%; 南线潮下带黏土和粉砂含量也明显增加, 黏土含量从0.28%上升到3.01%, 粉砂含量从3.17%上升到88.25%。相反地, 厦门黄厝潮下带砂和砾石含量明显减少, 砂含量从58.29%下降至32.61%, 砾石含量从41.10%下降至6.51%; 南线潮下带砂和砾石含量也明显减少, 砂含量从83.83%下降至7.33%, 砾石含量从12.72%下降至1.40% (图2)。
2017年9月, 厦门三处有文昌鱼栖息的黄厝、南线和小嶝岛潮下带的黏土含量分别是6.82%、4.62%和4.40%, 未发现文昌鱼分布的白城和鳄鱼屿潮下带黏土含量分别是14.42%和15.92%, 文昌鱼栖息的潮下带站位黏土含量较低; 粉砂含量的分布与黏土含量分布一致, 即有文昌鱼栖息的潮下带粉砂含量较低, 黄厝、南线和小嶝岛潮下带的砂含量分别是59.52%、57.80%和64.95%, 未发现文昌鱼分布的白城和鳄鱼屿潮下带站位砂含量分别是19.42%和14.04%, 有文昌鱼栖息的潮下带砂含量较高; 砾石含量的分布与砂含量分布一致(图2)。
从黄厝和南线潮下带底质粒径组成看, 粉砂含量从2014年夏季、2016年夏季和2017年9月的变化是先增加后减少, 黏土含量则是随时间推移而增加。但需要说明的是2014年夏季与2016年夏季在黄厝和南线的取样站和取样范围是一样的, 但2017年9月在黄厝和南线的取样站和取样范围与2014年夏季和2016年夏季在黄厝和南线的取样站和取样范围是不完全一样的, 2014年和2016年夏季的取样范围较小, 基本上均有文昌鱼分布, 而2017年9月的取样范围较大, 大概只有50%的取样站有文昌鱼分布。因此, 2017年9月与2014年和2016年夏季底质粒径的比较只能作为参考值。

图2 厦门五处潮下带底质粒径组成的时空分布
2.2 研究区域大型底栖动物群落的时空变化
2014年夏季厦门文昌鱼保护区黄厝潮下带的大型底栖动物物种数、栖息密度、生物量、多样性指数(′)、均匀度指数()和丰度指数()均比2016年夏季的低, 分别是23和89种, 80.3和298.9ind./m2, 13.17和17.57g/m2, 1.728和3.579, 0.630和0.815, 1.239和4.038。2014年夏季南线十八线潮下带的大型底栖动物物种数、栖息密度、多样性指数(′)、均匀度指数()和丰度指数()也均比2016年夏季的低, 分别是25和57种, 76.2和195.1ind./m2, 1.635和2.943, 0.620和0.715, 1.313和3.201(表1)。
根据2014年夏季和2016年夏季在黄厝和南线十八线35个取样站的大型底栖动物数据进行聚类分析得出, 在相似度20%处可分为两个大型底栖动物群落, 群落一位于2014年夏季的南线十八线潮下带S6取样站, 该取样站没有采集到文昌鱼, 优势种是红角沙蚕()和毛盲蟹(), 黏土含量在夏季所有测站中含量最高(1.43%)。群落二包括除了2014年夏季S6取样站外的所有取样站(图3), 群落二的优势种有白氏文昌鱼()、光滑倍棘蛇尾()和红角沙蚕等。
2017年9月, 厦门文昌鱼保护区黄厝、南线、鳄鱼屿和小嶝岛潮下带的大型底栖动物物种数分别是83种、80种、68种和54种, 均比对照区白城潮下带(97种)要低。厦门文昌鱼保护区黄厝、南线、鳄鱼屿和小嶝岛潮下带的大型底栖动物栖息密度、多样性指数(′)和丰度指数()也均比对照区白城潮下带的低(表2)。
表1 厦门潮下带两处海域大型底栖动物物种数合计及其他群落参数数值(平均值±标准差)
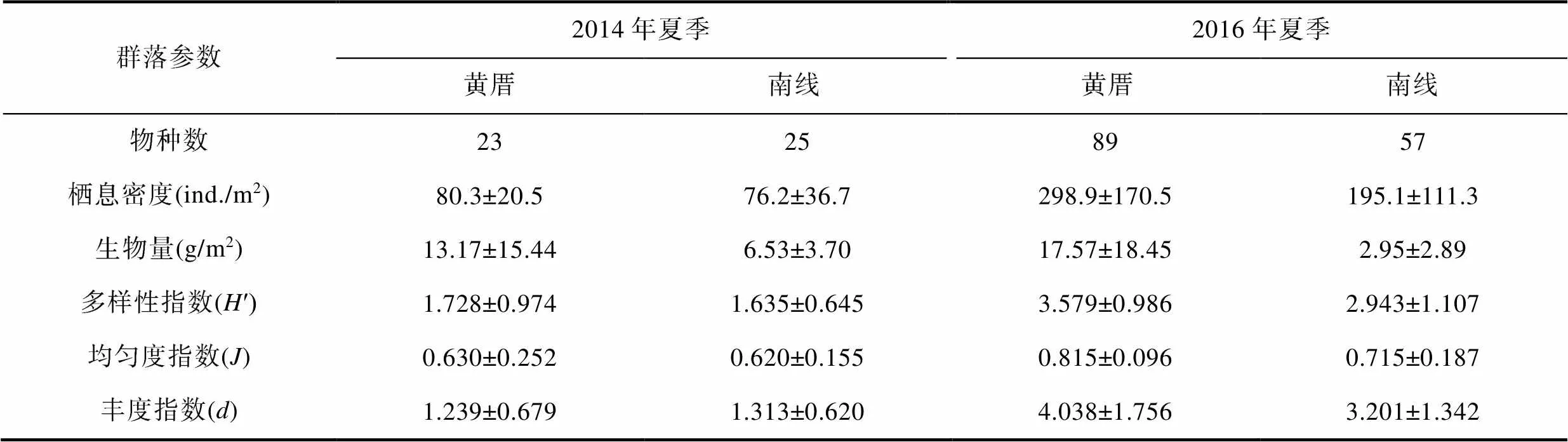
Tab.1 Benthic macrofaunal species number (total value) and other community parameters (average value±standard deviation) from two subtidal areas in Xiamen
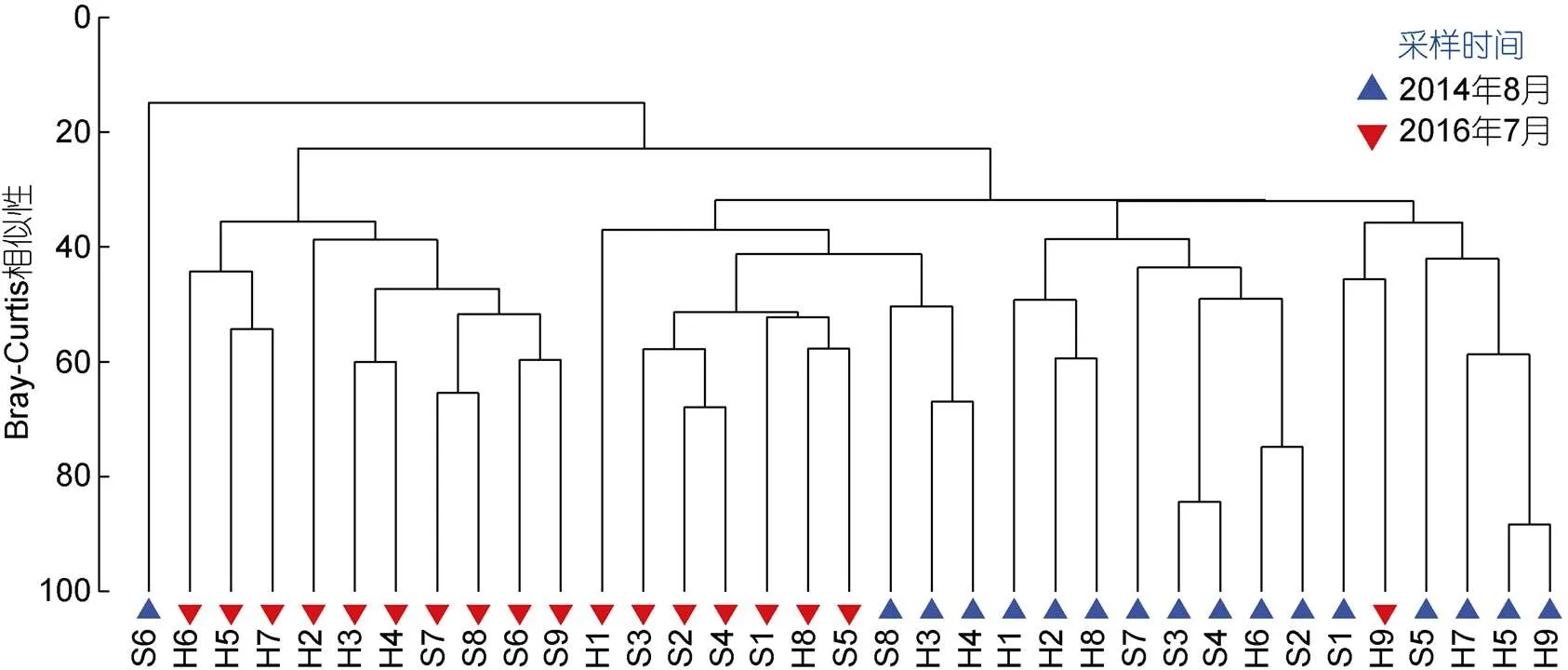
图3 厦门海域2014年夏季和2016年夏季两处潮下带35个取样站的聚类分析
表2 2017年9月厦门潮下带五处海域大型底栖动物物种数合计及其他群落参数的数值(平均值±标准差)
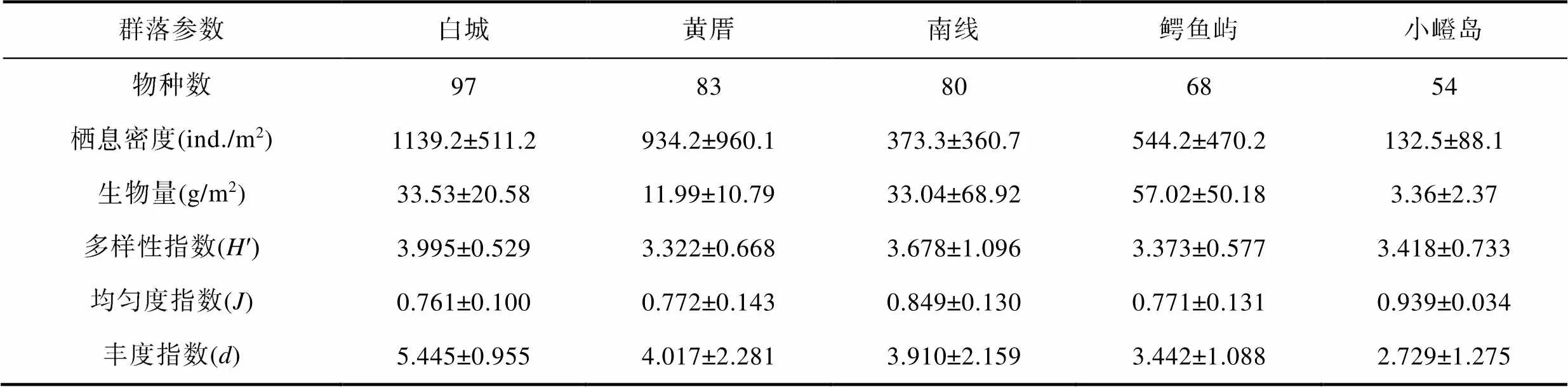
Tab.2 Benthic macrofaunal species number (total value) and other community parameters (average value±standard deviation) from five subtidal areas in Xiamen in September 2017
根据2017年9月在厦门潮下带五处海域30个取样站的大型底栖动物数据进行聚类分析得出, 在相似度20%处可分为两个大型底栖动物群落, 群落一位于小嶝岛潮下带的44和46取样站, 这两个取样站大型底栖动物物种少, 仅有常见种双鳃内卷齿蚕()和模糊新短眼蟹()等, 且44取样站的黏土含量高达16.43%。群落二包括白城、黄厝、南线和鳄鱼屿潮下带的各6个取样站以及小嶝岛潮下带的其余4个取样站(图4), 优势种是刚鳃虫()和沙钩虾。
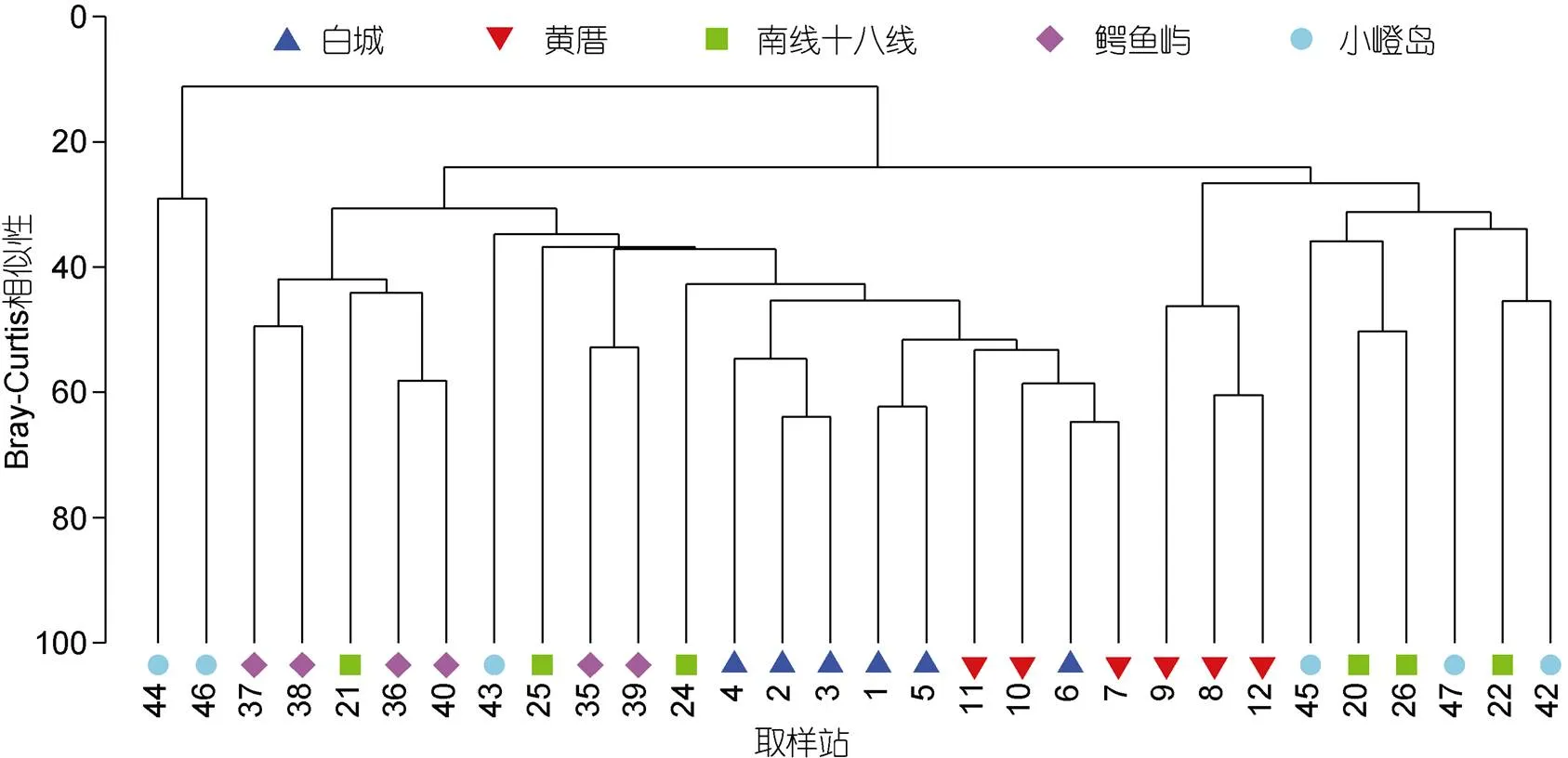
图4 2017年9月厦门五处潮下带30个取样站的聚类分析
2.3 研究区域大型底栖动物常见种栖息密度和生物量的时空变化
2014年夏季, 在黄厝潮下带未采集到背蚓虫()、刚鳃虫、双鳃内卷齿蚕、太平洋稚齿虫()、滑指矶沙蚕()、菲律宾蛤仔()和沙钩虾(spp.), 但在2016年夏季均有采集到。2014年夏季, 在南线十八线潮下带未采集到太平洋稚齿虫、滑指矶沙蚕、菲律宾蛤仔和沙钩虾, 但在2016年夏季均有采集到, 在南线十八线潮下带, 2014年夏季和2016年夏季均未采集到刚鳃虫。红角沙蚕、光滑倍棘蛇尾和白氏文昌鱼在黄厝和南线潮下带以及2014年夏季和2016年夏季均有采集到。背蚓虫、双鳃内卷齿蚕和短竹蛏()在2014年夏季的黄厝潮下带未采集到。模糊新短眼蟹在2016年夏季的南线潮下带未采集到(表3)。
表3 厦门黄厝和南线潮下带海域大型底栖动物常见种的平均栖息密度和生物量
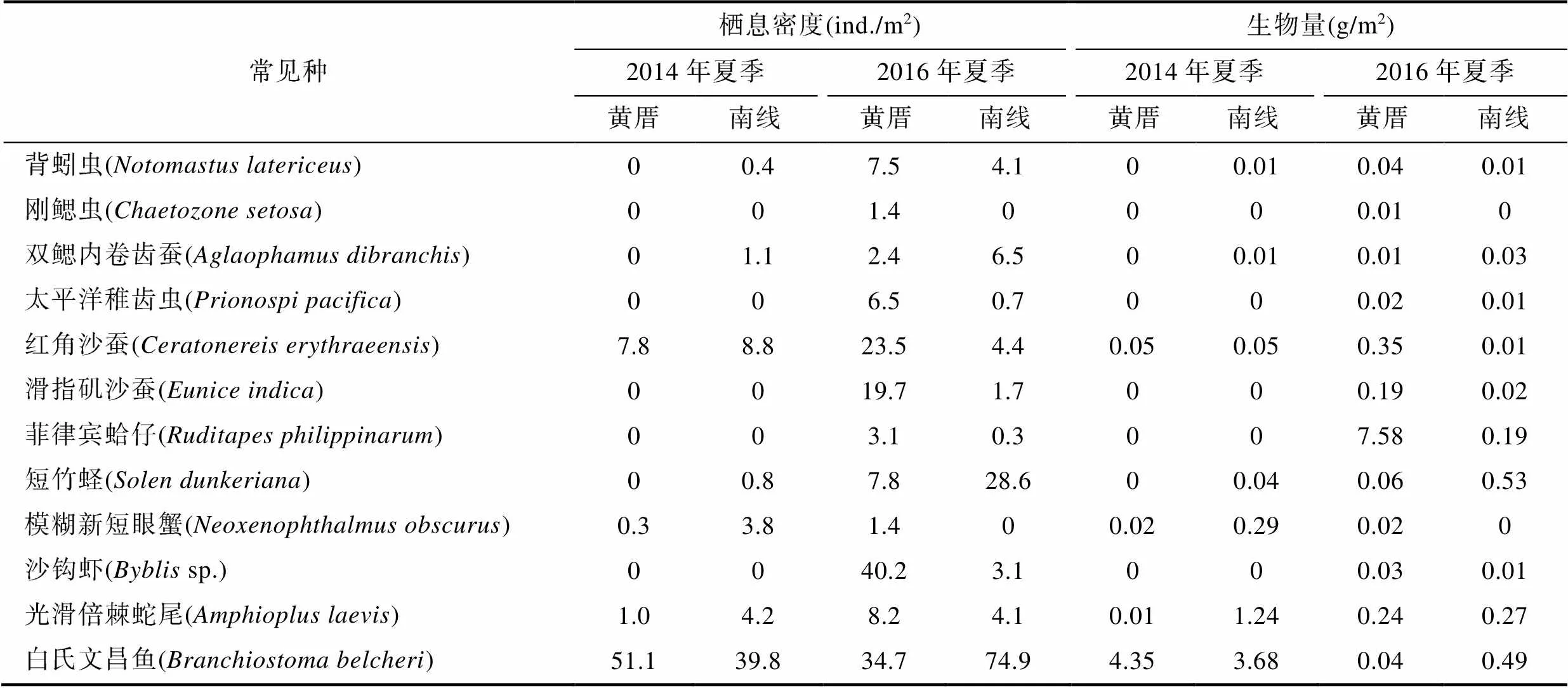
Tab.3 The average density and biomass of the common species of benthic macrofauna in Xiamen Huangcuo and Nanxian subtidal areas
2017年9月, 在鳄鱼屿潮下带海区未采集到白氏文昌鱼和模糊新短眼蟹, 该区域红角沙蚕的栖息密度是五处潮下带海区最低的, 而刚鳃虫、菲律宾蛤仔和光滑倍棘蛇尾栖息密度是五处潮下带海区最高的。在小嶝岛潮下带, 背蚓虫、刚鳃虫、双鳃内卷齿蚕、太平洋稚齿虫和菲律宾蛤仔的栖息密度是五处潮下带海区最低的; 在三处有白氏文昌鱼分布的黄厝、南线和小嶝岛潮下带, 小嶝岛潮下带的白氏文昌鱼栖息密度是最低的(表4)。
2017年9月在鳄鱼屿潮下带, 红角沙蚕的生物量是五处潮下带海区最低的, 而刚鳃虫、菲律宾蛤仔和光滑倍棘蛇尾生物量是五处潮下带海区最高的。在小嶝岛潮下带, 背蚓虫、刚鳃虫、太平洋稚齿虫、菲律宾蛤仔、短竹蛏和沙钩虾的生物量是五处潮下带海区最低的; 在三处有白氏文昌鱼分布的黄厝、南线和小嶝岛潮下带, 小嶝岛潮下带的白氏文昌鱼的生物量均是最低的(表4)。
表4 2017年9月厦门潮下带五处海域大型底栖动物优势种栖息密度和生物量数值(平均值±标准差)
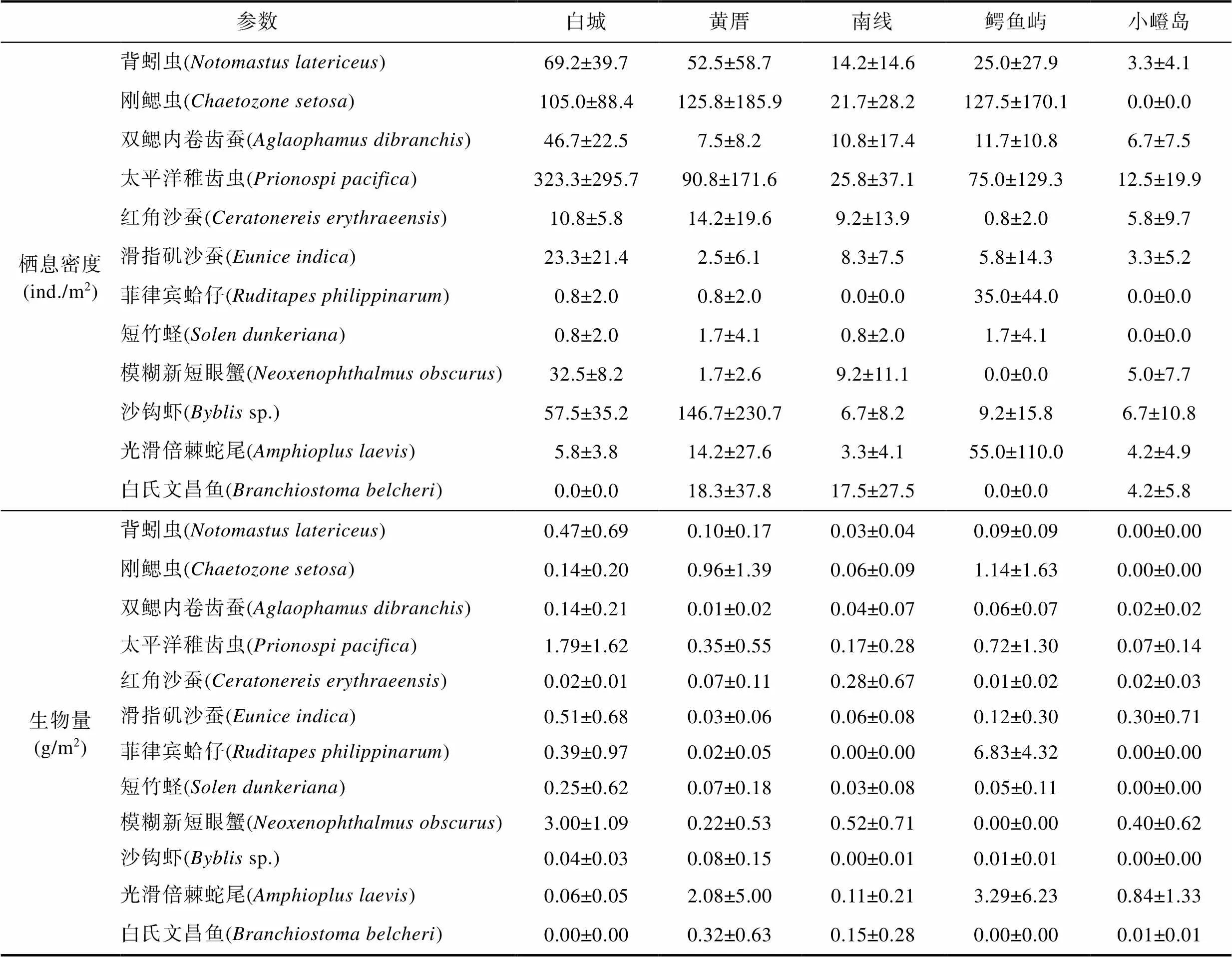
Tab.4 The values (average value±standard deviation) of the density and biomass of the dominant species in Xiamen five subtidal areas in September 2017
2.4 研究区域大型底栖动物群落和种群参数时空变化的方差分析
单因素方差分析表明, 除了大型底栖动物生物量以外, 2014年夏季与2016年夏季其余5类大型底栖动物群落参数均呈显著差异。优势种背蚓虫、双鳃内卷齿蚕、太平洋稚齿虫、滑指矶沙蚕和沙钩虾的栖息密度在2014年夏季与2016年夏季之间呈显著差异; 双鳃内卷齿蚕、太平洋稚齿虫、沙沟虾和厦门文昌鱼的生物量在2014年夏季与2016年夏季之间呈显著差异。大型底栖动物生物量、双鳃内卷齿蚕栖息密度和生物量、滑指矶沙蚕栖息密度、沙钩虾栖息密度和生物量在黄厝潮下带和南线十八线之间呈显著差异(表5)。
单因素方差分析表明, 2017年9月厦门潮下带海域6个大型底栖动物群落参数中, 仅大型底栖动物栖息密度在五处潮下带之间呈显著差异。优势种背蚓虫、双鳃内卷齿蚕、太平洋稚齿虫、滑指矶沙蚕、菲律宾蛤仔和模糊新短眼蟹的栖息密度在五处潮下带之间呈显著差异; 太平洋稚齿虫、菲律宾蛤仔和模糊新短眼蟹的生物量在五处潮下带之间呈显著差异(表6)。
表5 厦门黄厝和南线潮下带大型底栖动物群落和常见种种群参数的方差分析
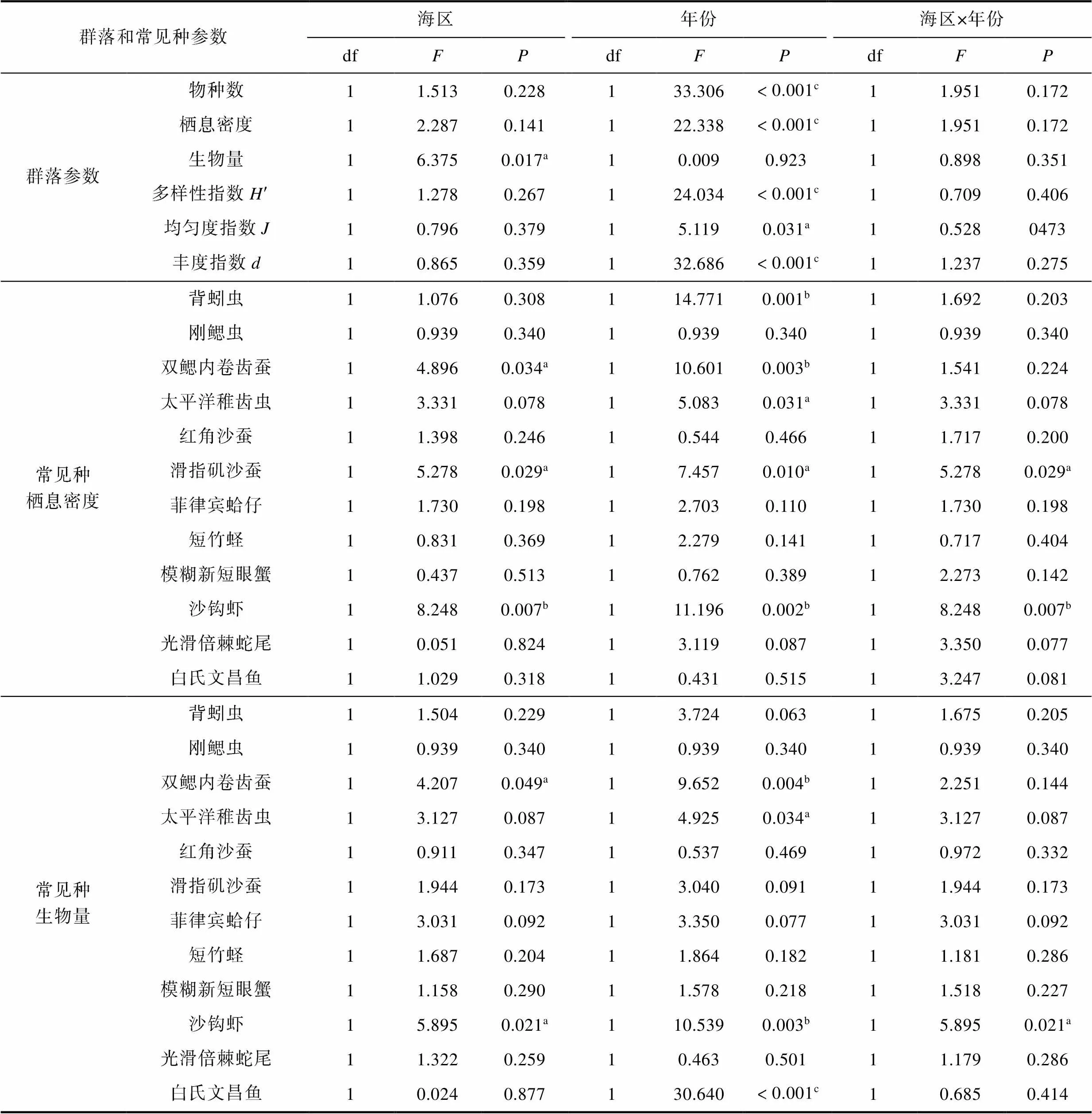
Tab.5 Variance analysis of community and population parameters of benthic macrofauna and common species in Xiamen Huangcuo and Nanxian subtidal areas
注:a: 0.05 水平显著差异;b: 0.01水平显著差异;c: 0.001水平显著差异.
2.5 研究区域大型底栖动物种群和群落参数与底质粒径参数的相关分析
根据2017年9月、2016年夏季和2014年夏季共65个站次的大型底栖动物群落参数、优势种栖息密度和生物量、沉积物粒径参数进行相关分析。结果表明, 除了均匀度指数, 其余5类大型底栖动物群落参数, 即物种数、栖息密度、多样性指数(′)、均匀度指数()和丰度指数()与底质粒径各组分含量呈显著相关。然而, 红角沙蚕、滑指矶沙蚕、短竹蛏和光滑倍棘蛇尾的栖息密度与砾、砂、粉砂和黏土含量均无显著相关; 红角沙蚕、滑指矶沙蚕、菲律宾蛤仔、短竹蛏和光滑倍棘蛇尾的生物量与砾、砂、粉砂和黏土含量也均无显著相关(表7)。
表6 2017年9月厦门潮下带五处海域大型底栖动物群落和常见种种群参数的方差分析
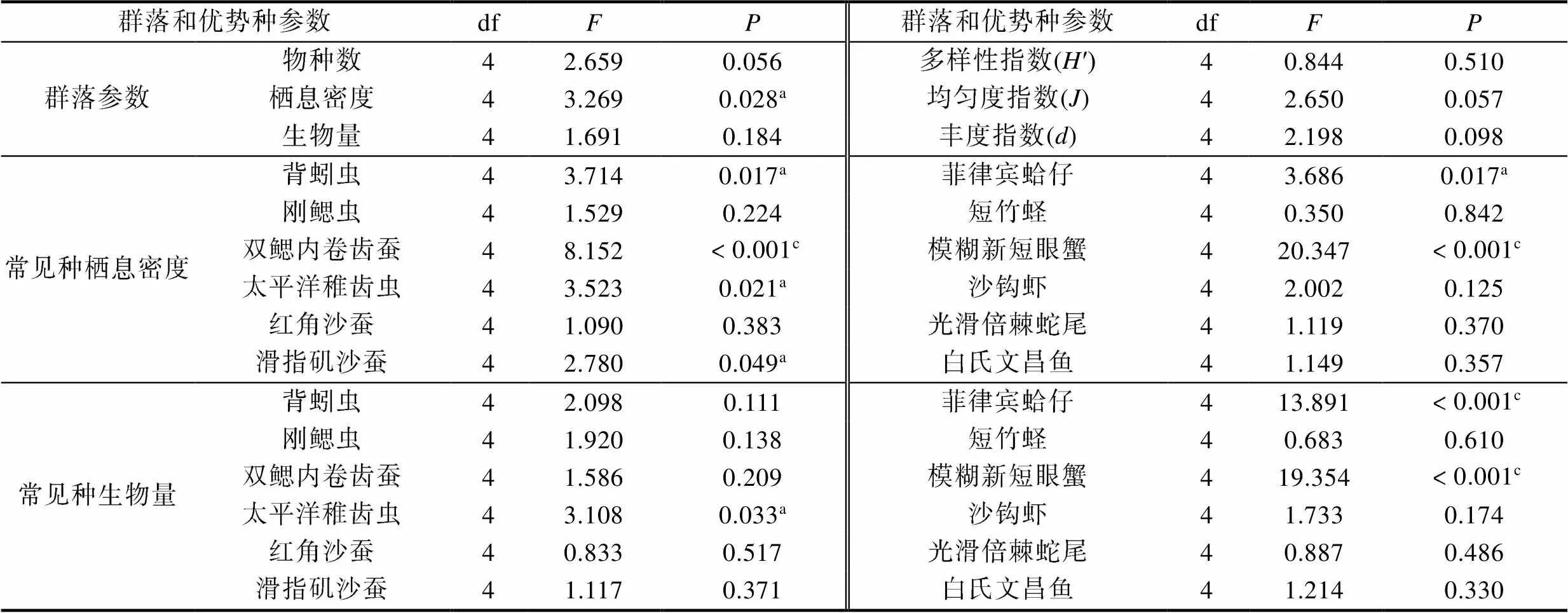
Tab.6 Variance analysis of community and population parameters of benthic macrofauna and common species in Xiamen five subtidal areas in September 2017
表7 大型底栖动物的群落参数和常见种的种群参数与底质粒径参数的相关系数(=65)

Tab.7 Correlation coefficients among benthic macrofaunal community parameters, the common species population parameters, and sediment grain size parameters
3 讨论
3.1 底质细化对大型底栖动物群落的影响
在本研究中, 潮下带细沙底质的大型底栖动物栖息密度和生物量较泥砂底质的大型底栖动物栖息密度和生物量低。2017年9月, 黄厝和南线十八线潮下带粉砂的平均含量分别是29.00%和23.27%, 而鳄鱼屿潮下带粉砂的平均含量是68.53%, 鳄鱼屿潮下带粉砂含量显著高于黄厝和南线十八线潮下带粉砂。黄厝和南线十八线潮下带大型底栖动物平均生物量分别是11.99和33.04g/m2, 而鳄鱼屿潮下带大型底栖动物平均生物量是57.02g/m2, 鳄鱼屿潮下带大型底栖动物平均生物量显著高于黄厝和南线十八线潮下带的大型底栖生物量。2014年夏季与2016年夏季的大型底栖动物物种数和栖息密度, 也证实了潮下带中细沙底质大型底栖动物栖息密度较泥砂底质的大型底栖动物栖息密度低。许多前人研究也证实了潮下带中细沙底质大型底栖动物生物量较泥砂底质的大型底栖动物生物量低, 如台湾海峡西部近海中细砂底质大型底栖动物生物量仅5.8g/m2, 而泥沙底质大型底栖动物生物量达24.7g/m2(江锦祥等, 1984); 台湾海峡北部多毛类高生物量和高栖息密度区出现在含粉砂量60%左右的沉积环境中, 其沉积环境中有机物含量丰富, 有利于吞咽型多毛类的大量繁衍(孙道元等, 1988); 兴化湾黏土质粉砂沉积物的大型底栖动物物种数较砂质沉积物的大型底栖动物物种数多(李荣冠等, 1999); 随着底质细化和底层溶解氧增加, 路易斯安那大陆架沙洲的大型底栖动物物种多样性和栖息密度显著增加(Dubois, 2009); 由于沉积物粒度的不同, 香港大浪湾(Tai Long Wan)和白腊湾(Pak Lap Wan)的大型底栖动物群落结构存在明显差异(Chen, 2013); 2014年夏季在厦门海域南线十八线潮下带的S6 取样站, 粉砂含量高达85.13%, 未能采集到文昌鱼, 但却采集到红角沙蚕和双鳃内卷齿蚕(饶义勇等, 2015); 由于底质的细化, 在厦门黄厝潮下带发现菲律宾蛤仔数量增加, 而这种现象在河北昌黎海域也发现(郭兴然等, 2019)。
在小嶝岛潮下带, 虽然粉砂平均含量与黄厝和南线十八线潮下带的粉砂平均含量接近, 但小嶝岛潮下带受厦门新机场建设及海岸工程的影响, 未发现刚鳃虫和菲律宾蛤仔等大型底栖动物, 说明小嶝岛潮下带受新的沉积物覆盖, 不利于刚鳃虫的大量繁衍; 新粉砂的沉积同时也不利于菲律宾蛤仔的摄食。因此, 2017年9月小嶝岛的大型底栖动物物种数、栖息密度和生物量均显著低于黄厝和南线十八线潮下带。
底质类型是影响大型底栖动物分布的重要环境因子(李晓静等, 2017)。近岸大型底栖动物群落的分布格局与沉积相类型密切相关(江锦祥等, 1984; 李亚芳等, 2018)。本研究发现黏土含量增加到15%和粉砂含量增加到60%有利于多数大型底栖动物栖息密度和生物量的增加, 而黏土含量超过60%则不利于多数大型底栖动物的栖息和生长。
厦门潮下带海域红角沙蚕、滑指矶沙蚕和光滑倍棘蛇尾受底质细化的影响较小。本研究红角沙蚕、滑指矶沙蚕、短竹蛏和光滑倍棘蛇尾的栖息密度与砾、砂、粉砂和黏土含量均无显著相关。倍棘蛇尾(含中华倍棘蛇尾和倍棘蛇尾未定种)不仅是粤西海陵湾湾内大型底栖动物群落的优势种, 也是湾外大型底栖动物群落的优势种, 还是周年优势种(李亚芳等, 2018)。在渤海湾曹妃甸围填海工程附近海域日本倍棘蛇尾()2013年和2014年的优势度分别为0.036和0.38, 变化较小(李晓静等, 2017)。矶沙蚕属()不仅是香港含沙量较高海域的优势属(Chen, 2013), 也是粉砂和黏土含量较高海域的优势属(唐盟等, 2017; 李亚芳等, 2018)。而本研究显示, 2017年9月, 除了小嶝岛海区未采集到红角沙蚕外, 白城、黄厝、南线十八线和鳄鱼屿海区均采集到红角沙蚕, 且各海区之间不呈显著性差异。可以推测红角沙蚕、滑指矶沙蚕和光滑倍棘蛇尾对底质类型的生态位较宽。生态位较宽的生物与其他生物之间的重叠较为明显(刘好真等, 2018)。
沉积物内有机质含量往往与沉积物粒径大小呈负相关关系。黏土和粉砂含量较高的沉积物比表面积较大, 吸附有机质能力较强, 沉积物内丰富的有机质可以为大型底栖动物提供丰富的食物来源, 从而可导致大型底栖动物栖息密度的增加(Wang, 2016)。深圳湾潮间带羽须鳃沙蚕()和腺带刺沙蚕()的栖息密度和生物量与有机质含量呈显著负相关, 而有机质含量与黏土含量呈显著正相关(Cai, 2013)。有机质污染将导致文昌鱼栖息地中多毛类种类的增多(Antoniadou, 2004), 同时也将导致文昌鱼种群数量减少(Rota, 2009)。
影响大型底栖动物分布的因素很多, 除了沉积物类型外, 还有水深、溶解氧、营养盐、有机质或海岸工程、水产养殖等人类活动的影响。网箱养殖产生的有机污染已使大鹏澳网箱养殖区沉积环境严重恶化, 养殖区大型底栖动物出现种数、生物量和个体数量明显低于对照区,其群落特征也与对照区有明显不同(黄洪辉等, 2005)。天津大沽口海域围海造陆工程对近岸海域生态环境的影响时发现, 围填海产生的主要污染物为悬浮物, 而悬浮物浓度与大型底栖动物的栖息密度呈显著的负相关关系(刘春琳等, 2019)。温度、盐度、水深、溶解氧是浙江檀头山岛周围海域对大型底栖动物影响程度较大的环境因子(刘迅等, 2017)。烟台近海大型底栖动物群落与底质和水深关系密切(王全超等, 2013)。围填海、采沙、水产养殖等是影响厦门潮下带海域大型底栖动物, 特别是影响文昌鱼的环境因子(周秋麟等, 1990; 方少华等, 2002; 饶义勇等, 2015)。
3.2 底质细化对文昌鱼种群的影响
厦门湾西部海域受到较强程度的陆源排污影响, 海域环境质量较差。东部海域靠近外海, 水动力交换较强, 陆源污染物被稀释较多, 环境质量较好(黄昆等, 2019)。本文调查结果表明, 文昌鱼主要分布在环境质量较好的东部海域, 而环境质量相对较差的区域文昌鱼栖息密度相对较低。近年来在厦门文昌鱼保护区出现的非法养殖与非法清淤活动, 严重破坏了文昌鱼的栖息环境。此外, 沉积物底质粒径对文昌鱼生存、分布具有重要影响, 该种生物喜好比较粗而疏松的砂质沉积物, 而泥质沉积物则不适合文昌鱼的生存。研究表明泥质底质有机质含量丰富, 水质浑浊, 不利于文昌鱼种群栖息(金德祥等, 1953); 底质有机质含量的高低与粒度的大小往往成负相关关系, 有机质含量最高的取样站为粉砂质泥底质类型, 有机质含量最低的取样站为中粗砂(周秋麟等, 1990); 文昌鱼栖息地底质类型以粗中砂为主, 有机质含量较低(曾国寿等, 1996); 文昌鱼()喜好栖息在有碎贝壳和砾石的沉积物中(Antoniadou, 2004); 文昌鱼喜欢栖息在含量为77.8%—99.4%、中值粒径为0.5—2.0的砂和砾沉积物中(Chen, 2013), 文昌鱼栖息地常被称为“文昌鱼沙(amphioxus sand)” (Antoniadou, 2004)。此外, 研究还表明鳄鱼屿周边的牡蛎吊养使海区沉积大量的污泥, 改变了文昌鱼生活的栖息地底质结构, 使文昌鱼无法生存(翁朝红等, 2012); 文昌鱼栖息密度和生物量与底质粒径分选系数呈显著负相关(饶义勇等, 2015); 文昌鱼对于有机质污染和底质质量的改变十分敏感(Rota, 2009; de Moura Barboza, 2013); 昌黎生态监控区适合文昌鱼生存的砂质底所占百分比逐年退化, 保护动物文昌鱼生物量呈退化趋势(高敏等, 2017); 昌黎海域近年来加速发展海上养殖, 养殖病害及养殖贝类粪沉降也加剧了青岛文昌鱼栖息环境的改变(郭兴然等, 2019)。本研究区域黏土平均含量变幅在0—16%之间、粉砂平均含量变幅在0—70%之间, 且文昌鱼栖息密度和生物量均与黏土和粉砂含量呈显著负相关, 证实黏土含量增加到15%和粉砂含量增加到60%导致白氏文昌鱼栖息密度和生物量的减少。
4 结论
底质细化对厦门潮下带文昌鱼栖息地的大型底栖动物群落产生了明显的影响, 不同物种对底质细化具有不同的响应。相关分析表明, 5个大型底栖动物群落参数如物种数、栖息密度、生物量、多样性指数(′)和丰度指数(), 以及7个常见种的栖息密度和生物量与粉砂和黏土含量呈显著正相关, 均匀度指数()和4种常见种的数量与粉砂和黏土含量无显著相关, 白氏文昌鱼的栖息密度和生物量与粉砂和黏土含量呈显著负相关。底质细化导致喜砂的白氏文昌鱼的数量明显下降, 却导致背蚓虫、刚鳃虫、双鳃内卷齿蚕、太平洋稚齿虫、菲律宾蛤仔和沙钩虾等物种数量增加, 而红角沙蚕、滑指矶沙蚕、光滑倍棘蛇尾受底质细化的影响较小。
致谢 感谢在实验室工作和学习过的张慧萍、林聃、颜露露、李想、李国强、曾佳丽、谭文娟、杨德援等帮忙在厦门海域进行大型底栖动物采样和分选。
马 骏, 付荣恕, 2010. 大型底栖动物生态学研究进展. 山东农业科学, (2): 78—81
王全超, 李宝泉, 2013. 烟台近海大型底栖动物群落特征. 海洋与湖沼, 44(6): 1667—1680
王金宝, 李新正, 王洪法, 2017. 山东宁津附近海域大型底栖动物群落结构及季节变化. 海洋与湖沼, 48(6): 1371—1382
方少华, 吕小梅, 张跃平, 2002. 厦门国家级自然保护区厦门文昌鱼资源及其保护. 海洋科学, 26(10): 9—12
邓玉娟, 董树刚, 刘晓收, 2016. 福建兴化湾大型底栖动物种类组成和生物多样性. 海洋科学, 40(1): 54—63
吕小梅, 方少华, 1997. 福建沿海文昌鱼的分布. 海洋通报, 16(3): 88—91
吕小梅, 张跃平, 郑承忠等, 2005. 厦门文昌鱼自然保护区的生态环境特点. 海洋科学, 29(10): 27—31
刘 迅, 王 莉, 顾晓英等, 2017. 浙江檀头山岛周围海域夏、秋季大型底栖动物群落结构特征. 海洋与湖沼, 48(3): 553—567
刘好真, 李祥付, 刘 懂等, 2018. 浙江象山东部海域夏季大型底栖动物优势种群和生态位. 海洋与湖沼, 49(4): 873—882
刘春琳, 孙 艺, 2019. 填海造陆对天津大沽口海洋环境影响分析研究. 盐科学与化工, 48(5): 28—32
江锦祥, 陈灿忠, 吴启泉等, 1984. 台湾海峡西部近海底栖生物生态初步研究. 海洋学报, 6(3): 389—398
孙道元, 陈必达, 1988. 台湾海峡(北部)多毛类生态的初步研究. 海洋科学, (2): 43—49
杜飞雁, 张汉华, 李纯厚等, 2008. 大亚湾大型底栖动物种类组成及物种多样性. 中国水产科学, 15(2): 252—259
李亚芳, 杜飞雁, 王亮根等, 2018. 粤西海陵湾养殖区邻近海域大型底栖动物生态学特征. 海洋与湖沼, 49(6): 1294—1307
李荣冠, 江锦祥, 1992. 应用丰度生物量比较法监测海洋污染对底栖生物群落的影响. 海洋学报, 14(1): 108—114
李荣冠, 江锦祥, 蔡尔西等, 1999. 兴化湾大型底栖生物生态研究. 海洋学报, 21(5): 101—109
李晓静, 周政权, 陈琳琳等, 2017. 渤海湾曹妃甸围填海工程对大型底栖动物群落的影响. 海洋与湖沼, 48(3): 617—627
金德祥, 郭仁强, 1953. 厦门的文昌鱼. 动物学报, 5(1): 65—78
周秋麟, 何明海, 邵合道等, 1990. 厦门黄厝海区文昌鱼的分布及其与底质的关系. 台湾海峡, 9(2): 185—189
饶义勇, 张慧萍, 林 聃等, 2015. 厦门海域文昌鱼及其他大型底栖动物资源现状研究. 海洋通报, 34(5): 577—585
翁朝红, 谢仰杰, 肖志群等, 2012. 厦门海域文昌鱼资源及其自然生态环境评价. 集美大学学报(自然科学版), 17(4): 259—264
高 敏, 高 杰, 韩晓庆等, 2017. 近十年来昌黎生态监控区生态环境演变分析. 海洋湖沼通报, (4): 91—97
郭兴然, 刘宪斌, 张青田等, 2019. 河北昌黎海域大型底栖动物群落特征及青岛文昌鱼资源现状. 生态与农村环境学报, 35(8): 1034—1042
唐 盟, 周 进, 2017. 福建三沙湾网箱养殖对多毛纲动物群落结构的影响. 海洋与湖沼, 48(3): 543—552
黄 昆, 陈 岚, 傅婷婷等, 2019. 厦门湾大型底栖动物多样性指数空间分布及其与环境因子的关系. 渔业研究, 41(4): 293—301
黄洪辉, 林 钦, 林燕棠等, 2005. 大亚湾网箱养殖海域大型底栖动物的时空变化. 中国环境科学, 25(4): 412—416
彭松耀, 李新正, 徐 勇等, 2017. 十年间黄海大型底栖动物优势种的变化. 海洋与湖沼, 48(3): 536—542
曾国寿, 何明海, 程兆第, 1996. 厦门黄厝文昌鱼保护区监测与研究. 台湾海峡, 15(2): 174—181
蔡立哲, 2006. 海洋底栖生物生态学和生物多样性研究进展. 厦门大学学报(自然科学版), 45(增刊2): 83—89
Antoniadou C, Krestenitis Y, Chintiroglou C, 2004. Structure of the “Amphioxus sand” community in Thermaikos bay (Eastern Mediterranean). Fresenius Environmental Bulletin, 13(11a): 1122—1128
Cai L Z, Hwang J S, Dahms H W, 2013. Does high organic matter content affect polychaete assemblages in a Shenzhen Bay mudflat, China? Journal of Marine Science and Technology, 21(S1): 274—284
Chen Y, Cheung S G, Shin P K S, 2013. A baseline study of benthic community associated with Amphioxus Sand in subtropical Hong Kong. Marine Pollution Bulletin, 72(1): 274—280
de Moura Barboza C A, Hadlich H L, Sandrini-Neto L, 2013. Is the distribution of the lanceletaffected by sewage discharges? An analysis at multiple scales of variability. Marine Pollution Bulletin, 69(1—2): 178—188
Dubois S, Gelpi Jr C G, Condrey R E, 2009. Diversity and composition of macrobenthic community associated with sandy shoals of the Louisiana continental shelf. Biodiversity and Conservation, 18(14): 3759—3784
Rota E, Perra G, Focardi S, 2009. The European lancelet(Pallas) as an indicator of environmental quality of Tuscan Archipelago (Western Mediterranean Sea). Chemistry and Ecology, 25(1): 61—69
Wang Y, Zhang M M, Zhang D, 2016. The influence of sediment particle size on the properties of adsorbed dissolved organic matter in the Yangtze Estuary and its interactions with As/Sb. Marine Pollution Bulletin, 105(1): 351—358
EFFECTS OF SEDIMENT FINING ON BENTHIC MACROFAUNAL COMMUNITY IN SUBTIDAL AMPHIOXUS HABITATS IN XIAMEN
CHEN Bing-Wen1, CAI Li-Zhe1, 2, RAO Yi-Yong1, LI Wen-Jun1, CHEN Xin-Wei1, FU Su-Jing1, PENG Wen-Qing1, ZHENG Bin1
(1. College of the Environment & Ecology, Xiamen University, Xiamen 361102, China; 2. Key Laboratory of Ministry of Education for Coastal and Wetland Ecosystems, Xiamen University, Xiamen 361102, China)
The effects of sediment fining on the benthic macrofaunal community in Xiamen subtidal zone was studied using the data of benthic macrofauna and grain size analyses for five subtidal areas in Xiamen from 2014 to 2017. The Cluster analysis and one-way ANOVA were conducted. Results show that in those stations with higher clay content, no amphioxus fish but few macrobenthic species were found. Significant differences were seen in some community parameters of benthic macrofauna and dominant species. The contents of clay and silt in Eyu Island and Baicheng subtidal areas were higher than those in Huangcuo, Nanxianshibaxian and Xiaodeng Island subtidal areas in September 2017. The contents of clay and silt in Huangcuo and Nanxianshibaxian subtidal areas in the summer 2016 were significantly higher than those in summer 2014. Significantly positive correlations were found not only between the five community parameters of benthic macrofauna such as the species number, density, biomass, diversity index (), richness index (), and silt plus clay contents, but also between the density and biomass of seven dominant species and silt plus clay contents. No significant correlations were observed between evenness index () and silt plus clay contents, and between density plus biomass of four dominant species and silt plus clay contents. Significantly negative correlations were revealed between the density plus biomass ofand silt plus clay contents. The above results confirmed that when silt or clay contents increased up to 15% or 60%, respectively, it favored the increase in density and biomass of most benthic macrofauna, but resulted in the decrease in the density and biomass of; however, those of,, andwere less affected by sediment fining.
benthic macrofauna; community structure; amphioxus habitats; sediment fining; subtidal areas; Xiamen
* 国家重点研发计划项目, 2016YFC0502904号。陈丙温, 硕士研究生, E-mail: 443614567@qq.com
蔡立哲, 博士生导师, 教授, E-mail: cailizhe@xmu.edu.cn
2019-12-02,
2020-01-28
X17
10.11693/hyhz20191200245
