LNC RNA-MEG 通过 miR-21-5p 影响大鼠脊髓损伤过程中神经细胞凋亡作用探究
2020-05-23唐一钒陈竞轩曹兵苑博石维李凤宁
唐一钒 陈竞轩 曹兵 苑博 石维 李凤宁
脊髓损伤 ( spinal cord injury,SCI ) 主要是由于事故或者暴力打击造成的脊髓组织的直接损伤,同时,肿瘤、病毒感染、血管病变、脊椎病骨质疏松等引发的继发性脊椎骨折、发育相关的疾病等也会造成脊髓损伤[1]。脊髓损伤发生后,会暂时或者永久性地损害脊髓组织的运动、感觉功能,导致患者身心严重受创[2]。目前,全球脊髓损伤发病率大约在 11~60/ 100万[3]。在以往的研究中发现,脊髓损伤的病理过程非常复杂,对患者的伤害主要包括原发性机械损伤和继发性伤害。其中,机械损伤会直接破坏神经元细胞,而继发性损伤则包括血管损伤、水肿、缺血、毒性、电解质的变化、自由基的产生、细胞延迟凋亡等[4-6]。因此,由于脊髓损伤的病理和生物机制的复杂性,导致目前对于脊髓损伤发生后神经细胞的凋亡机制还缺乏系统的了解。
在目前的研究中,研究人员发现,在脊髓损伤诱导的血管和细胞损伤过程中,伴随着大量基因表达的时空变化[7]。非编码 RNA ( non-coding RNA,ncRNA ) 是一类涉及基因表达和分子网络调控的重要分子,对 ncRNA 的研究,可能会为脊髓损伤的治疗提供新的视角[8]。近些年来,随着二代测序的发展和研究人员对 ncRNA 认识的不断加深,长链非编码RNA ( long chain non-coding RNA,LNC RNA ) 的作用日益受到研究人员的重视。一方面,LNC RNA 可以通过干扰转录因子与启动子的结合,诱导组蛋白修饰等方式,抑制蛋白编码基因的表达,从而发挥细胞生物学调控作用[9-12]。另一方面,LNC RNA 在细胞内还存在竞争性内源 RNA ( competing endogenous RNA,ceRNA ) 的调控机制,即通过 RNA-RNA 之间的相互竞争结合来调控基因的表达[13]。
本研究中,笔者重点关注了 ncRNA 在脊髓损伤过程中的作用。首先通过生物信息的方式,对于大鼠中可能与脊髓损伤过程相关的 miRNA 进行了分析,之后通过构建脊髓损伤大鼠模型中,对预测与脊髓损伤有关的 miRNA 进行 qPCR 检测。结果表明,多个基因在脊髓损伤大鼠模型中都有与预测一致的显著变化。之后,进一步发现,抑制 LNC RNA-MEG 可以通过促进 miR-21-5p 的表达,从而抑制脊髓损伤大鼠的神经细胞的凋亡。这表明抑制LNC RNA-MEG,可能是一种潜在的治疗脊髓损伤的方式。
材料与方法
一、大鼠脊髓损伤过程中 miRNA 变化预测
从 NCBI 数据库中分别选取脊髓损伤时间同样为 7天的大鼠及其假实验对照组的基因表达数据( GSE45006) 和 miRNA 表达数据 ( GSE19890)。使用 marray 包和 limma 包进行差异表达分析,使用miRDB、miRTarBase、TargetScan 三个数据库对选出的 miRNA 进行靶基因预测,将其中两个以上数据库都包含的靶基因作为预测结果,与另一套数据中的基因进行匹配。将比对成功的基因及其 miRNA 绘制网络图。
二、大鼠脊髓损伤模型构建
从江苏集萃药康生物科技有限公司购买 7周龄雄性大鼠 10只,根据随机数字分组法,将 10只大鼠分为假手术组和损伤 1周组,每组 5只。通过手术法用重锤暴露并损伤 T10胸段,导致中度脊髓损伤。采用 BBB ( Basso,Beattie 和 Bresnahan ) 方法评估后肢的运动功能变化,伤后 1周收集损伤的脊髓组织样本。
三、大鼠脊髓组织蜡块
大鼠模型构建后 7天,处死大鼠,取出脊髓组织,使用 4% 的中性固定液固定 30min,依次使用50%、70%、80%、95%、100%,100% 的乙醇进行脱水。使用 50% 的二甲苯和 50% 的无水乙醇处理20min,二甲苯分别处理 15min 和 25min。将处理好的组织置于 50% 石蜡+50% 二甲苯中,56℃ 水浴锅保温 40min。再放入石蜡中室温放置 90min。将恒温箱的温度设为 60℃,在包埋盒中放入材料,补足石蜡,倒好后将包埋盒置于冷水盆中,待石蜡完全凝固后取出晾干。
四、大鼠脊髓组织 HE 染色
将蜡块切片,使用二甲苯分别处理 15min 和5min 2次,50% 二甲苯+50%乙醇洗涤 1遍,再依次使用 100%、95%、85%、70%、50% 乙醇和清水洗涤,将切片放入苏木精中染色约 20min,用自来水冲洗约 15min,使切片变蓝。将切片放入 1% 盐酸乙醇溶液中褪色,在放入清水中冲洗。将切片放入 50%、70%、80% 乙醇中各 5min,然后使用0.5% 的伊红乙醇溶液对比染色 3min。放入 95% 的乙醇中洗去多余的红色,再放入无水乙醇中 5min。封片,显微镜下观察拍照。
五、大鼠脊髓组织 Tunel 染色
室温下将切片放入二甲苯中浸泡 5min,重复1次。用 100% 乙醇浸泡切片 5min,重复 1次。用90%、80%、70% 乙醇各浸洗 1次,每次 3min。用 PBS 洗涤切片 1次。每个样本滴加 100μl 浓度为20μg / ml 的 Proteinase K 溶液,室温孵育 20min。PBS 溶液润洗样本 2~3次。每个样本滴加 100μl 1×Equilibration Buffer,室温孵育 30min。扔去1×Equilibration Buffer,然后在 5cm2面积的细胞上加入 50μl TdT 孵育缓冲液。将载玻片置于湿盒内,在 37℃ 避光孵育 60min。PBS 溶液中室温孵育5min,再用 PBS 重复清洗 2次。使用 PI 溶液 ( 1μg /ml ) 染色缸避光染色,室温放置 5min。去离子水洗涤,室温放置 5min。重复 2次,总共洗 3次。立即在荧光显微镜下分析样本。
六、大鼠 BBB 评分标准
0分:无可见后肢运动;1分:一个或两个关节轻微运动,通常为髋 / 膝关节;2分:一个关节广泛活动或一个关节广泛活动且有另一个关节轻微活动;3分:两个关节广泛活动;4分:后肢全部三个关节可轻微活动;5分:两个关节轻微活动,第三个关节可广泛活动;6分:连个关节广泛活动,第三个关节可轻微活动;7分:后肢全部三个关节可广泛活动;8分:非承重情况下可以爪掌面着地;9分:间或爪掌面承重支撑或爪背面承重移动,无爪掌面支撑移动;10分:偶见爪掌面承重移动;无前后肢协调动作;11分:可较多的见爪掌面承重移动,但无前后肢协调动作;12分:可较多的见到掌面承重移动,偶见前后肢协调动作;13分:常见掌面承重移动,可常见前后肢协调动作;14分:有持续性掌面承重移动和前后肢协调动作;或出现常见的掌面移动,持续型前后肢协调动作,偶有爪背侧移动;15分:持续性掌面移动和持续性前后肢协调动作,前肢前进过程中无或偶有抓地;初接触时主动爪位置与身体平行;16分:步态中可见持续性掌面移动和持续性前后肢协调动作,前肢前进过程中常见爪抓地;初接触时主动爪位置与身体平行,负重转移后旋转;17分:步态中可见持续性掌面移动和持续性前后肢协调动作,前肢前进过程中常见爪抓地;初接触时和负重转移后主动爪位置均与身体平行;18分:步态中可见持续性掌面移动和持续性前后肢协调动作,前肢前进过程中可持续性爪抓地;初接触时主动爪位置均与身体平行,负重转移后旋转;19分:步态中可见持续性掌面移动和持续性前后肢协调动作,前肢前进过程中可持续性爪抓地;初接触时和负重转移后主动爪位置均与身体平行。尾巴有时或总是下垂;20分:持续性掌面移动,持续性协调步态,足趾持续抓地,初接触时和负重转移后主动爪位置均与身体平行,躯干不稳定,尾巴持续翘起;21分:持续性掌面移动,持续性协调步态,足趾持续抓地,活动过程中主动爪位置始终与身体平行,躯干持续稳定,尾巴持续翘起。
七、统计学处理
采用 GraphPad 软件对实验数据进行统计分析,采用±s的进行作图分析。使用T检验进行统计学显著性分析。
结 果
一、大鼠脊髓损伤过程中基因表达和 miRNA 变化的生物分析
从 NCBI 数据库中共获得 2189个上调基因和2656个下调基因 ( 图 1a ),与此同时,笔者还使用marray 包和 limma 包进行差异表达分析,得到了 20个上调的 miRNA 和 30个下调的 miRNA ( 图 1b )。在这之后,结合差异表达基因和差异表达 miRNA 的数据,在 miRDB、miRTarBase 和 TargetScan 三个数据库中进行筛选比对,笔者发现,miRNA 和靶标基因变化一致的 miRNA 17个,而下调的 miRNA 则有29个 ( 图 1c、d )。上述分析结果为后续研究提供了初步的候选 miRNA。

图1 大鼠脊髓损伤过程中 miRNA 变化的预测分析。通过从 NCBΙ 数据库获得损伤时间同样为 7天的大鼠及其假实验对照组的基因表达数据 ( GSE45006) 和 miRNA 表达数据 ( GSE19890) a~b:使用 limma 包 ( a ),marray 包和 limma 包 ( b ) 分析过程中表达差异的热点图 ( 纵坐标为 miRNAS )。红色代表上调,绿色代表下调;c~d:在 miRDB、miRTarBase 和 TargetScan 三个数据库中进行筛选比对,获得上调和下调的候选 miRNA 和其对应的潜在的靶标基因Fig.1Predictive analysis of miRNA changes during SCΙ in rats. Gene expression data ( GSE45006) and miRNA expression data ( GSE19890)of rats and their control group were also obtained from the NCBΙ database for 7days a - b: Heat maps showing the differences in expression during the analysis using the limma package ( a ), the marl package and the limma package ( b, The vertical coordinate represents the miRNAS ).Red represented upwards and green represented downwards; c - d: Screening and comparison among three databases of miRDB, miRTarBase and TargetScan to obtain up- and down-regulated candidate miRNAs and their corresponding potential target genes
二、大鼠脊髓损伤模型的构建
使用材料与方法中描述的方法构建大鼠脊髓损伤模型后,笔者首先对脊髓损伤大鼠进行 BBB 评分,笔者发现,脊髓损伤后,大鼠 BBB 评分迅速下降,但是随着时间的迁移,BBB 评分上升,这表明大鼠的脊髓神经元得到了恢复 ( 图 2a )。HE 染色结果表明,脊髓损伤大鼠相比较于对照组大鼠脊髓组织中,新生组织较多,表明脊髓损伤模型大鼠正处于受损后的恢复期。笔者进一步通过 Tunel 染色实验,发现脊髓损伤大鼠中凋亡细胞数量多于对照组大鼠,这表明脊髓损伤大鼠神经元细胞收到损害( 图 2b、c )。这些结果都表明,笔者成功建立了一个脊髓损伤大鼠模型。
三、脊髓损伤大鼠模型中候选 miRNA 的 qPCR验证
脊髓损伤大鼠模型建立后 7天,处死脊髓损伤大鼠和对照组大鼠后,抽提大鼠脊髓组织的总RNA。根据先前 NCBI 数据库分析的候选 miRNA,分别选择 10个上调和下调的候选 miRNA 通过 qPCR进行单点验证。qPCR 结果表明,在脊髓损伤大鼠中,10个候选的上调的 miRNA 中,miR-124-3p,miR184,miR215和 miR-300-3的表达水平显著增加,而 10个下调的 miRNA 中,miR-211-3p,miR-129-5p 和 miR-21-5p 出现显著下降 ( 图 3a、b )。

图2 大鼠脊髓损伤模型构建。采用手术暴露并用重锤损伤 T10胸段的建模方法建立大鼠脊髓损伤模型 a:大鼠模型建立前后,使用 BBB 评分评估大鼠的运动能力。1号为未处理大鼠,2~5号为模拟假手术对照组,6~10号为脊髓损伤模型组;b:大鼠脊髓损伤模型建立后 7天,大鼠脊髓组织 HE 染色 ( × 40);c:大鼠脊髓损伤模型建立 7天后,大鼠脊髓组织的 Tunel 染色( × 100)。蓝色:DAPΙ;红色:dUTP-PEFig.2Construction of rat SCΙ model. The rat SCΙ model was established by surgical exposure and T10 chest damage with heavy hammer a: BBB score was used to evaluate the exercise ability of the rat before and after the rat model establishment. Rat 1: An untreated rat. Rat 2- 5: Simulated control group. Rat 6- 10: SCΙ model group;b: HE staining of rat spinal cord tissues 7days after the SCΙ model establishment ( × 40), c: Tunel staining of rat spinal cord tissues 7days after the SCΙ model establishment ( × 100). Blue: DAPΙ. Red:dUTP-PE.
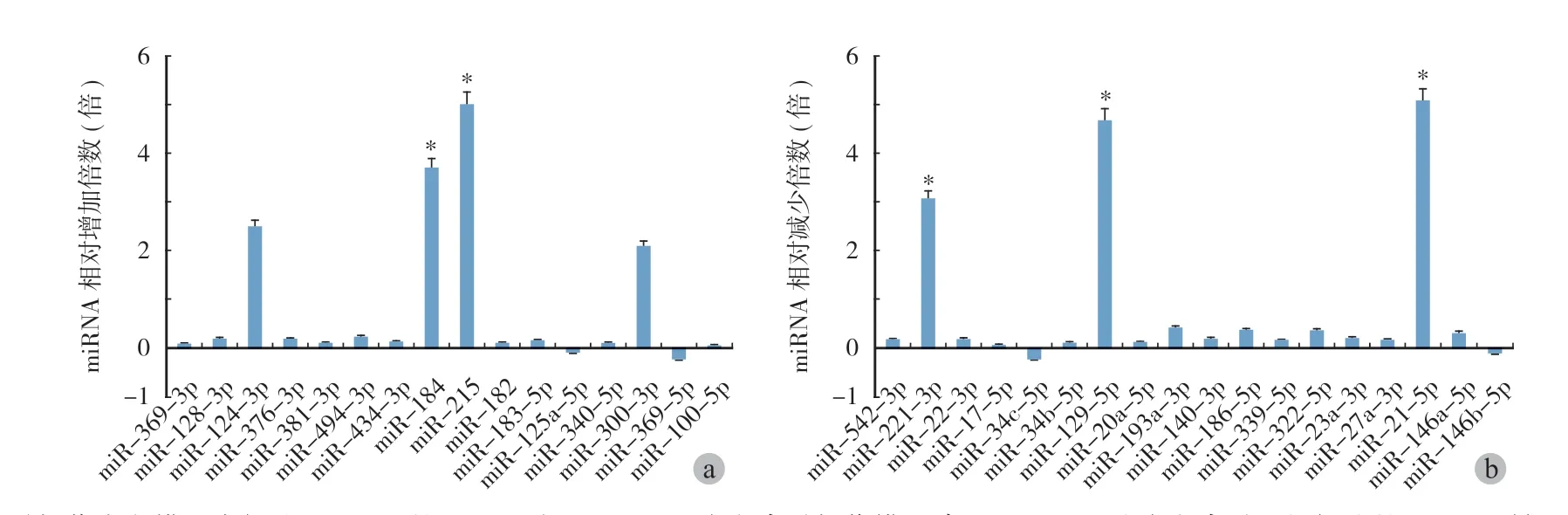
图3 脊髓损伤大鼠模型中候选 miRNA 的 qPCR 验证 a~b:大鼠脊髓损伤模型建立后 7天,取大鼠脊髓组织提取的 RNA 反转录样本作为模板。使用特异性引物进行 qPCR 验证候选 miRNA 表达的变化。预测上调的 miRNA ( a ) 和下调的 miRNA ( b ) 的相对变化倍数是针对 U6的。每个数据都是平均值 ± 标准差,每个实验都是独立重复 3次 (P < 0.05)Fig.3qPCR validation of candidate miRNAs in SCΙ rat models a - b: RNA reverse transcription samples extracted from rat spinal cord tissues were used as templates 7days after the rat SCΙ model establishment. qPCR was performed using specific primers to verify changes in candidate miRNA expression. The predicted relative fold changes of up-regulated miRNA ( a ) and down-regulated miRNA ( b ) were targeted to U6. Each data point represented the mean ± SD of three independent experiments and was analyzed with a T-test. *P < 0.05
四、LNC RNA-MEG 在脊髓损伤大鼠中表达量的变化
在确定脊髓损伤过程中显著变化的 miRNA 后,通过生物信息预测发现,LNC RNA-MEG 能够和miR-21-5p 发生竞争结合,因此推测,在脊髓损伤损伤过程中,LNC RNA-MEG 可能表达量上调,从而抑制 miR-21-5p,进而抑制脊髓损伤大鼠神经细胞的恢复。为了验证假设,首先在脊髓损伤大鼠模型和对照组大鼠中,检测了 LNC RNA-MEG 表达的变化,结果表明,在脊髓损伤模型大鼠中,LNC RNAMEG 的表达量是显著高于对照组大鼠的 ( 图 4)。这表明 LNC RNA-MEG 很可能在大鼠脊髓损伤过程中发挥重要的作用。
五、抑制 LNC RNA-MEG 可以抑制脊髓损伤大鼠神经细胞的凋亡
本研究中,在脊髓损伤大鼠模型中,脊髓损伤过程中 miR-21-5p 的表达出现了显著下降,而脊髓损伤大鼠中 LNC RNA-MEG 的表达则有显著增加。并且生物信息学也预测,LNC RNA-MEG 能够和miR-21-5p 发生竞争结合。因此笔者推测,通过抑制 LNC RNA-MEG,可以促进 miR-21-5p 的表达,从而抑制脊髓损伤大鼠的神经细胞凋亡。

图4 脊髓损伤大鼠模型中 LNC RNA-MEG 的表达变化。大鼠脊髓损伤模型建立后 7天,取大鼠脊髓组织提取的 RNA 反转录样本作为模板。使用特异性引物进行 qPCR 验证 LNC RNA-MEG 表达的变化。相对变化倍数是针对 U6的。每个数据都是平均值 ±标准差,每个实验都是独立重复 3次 ( **P < 0.01)Fig.4Changes of LNC RNA-MEG expression in SCΙ rat models.RNA reverse transcription samples extracted from rat spinal cord tissues were used as templates 7days after the rat SCΙ model establishment. qPCR was performed using specific primers to verify changes in LNC RNA-MEG expression. The predicted relative fold changes were targeted to U6. Each data point represented the mean ±SD of three independent experiments and was analyzed with a T-test.**P < 0.01

图5 敲低 LNC RNA-MEG 对脊髓损伤大鼠模型的作用 a:筛选有效抑制 LNC RNAMEG 的 siRNA。在 VSC4.1细胞中,瞬时转染靶向 LNC RNA-MEG 的 siRNA,通过 qPCR检测 LNC RNA-MEG 的相对敲低水平。相对变化是针对对照组的归一化的。每个数据都是,每个实验都是独立重复 3次;b:shRNALNC RNA-MEG 对于脊髓损伤大鼠模型的BBB 评分的影响。使用重锤暴露并损伤 T10 胸段的建模方法建立大鼠脊髓损伤模型。并同时原位注射 shRNA-LNC RNA-MEG 的慢病毒。在不同时项点采用 BBB 进行评分。1~5号为对照组模型大鼠,6~10号为实验敲低组大鼠;c:大鼠模型对照组和实验组中 miR-21-5p 和 LNC RNA-MEG 的表达变化。在大鼠建模后 7天,处死大鼠,取大鼠脊髓组织的RNA,反转录通过 qPCR 检测 LNC RNA-MEG和 miR-21-5p 的相对表达水平。相对变化倍数是针对 U6的。每个数据都是,每个实验都是独立重复 3次;d:大鼠模型对照组和实验组中凋亡相关蛋白 Caspased 3和 Bcl2的表达变化。通过大鼠脊髓组织提取总蛋白,使用Western blot 检测凋亡相关蛋白 Caspased 3和 Bcl2的表达变化,内参对照是 GAPDH ( *P < 0.01,**P < 0.01)Fig.5Knockdown of LNC RNA-MEG on SCΙ rat models a: Screen for siRNAs that effectively inhibited LNC RNA-MEG. Ιn VSC4.1cells,LNC RNA-MEG-targeting siRNA was transiently transfected, and the relative knockdown of LNC RNA-MEG was detected by qPCR. The relative changes were normalized for the control group. Each data point represented the mean ± SD of three independent experiments and was analyzed with a T-test. *P < 0.05; b: Effects of shRNA-LNC RNA-MEG on BBB score of SCΙ rat model. The rat SCΙ model was established by surgical exposure and T10 chest damage with heavy hammer. shRNA-LNC RNA-MEG was injected in situ at the same time. BBB evaluation was performed at different times. Rat 1- 5: Control group. Rat 6- 10: Experimental knock-down group; c: The expression changes of miR-21-5p and LNC RNAMEG in the rat model control group and experiment group. Rats were sacrificed, and the RNA from rat spinal cord tissues was taken 7days after the rat model establishment. The relative expression levels of LNC RNA-MEG and miR-21were detected by reverse transcription and qPCR. Each data point represented the mean ± SD of three independent experiments and was analyzed with a T-test. *P < 0.05; d: Changes in expressions of apoptosis-related proteins Caspased 3and Bcl2in the rat model control group and experiment group. Total protein was extracted from rat spinal cord tissues. Western blot was used to detect the expression of apoptosis-related proteins Caspased 3and Bcl2. The control was GAPDH ( *P < 0.01,**P < 0.01)
为此,笔者首先合成了 3对针对 LNC RNAMEG 的 siRNA,并瞬时转染 VSC4.1细胞,通过qPCR 检测 LNC RNA-MEG 的表达量变化。结果表明,siRNA-2对于 LNC RNA-MEG 的抑制作用最为显著 ( 图 5a )。因此,将 siRNA-2的序列构建到了慢病毒 shRNA-pLKO.1载体上进行慢病毒包装。之后,笔者在构建的脊髓损伤大鼠模型中,在建模后1天原位注射 shRNA-LNC RNA-MEG 慢病毒。通过BBB 评分评估,笔者发现,慢病毒处理组和未处理组大鼠的 BBB 评分没有显著差异 ( 图 5b )。于是,笔者在建模后第 7天,处死两组大鼠,抽提脊髓组织中的总 RNA 和蛋白,分别进行 qPCR 和 Western blot检测。结果表明,在原位注射 shRNA-LNC RNAMEG 慢病毒后,注射慢病毒的大鼠脊髓组织中 LNC RNA-MEG 的表达量显著下降,而 miR-21-5p 的表达量出现了显著上升 ( 图 5c )。笔者通过 Western blot检测凋亡相关蛋白 Cleaved caspased3和 Bcl2,结果发现,在慢病毒注射组中,凋亡相关蛋白 Cleaved caspased3和 Bcl2的表达量显著下降 ( 图 5d ),这表明通过抑制 LNC RNA-MEG,可以抑制凋亡相关蛋白的表达,因此可能抑制了神经细胞的凋亡。
讨 论
近些年来,随着脊髓损伤动物模型不断完善,对于脊髓损伤的研究取得了长足的进步。研究人员对于脊髓损伤过程中神经细胞凋亡、增殖等分子机制已经做了深入的研究[9-11]。尽管如此,基于脊髓损伤的分子机制提出的各种治疗策略,依然不能够很好的解决脊髓损伤的治疗困境[14]。因此,有必要对于脊髓损伤的分子机制进行进一步的研究。
随着二代测序和非编码 RNA 研究的不断进展,对于脊髓损伤过程中非编码 RNA 的作用的研究越来越多[9,15-16]。特别是对于 miRNA,已经有多个课题组报道了多个 miRNA 与脊髓损伤具有相关性。其中,研究人员发现,miR-21-5p 能够参与神经元细胞损伤后的修复过程[17];miR-17-5p 也是脊髓损伤后的重要的调节因子,其能够通过调节 P21或者RB1,从而调节星形胶质细胞的激活和增殖[18];在小鼠 P19细胞中,过表达 miR-124能够促进神经轴突生长,显著增加神经元细胞数量,从而有利于脊髓损伤小鼠的功能性恢复[19-20]。除了 miRNA,LNC RNA 也是非编码 RNA 中重要的成员。在之前的研究中,研究员发现,在外周神经损伤过程中,LNC RNA uc.217能促进背神经元细胞的生长和神经网络的恢复[21]。但是在脊髓损伤中,目前尚未发现和脊髓损伤过程有关的 LNC RNA,因此,在脊髓损伤过程中,是否有 LNC RNA 参与?以及 LNC RNA 参与脊髓损伤的作用和机制还缺乏研究。
在本研究中,笔者首先通过生物信息学从 NCBI数据库中分析脊髓损伤大鼠模型中,基因和 miRNA表达变化情况,并将两者结合,预测了可能与脊髓损伤作用过程相关的 miRNA。对于预测的候选miRNA,笔者通过构建脊髓损伤大鼠模型进行了单点验证。结果表明,笔者预测的部分候选 miRNA 在大鼠脊髓损伤模型中具有与预测一致的变化,这与其他研究人员之前的报道是一致的[22-25]。对于变化最为显著的 miR-21-5p,笔者进一步分析发现,其与 LNC RNA-MEG 具有竞争作用,因此推测,在脊髓损伤过程中,LNC RNA-MEG 可能通过抑制 miR-21-5p,从而促进大鼠神经细胞的增殖。因此,为了验证假设,笔者首先筛选了能够抑制 LNC RNAMEG 的 siRNA,并使用 shRNA 慢病毒在脊髓损伤大鼠模型中探究抑制 LNC RNA-MEG 对于脊髓损伤大鼠的作用。最终的结果表明,使用 shRNA-LNC RNA-MEG 慢病毒处理脊髓损伤大鼠后,大鼠中LNC RNA-MEG 的表达显著下降,而 miR-21-5p 的表达出现了显著上升,与此同时,在脊髓损伤大鼠脊髓神经组织中,凋亡相关的蛋白 Cleaved caspased3和 Bcl2表达量显著下降,表明抑制 LNC RNAMEG,可以抑制凋亡相关蛋白的表达,从而抑制了神经细胞的凋亡。
本研究通过生物信息学和实验结合,在脊髓损伤大鼠模型中,首次确证了 LNC RNA 参与了大鼠脊髓损伤的过程,并且在后续的动物实验中,也发现,LNC RNA-MEG 还可能是一个潜在的治疗脊髓损伤的靶标。最为重要的是,本研究可能打开了LNC RNA 与脊髓损伤研究的大门,并提示其他研究员探究更多与脊髓损伤有关的 LNC RNA,进一步完善脊髓损伤背后的分子机制。尽管如此,本研究还有进一步完善的空间,比如除了抑制神经细胞凋亡。LNC RNA-MEG 对于神经细胞的增殖是否有促进作用?其次,LNC RNA-MEG 抑制 miR-21-5p,下游的作用基因是什么,下游的基因又是如何抑制神经细胞凋亡的?LNC RNA-MEG 在大鼠脊髓损伤过程中具有的作用,在人类细胞中是否也具有类似的作用?这些问题,希望在未来的研究中进行进一步的验证和解答。
总之,本研究首次发现 LNC RNA-MEG 能够通过 miR-21-5p 影响大鼠脊髓损伤过程中神经细胞的凋亡作用。
