2000—2019年广西狂犬病病毒流行株的基因组遗传稳定性分析
2020-05-11覃晴梁星雪曹晗韦显凯梁晶晶李晓宁罗廷荣
覃晴 梁星雪 曹晗 韦显凯 梁晶晶 李晓宁 罗廷荣


摘要:【目的】揭示广西狂犬病病毒(RABV)的流行变异规律及进化特征,为广西狂犬病防控提供科學依据。【方法】对2018年从患狂犬病家犬脑组织分离获得的2株RABV野毒株(GXYZ2018和GXSL2018)进行全基因组扩增与测序,测序结果采用SeqMan进行拼接以获得全基因组序列,并与近20年来的RABV广西流行株进行系统地遗传变异分析。【结果】GXSL2018株和GXYZ2018株的全基因组序列长度均为11923 bp,不同RABV广西流行株间的核苷酸序列同源性在87.1%~99.4%,而2株毒株间的同源性为99.1%;与其他RABV广西流行株相比,GXSL2018株和GXYZ2018株的3'-UTR为69 bp,且在第20位缺失一个核苷酸。基于N基因和G基因核苷酸序列同源性构建的遗传进化树均显示GXSL2018株和GXYZ2018株同属于Group II型毒株,且2株毒株N蛋白上的B细胞表面抗原表位(第358~367位氨基酸残基)、Th细胞表位(第404~418位氨基酸残基)和RNA结合域(第298~352位氨基酸残基),以及G蛋白上与病毒毒力密切关联的第333、336、339和357位氨基酸残基及中和抗体相关表位和受体结合位点均高度保守。【结论】GXYZ2018株和GXSL2018株同属于Group II型毒株,与近20年来的RABV广西流行株高度相似,病毒蛋白主要功能区的氨基酸序列高度保守,尚未发生变异,全基因组遗传特性稳定。
关键词: 狂犬病病毒(RABV);基因组;遗传稳定性;Group II型毒株
0 引言
【研究意义】狂犬病是由狂犬病病毒(Rabies virus,RABV)引起的一种人畜共患传染病,具有高致死率。RABV主要侵害动物和人类的神经系统,通常是经受感染的动物咬伤传播(陈芳芳,2013;徐素萍等,2018)。广西是我国狂犬病主要疫区之一,每年因狂犬病死亡的人数居全国首位,且主要发生在乡村地区。虽然经过近年来的病原监测及家犬免疫接种,狂犬病疫情得到明显控制,但其引起的公共卫生问题仍不容小觑。【前人研究进展】RABV是一种高度噬神经性的单股负链RNA病毒,隶属于弹状病毒科(Rhabdoviridae)狂犬病病毒属(Lyssavirus)(周松峰等,2011;谭业平等,2018),其病毒蛋白包括核蛋白(N)、非酶促聚合酶辅因子磷蛋白(P)、基质蛋白(M)、糖蛋白(G)和病毒RNA聚合酶蛋白(L)(Liu et al.,2017)。由RNA、N蛋白、P蛋白及L蛋白共同组成的核糖核蛋白复合体(Ribonucleoprotein,RNP)是RABV转录和复制的核心组分(Jackson,2011)。P蛋白对机体免疫系统具有拮抗作用,Vidy等(2007)研究发现P蛋白能拮抗Ⅰ型干扰素的转录,通过阻断信号分子STAT1与干扰素基因启动子结合的方式发挥作用。L蛋白是一种多功能酶,既具备RNA依赖的RNA聚合酶活性(RNA dependent RNA polymerase,RDRP),又具有mRNA加帽酶作用,已有研究发现突变L蛋白携带的关键甲基化位点,能显著降低RABV对小鼠的致病性(Tian et al.,2015;王朝等,2017)。在RABV复制过程中,M蛋白可通过招募RNP使其固定于细胞膜,而有利于病毒出芽(彭娇娇等,2016)。G蛋白是RABV与宿主细胞结合的配体,对病毒毒力及致病性起关键作用(杨健等,2012),已有研究证实将Flury株G蛋白的第333位氨基酸残基突变为精氨酸(Arg)后,RABV可致死成年小鼠,即弱毒致病性发生改变(Tao et al.,2010;Zhang et al.,2013)。Yamada等(2014)研究发现,多数RABV流行株的G蛋白存在第37位和第319位2个糖基化位点,当G蛋白发生糖基化时也会影响病毒毒力。家犬在传播RABV的过程中起重要作用,广西农村散养家犬数量庞大,接种疫苗率低是狂犬病高发的主要原因。1999—2001年广西的临床健康犬RABV阳性率为1.10%(罗廷荣等,2007),2002—2005年的RABV阳性率为3.25%~4.39%,2006—2011年的RABV阳性率为3.72%~1.26%,2011—2012年的RABV阳性率为0.20%,2013—2018年的RABV阳性率为0。随着家犬免疫接种率的提高,广西狂犬病致死亡人数呈逐年下降趋势,由2004年的602人降至2016年的57人,说明人类的狂犬病发病率与家犬带毒率呈正相关(罗廷荣等,2017)。免疫接种是降低临床健康犬带毒的重要手段,但免疫抗体同时给RABV生存带来压力,诱发其产生变异,因此监测病毒遗传稳定性及分析其变异规律,对有效防控狂犬病具有重要意义。【本研究切入点】已有相关研究对不同时期广西RABV的遗传特性进行分析,并证实RABV广西流行株绝大部分属于Group I和Group II型毒株,Group III型毒株较少(Tang et al.,2013;Wei et al.,2018),但鲜见针对广西近20年来的RABV流行株进行综合评估分析。【拟解决的关键问题】对本课题组2018年从患狂犬病家犬脑组织分离获得的2株RABV野毒株进行全基因组测序,并与近20年来的RABV广西流行株进行遗传变异分析,旨在揭示广西RABV的流行变异规律及进化特征,为广西狂犬病防控提供科学依据。
1 材料与方法
1. 1 试验材料
2018年从患狂犬病家犬脑组织分离获得2株RABV野毒株(GXSL2018和GXYZ2018),其中,GXSL2018株分离自广西南宁市上林县咬人的患狂犬病家犬脑组织,GXYZ2018株分离自广西河池市宜州区咬牛的患狂犬病家犬脑组织。组织RNA提取试剂盒、pMD18-T载体、PrimeSTAR Max和M-MLV Reverse Transcriptase购自TaKaRa公司;DNA胶回收试剂盒、2×Taq PCR MasterMix、dNTPs和DNA Marker购自天根生化科技(北京)有限公司;胰蛋白胨和酵母提取物购自Oxid公司;LA固体培养基、LB液体培养基和大肠杆菌TOP10感受态细胞由亚热带农业生物资源保护与利用国家重点实验室自配或保存提供。
1. 2 引物设计与合成
参照RABV广西毒株GX074全基因组序列(GenBank登录号MG201923.1),采用Primer 5.0进行RABV全基因组合成引物(表1)设计,并委托深圳华大基因股份有限公司合成。
1. 3 RT-PCR扩增
参照组织RNA提取试剂盒说明提取GXSL2018和GXYZ2018株的总RNA,并反转录合成cDNA。RT反应体系25.0 μL:5×Buffer 5.0 μL,dNTPs 2.0 μL,RNasin 0.5 μL,M-MLV Reverse Transcriptase 0.5 μL,上游引物1.0 μL,RNA模板16.0 μL。反转录程序:42 ℃ 1 h,90 ℃ 5 min。以反转录合成的cDNA为模板进行PCR扩增,反应体系26.0 μL:PrimeSTAR Max 12.5 μL,上、下游引物各1.0 μL,cDNA模板3.0 μL,ddH2O 8.5 μL。扩增程序:98 ℃预变性3 min;98 ℃ 10 s,退火温度(表1)退火15 s,延伸温度72 ℃(延伸速率1 kb/min),据此计算延伸时间,进行35个循环;72 ℃延伸10 min。PCR扩增产物经1.0%琼脂糖凝胶电泳检测后,对目的条带进行DNA胶回收纯化。
1. 4 全基因组序列比对分析
将目的片段与pMD18-T载体在4 ℃下连接过夜,然后转化感受态细胞。挑取疑似阳性单克隆菌落进行扩大培养,经菌液PCR鉴定呈阳性的菌液送至深圳华大基因股份有限公司测序。测序结果采用SeqMan完成序列拼接,并与近20年来的RABV广西流行株(表2)全基因组序列进行比对分析。
2 结果与分析
2. 1 RABV全基因组序列比对分析结果
利用12对引物分别对GXSL2018和GXYZ2018株进行全基因组分段扩增,获得相应目的片段后采用SeqMan进行序列拼接,结果得到2株RABV野毒株的全基因组序列,其基因组全长均为11923 bp,GenBank登录号分别为MN186249和MN186250。
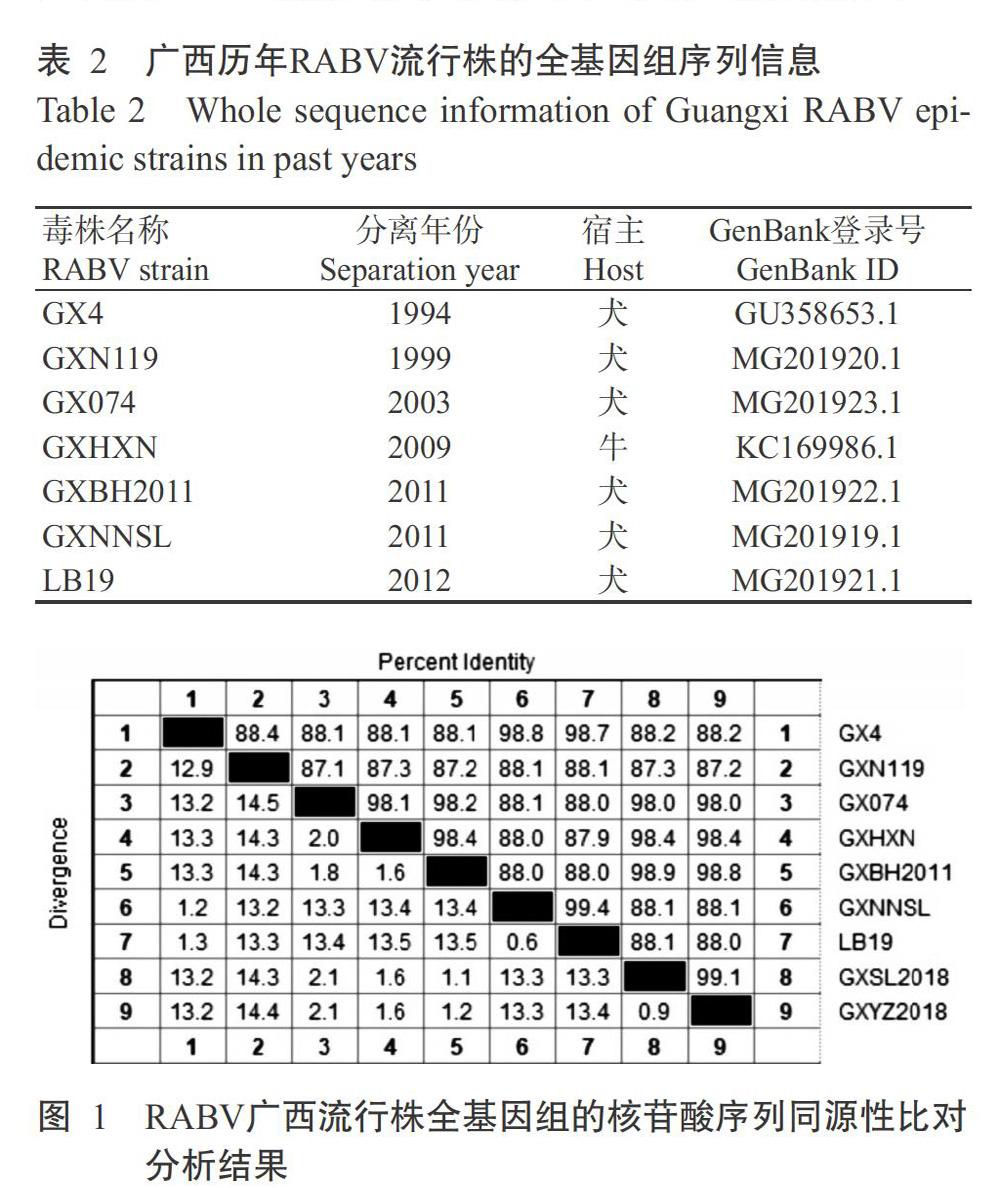
从NCBI下载2000—2019年RABV广西流行株的全基因组序列,运用MegAlign进行同源性比对分析,结果发现不同RABV广西流行株间的核苷酸序列同源性在87.1%~99.1%,其中GXSL2018株和GXYZ2018株的同源性为99.4%(图1)。9株RABV广西流行株的基因组序列全长为11921~11924 bp(表3),其主要区别在于P-M间隔区和5'-UTR,P-M间隔区多为87 bp(6株),只有3株毒株为86 bp,5'-UTR多为130或131 bp,但LB19株的5'-UTR为129 bp。2018年分离获得的GXSL2018株和GXYZ2018株全基因组序列均为11923 bp,与其他RABV广西流行株相比,其主要差异是这2株毒株的3'-UTR为69 bp,且在第20位缺失一个核苷酸(图2)。综上所述,近20年来广西的RABV流行株非常保守,其遗传特性稳定。
2. 2 RABV的N基因序列比对分析结果
从NCBI下载近20年来的RABV广西流行株及国内其他省份具有代表性毒株的N基因序列,进行遗传进化树分析。结果表明,2018年分离获得的GXSL2018株和GXYZ2018株同属于Group II型毒株(图3)。此外,在Group II内,所有RABV广西流行株聚类在同一个分支(红色实线圈出的毒株);在GroupⅠ内,所有RABV广西流行株也聚类在同一分支上。GXYZ2018株和GXSL2018株虽然分离自广西的不同地区,两地相距500 km,但2株毒株的N基因核苷酸序列同源性高达99.6%。与其他省份的RABV流行株相比,RABV广西流行株N蛋白上的RNA结合域(第298~352位氨基酸残基)、B细胞表面抗原表位(第358~367位氨基酸残基)和抗原位点(第359~366位氨基酸残基和第375~383位氨基酸残基)及Th细胞表位(第404~418位氨基酸残基)均高度保守,因此在遗传进化树上RABV广西流行毒株紧密聚在一起。
2. 3 RABV的G基因序列比对分析结果
基于G基因序列的遗传进化分析结果也表明,GXSL2018株和GXYZ2018株同属于Group II型毒株(图4)。从遗传进化树可直观看出,近年来RABV广西流行株主要为Group I型和Group II型毒株,且同一组别内的RABV广西流行株均聚类在同一分支上。成熟的G蛋白从结构上可分为3个部分:膜外区(第1~439位氨基酸残基),该区携带有RABV的主要抗原决定簇位点,具有强大的免疫原性和抗原性,在受体识别和膜融合过程中发挥重要作用;跨膜区(第440~461位氨基酸残基),该区参与G蛋白在病毒脂质双分子层膜上的固定;膜内区(第462~505位氨基酸残基),该区位于病毒膜内,与M蛋白和N蛋白相互作用有关(汪孟航等,2016)。通过氨基酸序列比对分析发现,RABV广西流行株在以上重要抗原位点均高度保守,发生突变的氨基酸位点是否与病毒毒力及抗原性相关,目前尚未得到验证。
3 讨论
病毒变异是病毒病防控的主要难题。本课题组2018年从患狂犬病家犬脑组织中分离获得2株RABV野毒株(GXSL2018和GXYZ2018),在全基因組测序的基础上,与近20年来的RABV广西流行株进行系统地遗传变异分析,结果发现GXSL2018株和GXYZ2018株的基因组全长均为11923 bp,且2株毒株的开放阅读框(ORF)长度与之前分离获得的RABV野毒株及固定毒株一致,仅少数碱基缺失或插入出现在间隔区。何晓霞(2013)研究发现,RABV的3'-UTR较保守,通常为70 bp;但本研究发现GXSL2018株和GXYZ2018株的3'-UTR在第20位点缺失一个核苷酸。病毒的3'-UTR和5'-UTR通常被认为与病毒的转录和复制有关,水疱性口炎病毒(VSV)3'-UTR前24个核苷酸缺失会极大降低其转录复制水平(Li and Pattnaik,1999),故推测GXSL2018株和GXYZ2018株的3'-UTR缺失一个核苷酸也会影响其转录复制水平。
开展RABV不同功能基因的遗传进化分析有利于判断其流行趋势。N基因高度保守和高效表达的特性,被视为RABV基因分型的重要判断指标(路静等,2015)。RABV广西流行株主要属于Group I和Group II型毒株(熊毅等,2008),本课题组2018年分离获得的GXYZ2018株和GXSL2018株同属于Group II型毒株,其N蛋白上的B细胞表面抗原表位(第358~367位氨基酸残基)、Th细胞表位(第404~418位氨基酸残基)和RNA结合域(第298~352位氨基酸残基)均高度保守,与Kouznetzoff等(1998)、Faber等(2002)分离报道的毒株高度一致,而其他发生变异的少数氨基酸残基,与N蛋白功能位点无关。G蛋白负责诱导机体产生中和抗体,也是决定RABV致病性的重要蛋白,位于病毒膜表面,与细胞受体结合(Faber et al.,2002)。本研究对近20年来的RABV广西流行株进行比对分析,结果发现G蛋白上与病毒毒力密切关联的第333、336、339和357位氨基酸残基及中和抗体相关表位和受体结合位点的氨基酸残基均未发生变异,高度保守。此外,P蛋白和M蛋白功能位点的氨基酸残基也非常保守,均未发生变异。综上所述,GXYZ2018株和GXSL2018株与近20年来的RABV广西流行株高度相似,病毒蛋白主要功能区的氨基酸序列高度保守,尚未发生变异,全基因组遗传特性稳定。
4 结论
GXYZ2018株和GXSL2018株同属于Group II型毒株,与近20年来的RABV广西流行株高度相似,病毒蛋白主要功能区的氨基酸序列高度保守,尚未发生变异,全基因组遗传特性稳定。
参考文献:
陈芳芳. 2013. 2006年—2010年广西狂犬病的流行病学调查及综合防控[D]. 南宁:广西大学. [Chen F F. 2013. Epidemiology investigation and prevention of rabies in Guangxi from 2006 to 2010[D]. Nanning:Guangxi University.]
何晓霞. 2013. 2011—2012年广西犬脑组织狂犬病病毒检测及其全基因组序列分析[D]. 南宁:广西大学. [He X X. 2013. The detection of rabies virus of dog brain in Guangxi from 2011 to 2012 and analysis of complete genome sequences[D]. Nanning:Guangxi University.]
路静,付玉荣,伊正君. 2015. 狂犬病病毒核蛋白的结构、功能及其应用研究进展[J]. 中国病原生物学杂志,10(5):472-475. [Lu J,Fu Y R,Yi Z J. 2015. Research on the structure,functions,and applications of the rabies nucleoprotein[J]. Journal of Pathogen Biology,10(5):472-475.]
罗廷荣,刘棋,熊毅,郑敏,韦显凯,李军,唐海波. 2017. 广西狂犬病防控进展[C]//中国畜牧兽医学会. 2017年中国狂犬病年会论文集. [Luo T R,Liu Q,Xiong Y,Zheng M,Wei X K,Li J,Tang H B. 2017. Progress in prevention and control of rabies in Guangxi[C]//China Animal Husbandry and Veterinary Society. 2017 Chinese Rabies Annual Conference Proceedings.]
罗廷荣,张天生,邵帅,陆专灵,谢琪辉,韦友传,刘棋,熊毅. 2007. 广西狂犬病的流行病学[C]//中国畜牧兽医学会. 中国畜牧兽医学会动物传染病学分会第十二次学术研讨会论文集. [Luo T R,Zhang T S,Shao S,Lu Z L,Xie Q H,Wei Y C,Liu Q,Xiong Y. 2007. Epidemiology of rabies in Guangxi[C]//China Animal Husbandry and Ve-terinary Society. Proceedings of the 12th Academic Symposium of Animal Infectious Diseases Branch of China Animal Husbandry and Veterinary Society.]
彭娇娇. 2016. 狂犬病病毒诱导人和鼠神经母细胞瘤细胞自噬的研究[D]. 广州:华南农业大学. [Peng J J. 2016. Wild-type rabies virus induces autophagy in human and mouse neuroblastoma cell lines[D]. Guangzhou:South China Agricultural University.]
譚业平,赵静,郭霄峰,何孔旺. 2018. 一株携带犬细小病毒VP2融合基因的重组狂犬病毒的构建[J]. 江苏农业学报,34(4):866-870. [Tan Y P,Zhao J,Guo X F,He K W. 2018. Construction of a recombinant rabies virus carrying canine parvovirus VP2 fusion gene[J]. Jiangsu Journal of Agricultural Sciences,34(4):866-870.]
王朝,周明,傅振芳,趙凌. 2017. 狂犬病病毒逃逸宿主天然免疫反应的研究进展[J]. 生命科学,29(3):237-244. [Wang C,Zhou M,Fu Z F,Zhao L. 2017. The research progress on the mechanism of evasion from the host innate immune response by rabies virus[J]. Chinese Bulletin of Life Scien-ces,29(3):237-244.]
汪孟航,朱洪伟,何民辉,温永俊,程世鹏. 2016. 狂犬病病毒糖蛋白生物学功能研究进展[J]. 中国畜牧兽医,43(12):3349-3355. [Wang M H,Zhu H W,He M H,Wen Y J,Cheng S P. 2016. Advances in biological function of rabies virus glycoprotein[J]. China Animal Husbandry & Veterinary Medicine,43(12):3349-3355.]
熊毅,刘棋,盘龙波,王雯,罗廷荣,朱伟,韦友传,陆专灵,郭建刚,李华明. 2008. 广西狂犬病分子流行病学的研究[J]. 西南农业学报,21(4):1131-1135. [Xiong Y,Liu Q,Pan L B,Wang W,Luo T R,Zhu W,Wei Y C,Lu Z L,Guo J G,Li H M. 2008. Study on molecular epidemiology of rabies of Guangxi[J]. Southwest China Journal of Agricultural Sciences,21(4):1131-1135.]
徐素萍,黄小芳,刘宇明,苏荷,李晓宁,罗廷荣. 2018. 4株狂犬病毒感染小鼠的临床症状及脑组织病理学观察[J]. 南方农业学报,49(3):586-591. [Xu S P,Huang X F,Liu Y M,Su H,Li X N,Luo T R. 2018. Clinical symptoms of mice infected with four rabies viruses and brain tissue histopathology observation[J]. Journal of Southern Agriculture,49(3):586-591.]
杨健,张红普,章民,李晓宁,何晓霞,谢琳娟,陆专灵,韦显凯,唐海波,罗廷荣. 2012. 广西流行狂犬病毒糖蛋白基因的遗传性分析[J]. 南方农业学报,43(10):1575-1579. [Yang J,Zhang H P,Zhang M,Li X N,He X X,Xie L J,Lu Z L,Wei X K,Tang H B,Luo T R. 2012. Genetic analysis of glycoprotein genes derived from the rabies virus epidemic in Guangxi[J]. Journal of Southern Agriculture,43(10):1575-1579.]
周松峰,廖文军,袁书智,覃绍敏,白安斌,吴健敏,陆芹章. 2011. 狂犬病毒G基因主要抗原表位区(Rmg)的表达及纯化[J]. 南方农业学报,42(10):1276-1279. [Zhou S F,Liao W J,Yuan S Z,Qin S M,Bai A B,Wu J M,Lu Q Z. 2011. Cloning and expression of main antigen epitope gene(Rmg) and purification of proteins[J]. Journal of Southern Agriculture,42(10):1276-1279.]
Faber M,Pulmanausahakul R,Hodawadekar S S,Spitsin S,McGettigan J P,Schnell M J,Dietzschold B. 2002. Overexpression of the rabies virus glycoprotein results in enhancement of apoptosis and antiviral immune response[J]. Journal of Virology,76(7):3374-3381.
Jackson A C. 2011. Research advances in rabies. Preface[J]. Advances in Virus Research,79:xvii. doi: 10.1016/B978-0-12-387040-7.00022-6.
Kouznetzoff A,Buckle M,Tordo N. 1998. Identification of a region of the rabies virus N protein involved in direct binding to the viral RNA[J]. The Journal of General Virology,79(5):1005-1013.
Li T,Pattnaik A K. 1999. Overlapping signals for transcription and replication at the 3' terminus of the vesicular stomatitis virus genome[J]. Journal of Virology,73(1):444-452.
Liu J,Wang H L,Gu J,Deng T J,Yuan Z C,Hu B,Xu Y B,Yan Y,Zan J,Liao M,DiCaprio E,Li J R,Su S,Zhou J Y. 2017. Becn1-dependent CASP2 incomplete autophagy induction by binding to rabies virus phosphoprotein[J]. Autophagy,13(4):739-753.
Tang H B,He X X,Zhong Y Z,Liao S H,Zhong T Z,Xie L J,Pan Y,Lu Z L,Wei X K,Luo Y. 2013. Complete genome sequence of a rabies virus isolate from cattle in Guangxi,Southern China[J]. Genome Announc,1(1). doi: 10.1128/genomeA.00137-12.
Tao L,Ge J,Wang X,Zhai H,Hua T,Zhao B,Kong D,Yang C,Chen H,Bu Z. 2010. Molecular basis of neurovirulence of flury rabies virus vaccine strains:Importance of the polymerase and the glycoprotein R333Q mutation[J]. Journal of Virology,84(17):8926-8936.
Tian D Y,Luo Z C,Zhou M,Li M M,Yu L,Wang C,Yuan J L,Li F,Tian B,Sui B K,Chen H C,Fu Z F,Zhao L. 2015. The critical role of K1685 and K1829 in the large protein of rabies virus on viral pathogenicity and immune evasion[J]. Journal of Virology,90:232-244.
Vidy A,El Bougrini J,Chelbi-Alix M K,Blondel D. 2007. The nucleocytoplasmic rabies virus p protein counteracts interferon signaling by inhibiting both nuclear accumulation and DNA binding of STAT1[J]. Journal of Virology,81(8):4255-4263.
Wei X K,He X X,Pan Y,Liu C,Tang H B,Zhong Y Z,Li X N,Liang J J,Luo T R. 2018. Evolutionary analysis of rabies virus isolates from Guangxi Province of southern China[J]. BMC Veterinary Research,14(1):188. doi: 10. 1186/s12917-018-1514-0.
Yamada K,Noguchi K,Nishizono A. 2014. Efficient N-glycosylation at position 37,but not at position 146,in the street rabies virus glycoprotein reduces pathogenicity[J]. Virus Research,179:169-176.
Zhang G Q,Wang H L,Mahmood F,Fu Z F. 2013. Rabies virus glycoprotein is an important determinant for the induction of innate immune responses and the pathogenic mechanisms[J]. Veterinary Microbiology,162(2-4):601-613.
(責任编辑 兰宗宝)
