朱鹮圈养种群与野外重引入种群行为差异研究
2020-05-06邱国强付春正张睿灿万秋红方盛国
邱国强 付春正 张睿灿 万秋红 方盛国*
(1.德清县珍稀野生动物繁育研究中心,德清,313200;2.浙江大学生命科学学院,生命系统稳态与保护教育部重点实验室,国家濒危野生动植物种质基因保护中心,杭州,310058)
动物的行为是动物在个体层面对外界环境的改变和内在生理状况的变化所做出的整体性反应,且具有一定的生物学意义[1]。动物只有借助行为,才能适应多变的外界环境,并以最适合的方式完成饮水、取食、寻找配偶、筑巢和繁殖后代等生命活动,从而最大限度地保证自身的存活以及子代的延续[1]。对动物的行为进行科学研究的起点和基础是正确而详细的收集和整理所研究动物的各种行为类型,即行为谱研究[1]。建立濒危物种的行为谱,可以为动物保护工作者提供濒危物种的行为基线,从而更好地保护这些濒危物种[2]。了解一个物种如何与同种其他个体、其他物种、人类以及环境如何相互作用是有效实施就地保护和迁地保护的必要条件[3-4]。通过比较不同群体的行为差异和评估同一群体对栖息环境变化的行为反应对于濒危物种的管理和保护均具有重要意义[2]。
朱鹮(Nipponianippon)是一种在全球范围内濒危的鸟类,隶属于鹈形目(Pelecaniformes),鹮科(Threskiornithidae)[5]。该鸟类曾广泛分布于东亚地区,西起中国甘肃,东至日本宫古;南起中国台湾,北到俄罗斯布拉戈维申斯克均有朱鹮分布[6-7]。20世纪中期,由于气候变化、朱鹮栖息地丧失、人类滥用农业化学品和对朱鹮的过度捕杀,使得该鸟类个体数量急剧下降并一度被认为在野外灭绝[7]。自1981年于中国陕西洋县发现仅存的2对朱鹮和3只幼鸟后,中国政府立即采取了一系列保护措施对该鸟类进行保护,如朱鹮栖息地保护、禁止捕杀朱鹮以及建立朱鹮保护机构使得该鸟数量和分布稳定增长[7-8]。至2016年该鸟至少分布在16个区域,总量接近3 000余只[9]。圈养朱鹮种群主要分布在洋县(中国陕西)、周至(中国陕西)、罗山(中国河南)、北京(中国)、德清(中国浙江)、峨眉山(中国四川)、上海(中国)、珠海(中国广东)、新潟(日本)和庆尚南道(韩国);野生朱鹮种群和重引入朱鹮种群主要分布在洋县(中国陕西)、宁陕(中国陕西)、千阳(中国陕西)、铜川(中国陕西)、罗山(中国河南)、德清(中国浙江)和新潟(日本)[9]。
圈养环境下的动物由于缺乏行为的表达空间和必要的行为刺激,经常表现出刻板行为,却丧失一些与繁殖和生存相关的必需行为,进而导致其繁殖能力和生存能力的下降[10]。圈养动物的行为多态性下降对于其重新回归自然和对野外生存环境的适应是十分不利的。动物只有保留丰富的行为多态性,才能够适应复杂多变的自然环境[10]。前人对朱鹮行为的研究多局限于通过观察朱鹮行为构建朱鹮行为谱,或仅对单一类别的朱鹮种群的行为或单一类别的朱鹮种群在不同时期的行为差异进行研究,如仅研究圈养朱鹮种群的行为[11-15]。并未将圈养朱鹮种群的行为与重引入朱鹮种群及野生种群的行为进行对比研究,使得我们对圈养环境下朱鹮行为的退化或重引入朱鹮种群对野外环境的适应缺乏了解。本研究通过对德清圈养朱鹮种群和野外重引入朱鹮种群的朱鹮行为进行观察,比较德清圈养朱鹮种群和野外重引入朱鹮种群的行为差异,并分析行为差异产生的原因,从而了解野外重引入朱鹮种群的野外适应情况,并为改善朱鹮圈养环境和更好地保护朱鹮提供参考。
1 研究地点和方法
1.1 研究地点
浙江德清下渚湖朱鹮繁育基地,位于浙江省德清县东南部,地理坐标为30°31′N,120°03′E。该繁育基地内圈养朱鹮种群是由2008年从陕西周至楼观台圈养朱鹮种群引入的5对朱鹮作为奠基者繁育而成,是中国南部第一个朱鹮圈养种群。2014年11月,33只圈养朱鹮个体于浙江德清下渚湖被野化放飞,建立了中国南部第一个朱鹮野外重引入种群。经过数年的保护繁育,截至2016年繁殖期结束,德清圈养朱鹮种群的朱鹮个体数量为179只,德清野外重引入朱鹮种群的朱鹮个体数量为65只。
1.2 研究方法和数据分析
本研究在德清圈养朱鹮种群中随机挑选15只朱鹮个体,在德清野外重引入朱鹮种群中随机挑选10只带有GPS信号发射器(HQBP2525,湖南环球信士科技有限公司)的朱鹮个体,于2014年12月—2015年9月、2016年3月—6月,利用双筒望远镜(施华洛世奇SLC 7X42,熊猫303FT 10X42)和数码相机(Canon EOS 760D),交替对德清圈养朱鹮种群和野外重引入种群的朱鹮个体进行行为观察和记录。观察采用扫描观察法结合焦点动物观察法的方式进行:扫描观察法设定为每5 min对预先选定的每一个观察个体的行为进行一次扫描并记录;焦点动物观察法,只关注一个个体,对观察目标的全部行为进行详细记录,包括行为的起止时间和行为细节。观察从每天7:00—11:00、12:00—17:00,每天记录9 h行为观察数据。遇恶劣天气和突发事件则顺延,共记录行为数据6 600个,有效观察时间540 h。此外,本研究还于2016年6月利用数字照度计(TES1330A)分别测量了德清圈养朱鹮种群的朱鹮和野外重引入朱鹮种群的朱鹮傍晚飞至夜宿栖杠(树)和清晨飞离夜宿栖杠(树)的光照强度,每组光照强度各记录6次,计24个数据。
基于以上数据,本文对德清圈养朱鹮种群和野外重引入朱鹮种群的朱鹮个体行为进行分类、描述,并比较德清圈养朱鹮种群和野外重引入朱鹮种群的朱鹮行为差异。数据分析利用SPSS 20.0(SPSS Inc.,Chicago,USA)和Microsoft Office Excel 2013进行。其中,德清圈养朱鹮种群和野外重引入朱鹮种群傍晚飞至夜宿栖杠(树)和清晨飞离夜宿栖杠(树)时光照强度的对比采用成对样本t检验;德清圈养朱鹮种群和野外重引入朱鹮种群的行为类别时间分配的比较采用卡方检验。
2 结果
2.1 朱鹮的行为分类及描述
本研究对德清圈养朱鹮种群和野外重引入朱鹮种群的朱鹮个体行为进行分类、描述和统计,按照朱鹮行为的功能和目的描述定义朱鹮行为计8类55种,其中停歇行为4种、警戒行为4种、整理行为10种、运动行为3种、社会行为8种、繁殖行为12种、觅食行为7种、刻板行为7种。具体行为分类和定义如下。
2.1.1 停歇行为(Resting behavior)
朱鹮的停歇行为指朱鹮在一个位置上保持一定时间的固定姿态,行为的主要目的为休憩,也包括许多同时发生的极短暂理羽等行为(图1)。
(1)站立:朱鹮以双腿或单腿支撑身体,身体保持不动,双翅收紧于体侧,头向前或侧方。
(2)站立睡眠:朱鹮以双腿或单腿支撑身体,身体保持不动,双翅收紧于体侧,头转向后方,喙插入背部羽毛中。
(3)静卧:朱鹮双腿蜷曲收置于胸腹部下方,以腹部接触地面或其他支持物,身体保持不动,双翅收紧于体侧,头向前或侧方。
(4)卧眠:朱鹮双腿蜷曲收置于胸腹部下方,以腹部接触地面或其他支持物,身体保持不动,双翅收紧于体侧,头转向后方,喙插入背部羽毛中。
2.1.2 警戒行为(Vigilant behavior)
朱鹮的警戒行为是朱鹮对外界异常情况的反应,行为的主要目的为发现危险并远离危险(图2)。
(1)张望:朱鹮保持前一个动作不动,同时颈部伸展,头略高于正常位置并向四周张望。
(2)惊叫:朱鹮连续发出“Ao-Ao”的单音节叫声,并停止其他动作。
(3)惊走:朱鹮受到外界刺激后,颈向前伸,头部略低于正常位置,快速行走,偶伴随有扇翅的动作。
(4)惊飞:朱鹮受到外界刺激后,猛力扇动双翅飞行,常伴随有惊叫行为。
2.1.3 整理行为(Maintenance behavior)
朱鹮的整理行为种类较多,行为的主要目的是为了使自身保持清洁舒适,行为通常多种组合在一起发生或循环往复(图3)。
(1)理羽:朱鹮用喙梳理羽毛的行为,通过转动头部和伸展翅膀,可以梳理背部、胸腹部和翅膀上的羽毛,梳理过程中伴随着喙的快速小幅度张合。
(2)抖羽:朱鹮抖动全身羽毛的行为,常见于理羽过程中。
(3)水浴:朱鹮站立于水池或池塘中,通过喙沾水梳理羽毛、下蹲使胸腹部羽毛沾水、双翅拍击水面等方式使全身羽毛浸水的行为,常见于炎热的夏季和繁殖期涂抹行为之前。

图1 停歇行为——静卧(A)与卧眠(B)Fig.1 Resting behavior—crouching(A)and crouching with sleep(B)

图2 警戒行为——张望Fig.2 Vigilant behavior—looking around

图3 整理行为——理羽Fig.3 Maintenance behavior—preening

图4 运动行为——行走Fig.4 Locomotive behavior—walking

图5 社会行为——驱赶、惧缩(A)与咬腿(B)Fig.5 Social behavior—expelling,shrinking(A)and biting other’s leg(B)
(4)抖尾:尾部快速左右摆动的行为。
(5)甩头:头部快速甩动以去除头部和喙部黏附的泥土等异物的行为。
(6)抓挠:朱鹮头颈向下伸,用一侧脚在头部和喙部抓挠以去除异物。
(7)涂抹:常见于水浴之后,朱鹮头颈向后弯曲,用枕部及脖子在背部和翅膀上涂抹分泌物。
(8)扇翅:朱鹮站立,头颈及胸部向上伸展,同时拍打双翅。
(9)展翅:站立不动,头颈略向前下方伸展,将一侧翅膀向侧后方打开。
(10)排便:尾部上翘,身体微蹲,粪便从泄殖腔排出的行为。
2.1.4 运动行为(Locomotive behavior)
朱鹮的运动行为包含行走、跳跃和飞行,该行为的目的是通过腿部、脚部及翅膀的运动行为改变自身的位置(图4)。
(1)行走:通过双脚交替移动使身体位置发生改变,行走过程中头部位置与平常一致,不会出现惊走时的头部低于正常位置的现象,行走速度也明显低于惊走速度。
(2)跳跃:通过双脚蹬地或栖架等使身体短时间腾空而改变位置的行为,有时伴随有双翅的扇动。
(3)飞行:凭借翅膀的扇动或借助气流在空中快速移动。
2.1.5 社会行为(Social behavior)
朱鹮的社会行为主要是通过发出声音,竖立羽毛等独特的动作与其他朱鹮个体进行交流,该行为的主要目的是个体与个体或群体间的信息传递(图5)。
(1)鸣叫:通过发出各种声音来进行信息传递的行为,在朱鹮的繁殖期其鸣叫行为比非繁殖期常见。
(2)竖羽:竖起头部后方冠羽的行为,常见于相互理羽过程中和假交尾之后。
(3)相互理羽:一只朱鹮给另一只朱鹮梳理胸部和颈部的羽毛。
(4)共鸣:由一只朱鹮先叫而另一只或几只跟随鸣叫的行为,常见于假交尾、打嘴和相互理羽结束之后。
(5)驱赶:头部和颈部向前伸并急冲向被驱赶对象。
(6)惧缩:由于其他朱鹮接近而出现的,身体向远离其他朱鹮接近一侧倾斜,并不断移动的行为。
(7)互啄:两只朱鹮头颈前伸,互相用喙啄击对方。
(8)咬腿:一只朱鹮用喙叼住另一只朱鹮的腿或脚并拉拽的行为。
2.1.6 繁殖行为(Reproductive behavior)
朱鹮在繁殖期特有的行为,包括求偶炫耀、交配和孵化及育雏的行为,行为的目的为繁殖后代(图6)。
(1)理巢:繁殖期朱鹮站立或卧于巢中,用喙整理巢材的行为。
(2)叼树枝炫耀:繁殖期雄性朱鹮用喙捡取树枝,并发出连续低鸣的炫耀行为。
(3)打嘴:雌雄朱鹮通过头部的快速摆动,使喙互相碰撞,常伴随有连续的低鸣。
(4)交尾:雌性朱鹮卧或蹲,雄性朱鹮踏上雌性背部做交尾状,雄性尾部下压,雌性尾部上翘,雌雄个体泄殖腔相互接触,常伴随有打嘴和低鸣,交尾结束后,雄性从雌性背部跳下,雌雄共同高声鸣叫。
第二,“互联网+养老”模式有助于实现养老服务的全覆盖和专业化提升。在大数据技术的支持下,“互联网+养老”模式可以针对不同养老需求提供多层次、多样化、个性化的具体服务方式,使“互联网+养老”模式呈现全方位养老服务部署的态势,从而实现养老服务对象、内容、过程和环节的全覆盖。另外,由于大数据技术可以提供精准信息分析,帮助工作人员和机构能够提供更为精确的服务,这也养老服务行业的专业化提升奠定了基础。
(5)假交尾:雌性朱鹮卧或蹲,雄性朱鹮踏上雌性背部做交尾状,常伴随有打嘴和低鸣,但雌雄个体泄殖腔不接触。
(6)产卵:雌性朱鹮双腿微曲,身体下蹲,翅膀收紧在身体两侧并略微向下打开,行为持续时间较长直到卵产出。
(7)坐巢孵化:雌性朱鹮或雄性朱鹮卧于巢中,用腹部羽毛覆盖卵进行孵化的行为。
(8)换孵:雌性和雄性朱鹮轮流孵化的行为,通常一只孵化,一只飞出觅食或站立于巢旁树枝休憩和警戒。
(9)翻卵:朱鹮暂时停止孵化行为,从巢中站起,通过喙的拨动,改变卵的位置使卵温度均衡。
(10)晾卵:朱鹮暂时停止孵化行为,从巢中站起或离巢的行为,有时伴随有理巢行为的发生。
(11)暖雏:朱鹮将雏鸟覆于羽毛下,维持雏鸟体温的行为。
2.1.7 觅食行为(Foraging behavior)
朱鹮的觅食行为包括搜寻、捕捉、处理和摄取食物等行为(图7)。
(1)低头寻觅:常见于圈养朱鹮个体,朱鹮低头,颈向下伸,来回行走寻找食物。
(2)划动寻觅:常见于野外朱鹮个体,朱鹮站立于水质较浑浊的觅食地中,通过头颈的左右大范围划动,依靠喙来寻找食物。
(3)掏挖:朱鹮将喙伸入地面洞中,头颈扭动且喙快速张合。
(4)探啄:用喙在地面快速连续啄击,并不断移动。
(5)咬甩:朱鹮将食物叼住并猛甩头部,有时还将猎物向地面摔打,是一种常见的处理活猎物的行为。
(6)洗涮:用喙叼住猎物后将猎物浸入水中清洗,以便吞咽。
(7)吞咽:朱鹮叼住食物后通过喙不断地张合调整猎物位置,调整完毕后头颈向斜上方猛伸多次,使猎物从喙向喉部移动,从而完成整个觅食动作。
2.1.8 刻板行为(Stereotypic behavior)
朱鹮的刻板行为指朱鹮表现出的一种无明显行为目的和功能,并且经常反复的行为(图8)。
(1)啄尼龙绳:野外朱鹮个体站立于野化训练笼舍顶,用喙反复啄笼舍顶的尼龙绳。
(2)啄树干:野外朱鹮个体站立于树干,用喙反复啄树干。
(3)啄栖架:圈养朱鹮个体站立于栖架或人工鸟巢边缘,用喙反复啄栖架。
(4)啄铁条:圈养朱鹮个体站立于栖架或人工鸟巢边缘,用喙反复啄支撑鸟巢和栖架的铁条。
(5)啄塑料管:圈养朱鹮个体站立于地面,用喙反复啄给池塘灌水的塑料管。
(6)啄尼龙网:圈养朱鹮个体站立于地面,用喙反复啄尼龙网。
(7)啄铁丝网:圈养朱鹮个体站立于地面,用喙反复啄铁丝网。

图6 繁殖行为——假交尾Fig.6 Reproductive behavior—pseudocopulation
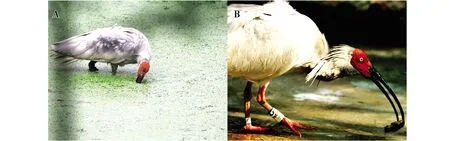
图7 觅食行为——划动寻觅(A)与咬甩(B)Fig.7 Foraging behavior—hunting with paddling(A)and biting with tossing(B)

图8 刻板行为——啄铁条(A)与啄尼龙网(B)Fig.8 Stereotypic behavior—pecking iron bar(A)and pecking nylon net(B)
2.2 圈养朱鹮种群和野外重引入朱鹮种群的行为差异比较
2.2.1 圈养朱鹮种群和野外重引入朱鹮种群傍晚飞至夜宿栖杠(树)和清晨飞离夜宿栖杠(树)时的光照强度比较
德清圈养朱鹮种群傍晚飞至夜宿栖杠时的光照强度显著低于野外重引入朱鹮种群傍晚飞至夜宿栖树时的光照强度(P<0.01,表1);并且,德清圈养朱鹮种群清晨飞离夜宿栖杠时的光照强度也显著低于野外重引入朱鹮种群清晨飞离夜宿栖树时的光照强度(P<0.01,表1)。
2.2.2 圈养朱鹮种群和野外重引入朱鹮种群的行为类别时间分配的比较
在8种行为类别中,德清圈养朱鹮种群和野外重引入朱鹮种群发生频次最多的行为均为停歇行为、其次均为整理行为,但德清圈养朱鹮种群和野外重引入朱鹮种群在行为类别的时间分配上具有显著差异(χ2=135.07,df=7,P<0.01,表2)。此外,在各行为类别的时间分配上,德清圈养朱鹮种群和野外重引入朱鹮种群在停歇行为、警戒行为、社会行为、繁殖行为和觅食行为上均存在显著差异(P<0.01,表2)。德清圈养朱鹮种群的停歇行为、繁殖行为均显著多于野外重引入朱鹮种群,而德清野外重引入朱鹮种群的警戒行为、社会行为和觅食行为显著多于圈养种群(P<0.01,表2)。
2.2.3 圈养朱鹮种群和野外重引入朱鹮种群行为日节律的比较
朱鹮的各类行为的发生在一天内具有一定的规律性:德清圈养朱鹮种群停歇行为在12:00—13:00、13:00—14:00出现低谷;整理行为在7:00—8:00和14:00—15:00出现高峰;运动行为和觅食行为的日节律变化具有一定的同步性,在 12:00—13:00出现一个较为明显的高峰(图9)。
德清野外重引入朱鹮种群的停歇行为在12:00—13:00,14:00—15:00出现低谷;整理行为的高峰发生在7:00—8:00和14:00—15:00,与圈养朱鹮种群整理行为的日节律高峰一致;觅食行为的高峰发生在8:00—9:00、12:00—13:00和14:00—15:00,相比圈养朱鹮种群,野外重引入朱鹮种群表现出较多的觅食高峰;此外,重引入朱鹮种群的运动行为和觅食行为的日节律也具有一定的同步性,在12:00—13:00和14:00—15:00表现为高峰(图10)。德清圈养朱鹮种群和野外重引入朱鹮种群的警戒行为、社会行为、其他行为(繁殖行为和刻板行为)的日节律均较为平稳,无明显的高峰和低谷出现(图9,图10)。
表1 德清圈养朱鹮种群和野外重引入朱鹮种群傍晚飞至夜宿栖杠(树)和清晨飞离夜宿栖杠(树)时的光照强度的比较

Tab.1 The comparison of the light intensity when the birds fly to the night-roosting perch at nightfall and fly away from the night-roosting perch in the morning between the captive and reintroduced population of the crested ibis in Deqing
表2 德清圈养朱鹮种群和野外重引入朱鹮种群的行为类型时间分配差异
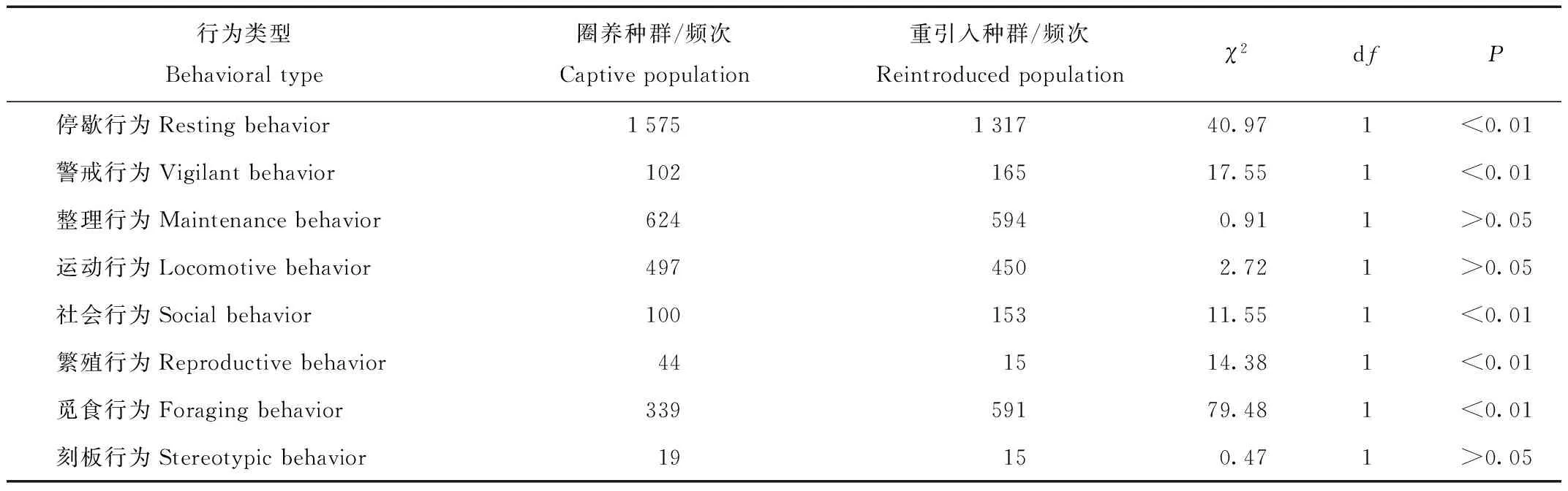
Tab.2 The differences of behavior time budget between the captive and reintroduced population of the crested ibis in Deqing
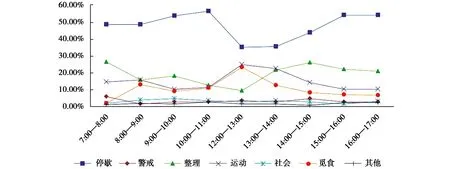
图9 德清圈养朱鹮种群的行为日节律Fig.9 Daily rhythm of the captive population of the crested ibis in Deqing

图10 德清野外重引入朱鹮种群的行为日节律Fig.10 Daily rhythm of the reintroduced population of the crested ibis in Deqing
3 讨论
动物的行为是动物适应外界环境的方式,也是其心理和意识的外部表现[16]。当动物所处的外界环境发生变化时,为适应新的生存环境,保证种族的延续,则需要动物的行为发生适应性的改变。相比于德清圈养朱鹮种群,野外重引入朱鹮种群傍晚飞至夜宿栖树和清晨飞离夜宿栖树时的光照强度显著较高,这一结果与野放前、后白鹇(Lophuranycthemera)傍晚上树开始夜栖和清晨下树结束夜栖时的光照强度的比较研究结果一致[17]。相比于圈养朱鹮种群,野外重引入朱鹮种群在傍晚飞至夜宿栖树开始夜栖和清晨飞离夜宿栖树结束夜栖时可能需要更高的光照强度,从而降低外界不利因素如夜栖树林区域郁闭度高造成的林内光照明暗不均、外界敌害等对自身的影响,进而提高自身的生存几率,更好地适应野外环境[17]。
动物各行为类型的时间分配是既对外界环境适应的表现,也是外界环境各种因素影响动物行为的综合表现[18]。德清圈养朱鹮种群和野外重引入朱鹮种群发生频次最多的行为均为停歇行为,其次均为整理行为,这一结果与阮祥锋等[19]对笼养朱鹮种群非繁殖期昼间行为时间分配的研究结果一致。而相对于德清野外重引入朱鹮种群,德清圈养朱鹮种群表现出显著多的停歇行为和繁殖行为,但表现出显著少的警戒行为、社会行为和觅食行为。这可能是由于在圈养环境下食物分布相对集中且充足,活动空间较小和个体密度较大,使得圈养朱鹮种群不需要花费过多的时间去觅食,从而导致圈养朱鹮种群表现出显著多的停歇行为和繁殖行为。而在野外环境下,食物分布相对零散且稀少,使得野外重引入朱鹮种群需要在多个地方觅食才能满足正常的生存所需;并且在农田、菜地或池塘觅食的过程中,野外重引入朱鹮种群遭遇其他动物及人类的机会相对较多,从而导致野外重引入朱鹮种群表现出显著较多的觅食行为、警戒行为和社会行为。野放前、后白鹇行为类型的时间分配比较也表现出相似的模式,如野放后雌性白鹇表现出显著多的觅食行为,且表现出显著少的停歇行为[17]。朱鹮的野外生存能力主要包括觅食能力和逃避敌害的能力,而相比德清圈养朱鹮种群,德清野外重引入朱鹮种群表现出显著多的觅食行为和警戒行为,这一结果与朱鹮野化训练前期与后期的行为差异对比的结果保持一致[15,20]。此外,相比于德清圈养朱鹮种群,野外重引入朱鹮种群在觅食行为的日节律上表现出较多的觅食高峰,但整理行为的日节律高峰与圈养朱鹮种群保持一致。德清野外重引入朱鹮种群觅食行为的高峰发生在8:00—9:00、12:00—13:00和14:00—15:00,这一结果与陕西宁陕县野外重引入朱鹮种群的觅食高峰发生时间基本一致[14]。相比德清圈养朱鹮种群,德清野外朱鹮种群在各行为类型的时间分配和行为日节律上的差异,可能是其在自觉或不自觉地调整自身的行为习惯,逐步适应野外的生存环境。
虽然德清圈养朱鹮种群和野外重引入朱鹮种群在刻板行为的时间分配上没有表现出显著差异,但德清圈养朱鹮种群表现出的刻板行为类型(啄栖架、啄铁条、啄塑料管、啄尼龙网、啄铁丝网)明显多于野外重引入朱鹮种群表现出的刻板行为类型(啄尼龙绳、啄树干)。在圈养与半野外大熊猫(Ailuropodamelanoleuca)行为研究中也发现相同现象,即相比于半野外环境下的大熊猫,圈养环境下的大熊猫表现出更多的刻板行为[21]。将动物重新引入到野外自然环境的目的是使其能够在曾经存在过的自然栖息地重建自然野外种群,但影响重引入成功的因素多种多样,而刻板行为就是一个主要的影响因素[21]。在圈养动物的日常管理中,减少圈养动物的刻板行为是一个重要的管理课题[21]。通过科学合理的环境丰容,有利于减少圈养动物的刻板行为,如通过环境丰容减少了圈养环境下赤斑羚(Naemorhedusbaileyi)和华南虎(Pantheratigrisamoyensis)的刻板行为[22-23]。因此,我们可以尝试运用环境丰容的方法来改善德清圈养朱鹮种群的生存条件,从而减少圈养朱鹮种群刻板行为的发生。
本研究通过比较、分析德清圈养朱鹮种群和野外重引入朱鹮种群的行为差异,了解到德清野外重引入朱鹮种群通过改变各行为类型的时间分配和日节律,已经开始逐步适应野外复杂的生存环境。但野外环境复杂多变,且该重引入朱鹮种群的野外栖息地毗邻下渚湖湿地风景区和多个村落,使其极易受到人类活动的干扰,因此为确保该重引入朱鹮种群进一步的适应野外栖息环境,仍需更多的观测调查。此外,德清下渚湖朱鹮繁育基地应进一步改善圈养朱鹮种群的生存环境,对圈养环境进行丰容,从而减少圈养朱鹮种群的刻板行为,使其产生更多的自然行为,为进一步进行野外放归,使其更好地适应野外环境奠定基础。
