不同种源楠木叶片表型性状变异分析
2020-05-05欧汉彪林建勇梁瑞龙
欧汉彪,林建勇,李 娟,梁瑞龙
(广西林业科学研究院/国家林业与草原局中南速生材繁育重点实验室/广西优良用材林资源培育重点实验室,广西 南宁 530002)
【研究意义】楠木(Phoebebournei)属樟科(Lauraceae)高大乔木,在我国主产于福建、浙江、湖北、湖南、江西、四川、重庆、广东和广西等地,大部分散生长于山地丘陵或溪沟旁。其树干通直、树形优美、材质细密且极耐腐,是我国特有珍贵阔叶树种,具有极高的经济价值和观赏价值。目前,楠木制作的高档家具、建筑、雕刻等闻名中外,优良材质让消费者趋之若鹜,市场供不应求[1]。由于人类长期强烈的干扰,导致楠木天然资源已急剧减少,因此楠木已被列为国家Ⅱ级野生濒危保护植物。已有研究表明,植物叶片性状受控于植物遗传特性,具有相对稳定性,但也受生境条件和气候特性等综合环境影响而表现出差异性[2-3],常作为区分品种间、种源间及家系间的重要生理性状指标[4]。因此,分析不同种源楠木的叶片形态性状及了解楠木不同种源间的遗传变异规律,对其良种选育具有重要意义。【前人研究进展】迄今,学者对楠木的研究主要集中在生理生态学[5]、育苗技术[6]、遗传改良[7]和造林技术[8]等方面,对其他叶片性状变异研究的树种有红锥(Castanopsishystix)[9]、刨花润楠(Machiluspauhoi)[10]、米老排(Mytilarialaosensis)[11]、苦楝(Meliaazedarach)[12]、厚朴(Magnoliaofficinalis)[3]、油茶(Camelliaolerfera)[14]等,并发现叶片形态性状对环境因子的响应表现出显著变异。植物群体在长期适应环境过程中会产生不同形式及程度的变异形态[15],叶片是决定植物生长的重要器官,具有较强的可塑性和环境敏感性[16-17]。在不同光照、温度、水分条件下植物叶片形态、内部结构和厚度会产生不同的变化[18]。表型多样性是遗传多样性的直接体现,是基因表达与环境因子在长期进化过程中共同作用的结果[3,19],而不同生境下植物种源的表型变异是遗传变异格局、适应机制和环境可塑性研究的前提[20]。叶片性状受控于植物遗传特性,故物种的叶片性状常被认为相对稳定[13]。此外,叶片性状还受生境条件和气候特性等综合环境影响而表现出差异性,常作为品种间、种源间及家系间的重要生理性状加以研究[21]。【本研究切入点】目前,关于楠木叶片表型性状变异的研究鲜见报道。【拟解决的关键问题】分析6个不同地理种源4年生楠木幼树的叶片形态变异情况,为楠木种质资源收集、保存、评价及其良种选育和开发利用提供参考依据。
1 材料与方法
1.1 试验地概况
试验在广西南宁市广西林业科学研究院苗圃进行,该区海拔约80 m,属北热带季风气候,年均气温21.7 ℃,年极端低温-1.5 ℃,年极端高温39.4 ℃,年均降水量1300 mm,日照时数1550 h以上,具有雨热同期、阳光充足等特征。土壤属赤红壤,pH约4.2。
1.2 试验材料
供试验6个楠木种源分别来自重庆江津、贵州思南、广西富川、湖南沅陵、江西崇义和福建建瓯的自然分布天然群落,于2015年12月采集种子进行育苗;2017年4月换大袋移植于直径20 cm、高20 cm的无纺布育苗袋继续培育,行株距为1.0 m×1.5 m;2018年4月移植于苗圃空地。各种源地理位置和气候因子概况见表1。
1.3 试验方法
1.3.1 试验设计 从本研究项目团队收集的楠木种源中,按照其天然群落地理分布范围,筛选6个最具代表性的种源,每个种源选取3株生长状态良好的幼树为重复,在每株树冠外层向阳面的枝条摘取30片长势良好、无病虫害的成熟叶片进行标记,装入密闭自封袋带回实验室测量性状指标。
1.3.2 测定指标及方法 叶片性状指标中,叶面积、叶周长、叶长、叶宽、长宽比和形状因子(叶片面积与周长的校正比)使用手持式激光叶面积仪CI-203(美国CID公司)测定,叶厚用游标卡尺测量,叶柄长和宽基距用直尺测量,叶基角、叶尖角和叶脉角用量角器测量。
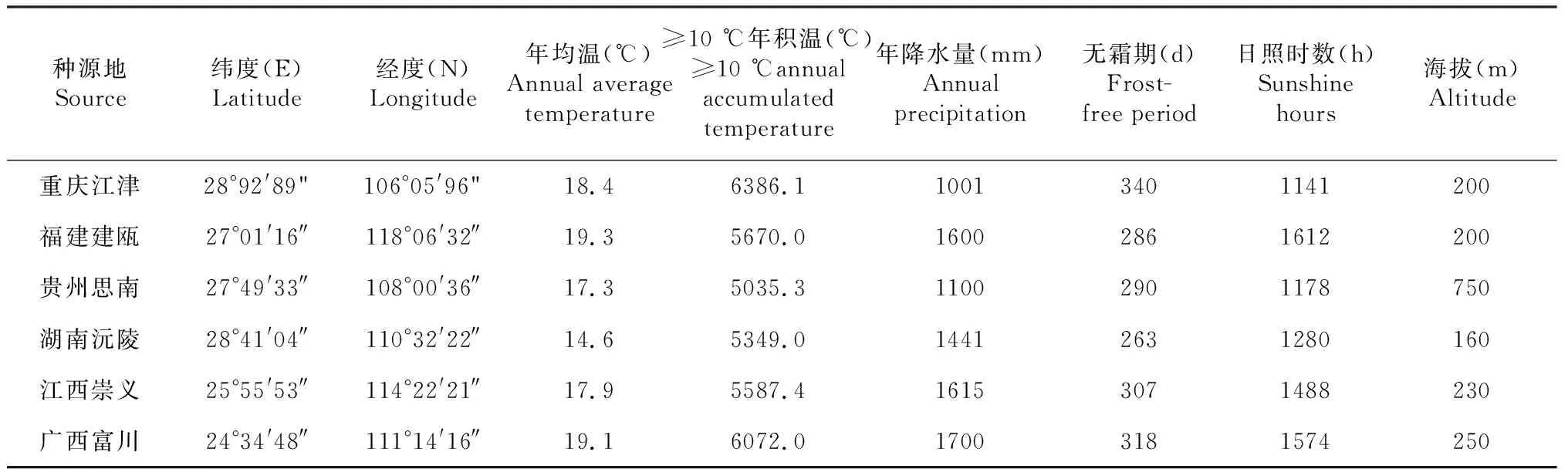
表1 6个天然楠木种源概况Table 1 Overview of 6 natural populations of P. bournei

表2 楠木叶片表型性状的方差分量及种源间的表型分化系数Table 2 The variance component of phenotypic traits of P. bournei and the phenotypic differentiation coefficient between populations
注:*表示差异显著(P<0.05),**表示差异极显著(P<0.01),下同。
Note:* represented significant difference(P<0.05),** represented extremely significant difference(P<0.01),the same as below.
1.4 统计分析

2 结果与分析
2.1 楠木叶片的表型变异分析结果
由表2可知,楠木叶片表型性状在种源内和种源间均存在极显著差异(P<0.01,下同),其中叶柄长、叶长和叶面积在种源间的方差分量百分比在30.00 %以上,说明这3个叶片表型性状在种源间变异较大,易受地理位置的影响,而其他性状变异主要表现在种源内,易受环境因子影响;叶柄长、叶厚、叶面积、叶基角和叶长5个性状的表型分化系数均大于60.00 %,说明这5个性状在种源间变异较大,而长宽比和宽基距的表型分化系数较小,为3.79 %和3.84 %,即该2个性状在种源间表现相对稳定。楠木叶片表型性状在种源间的方差分量百分比(18.01 %)大于种源内的方差分量百分比(17.92 %) ,说明叶片的表型性状以种源间分化为主。
由表3可知,楠木叶片表型性状表现出明显的变异,变幅为11.96 %~31.97 %。其中,叶基角的变异系数最大,为31.97 %,最小值为10°,最大值为65°,极差达55°;叶柄长的变异系数较大,为27.02 %,最小值0.15 cm,最大值1.66 cm,极差达1.51 cm;叶尖角的变异系数也较大,为24.04 %,最小值27°,最大值89°,极差达62°;叶厚的变异系数最小,为11.96 %;其他叶表型性状变异的变幅在12.09 %~19.92 %。说明叶基角和叶尖角的变异非常明显,离散程度大,而叶厚性状较稳定。
由表4可知,福建建瓯种源的叶面积最大(26.82 cm2),重庆江川种源次之,二者差异不显著(P>0.05,下同),但显著大于其他种源(P<0.05,下同),而广西富川种源的叶面积最小(20.01 cm2),江西崇义种源次之,二者差异不显著,但显著小于其他种源;叶长以重庆江津种源最长(12.85 cm),福建建瓯和湖南沅陵种源次之,三者差异不显著,但显著长于其他种源,而江西崇义种源的叶长最短(10.36 cm),广西富川种源次之,二者差异显著且均显著短于其他种源;叶宽以贵州思南种源最宽(3.43 cm),重庆江津和福建建瓯种源次之,三者差异不显著,而广西富川种源的叶宽最窄(2.90 cm),江西崇义种源次之,二者差异不显著,但显著窄于其他种源;叶周长以湖南沅陵种源最长(28.95 cm),福建建瓯种源次之,二者差异不存在,而江西崇义种源的叶周长最短(25.25 cm),广西富川次之,二者差异不显著,但显著短于其他种源。说明重庆江津种源和福建建瓯种源的叶形整体偏大,广西富川种源和江西崇义种源的叶形整体偏小。

表3 楠木叶片表型性状的变异情况Table 3 Variation coefficient of phenotypic traits in P. bournei leaves
从表4可看出,重庆江津种源、福建建瓯种源、江西崇义种源、广西富川种源、湖南沅陵种源和贵州思南种源楠木的形状因子分别为0.42、0.41、0.40、0.38、0.37和0.36,长宽比分别为3.91、3.72、3.50、3.88、3.85和3.53,说明狭长形数值较小,宽阔形数值较大,楠木叶片具有向圆形变异的趋势。
一般气温高、阳光辐射程度较强会使叶片变厚。在表4中,不同种源楠木的叶厚排序为湖南沅陵种源(0.22 cm)>江西崇义种源(0.19 cm)=广西富川种源(0.19 cm)=福建建瓯种源(0.19 cm)>贵州思南种源(0.17 cm)>重庆江津种源(0.16 cm),其中重庆江津和贵州思南种源的叶厚较薄,二者差异不显著,但显著薄于其他种源,说明重庆江津和贵州思南种源较耐阴。
叶基角、叶尖角、宽基距、叶脉角和宽基距叶长比等指标可以表征心形叶的基部凹陷和先端削尖程度。从表4还可看出,叶柄长、宽基距和宽基距叶长均以湖南沅陵种源最大,即湖南沅陵种源叶片的基部较狭小;叶尖角、叶基角和叶脉角均以江西崇义种源最大,即江西崇义种源基部叶片的先端最宽阔。
2.2 楠木叶片表型性状与地理因子和气候因子的相关性
由表5可知,楠木的叶面积与叶长、叶宽和叶周长呈极显著正相关,相关指数均大于0.600,表明叶面积随着叶长、叶宽和叶周长的增加而增加,叶面积主要受这3个性状指标影响;与长宽比呈极显著负相关,与性状因子呈极显著正相关,说明叶片形态狭长则叶面积较小;与叶柄长、宽基距和宽基距叶长比呈极显著正相关,表明叶面积较大则叶柄也较长,且叶片最宽处离叶基较远。
楠木种源采集点位于东经106°03′35″~118°06′32″、北纬24°34′48″~28°55′44″,海拔160~750 m,包含楠木所有分布区域,各种源的地理位置和气候条件差异明显(表1)。相关性分析结果表明,经度与叶厚、叶柄长、叶尖角和叶基角呈极显著正相关,与叶长和叶脉角呈极显著负相关,表明楠木种源地从西到东其叶厚、叶柄长、叶尖角和叶基角随经度增加而增大,叶长和叶脉角随经度增加而减小,即不同种源地楠木叶片从西到东表现变近圆的变化趋势;纬度与叶面积、叶长、叶宽、叶周长、叶柄长和宽基距均呈极显著正相关,与叶尖角和叶基角呈极显著负相关,表明楠木种源地自南到北其叶面积、叶长、叶宽、叶周长、叶柄长、宽基距和叶脉角随纬度的增加而增大,叶尖角和叶基角随纬度的增加而减小,即不同种源地楠木叶片从南到北表现变狭长趋势;年降水量与叶面积、叶长、叶宽、叶周长、宽基距和叶脉角呈极显著负相关,表明随年降水量的增加叶片呈变小趋势;≥10 ℃年积温与叶宽、叶周长、叶柄长、宽基距和宽基距叶长比呈极显著负相关,表明随≥10 ℃年积温的增加叶片呈变窄趋势;海拔与叶宽和叶柄长呈极显著正相关,与长宽比、形状因子、叶厚、叶尖角和叶基角呈极显著负相关,表明随海拔的升高叶片呈变宽长趋势。

表4 6个种源楠木13个叶片表型性状的多重比较结果Table 4 Multiple comparisons of 13 leaf phenotypic traits in 6 provenances of P. bournei
注:同行数据后不同小写字母表示差异显著(P<0.05)。
Note:Different lowercase letters in the same row represented significant difference(P<0.05).

表5 楠木叶片表型性状间及其与地理因子的相关性分析结果Table 5 Analysis of phenotypic traits of P. bournei leaf and its correlation with geographical factors
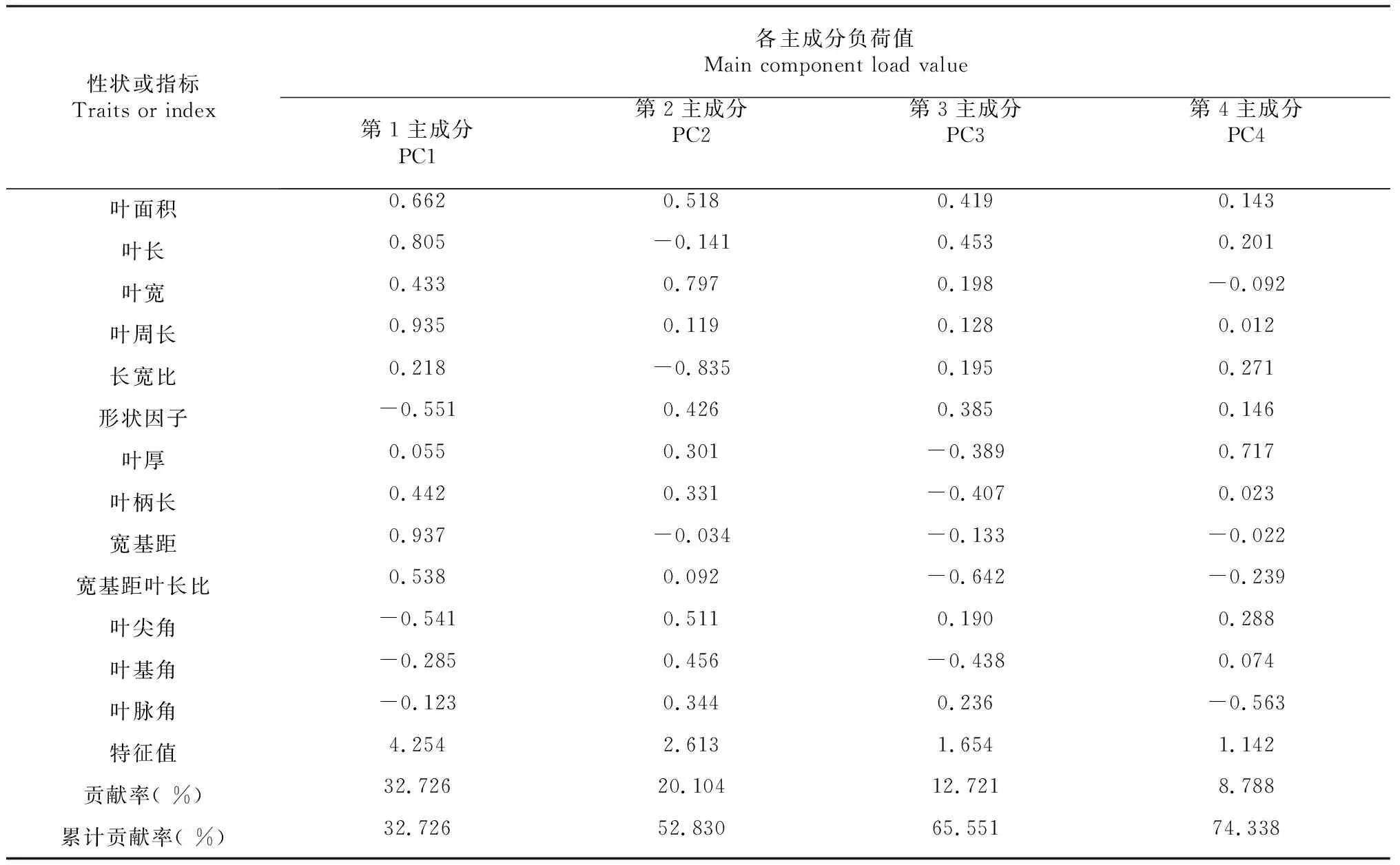
表6 楠木叶片表型性状的主成分分析结果Table 6 P. bournei leaf phenotype principal component analysis
2.3 楠木叶片表型性状的主成分分析
由表6可知,楠木叶片表型性状的第1~第4特征值分别为4.254、2.613、1.654和1.142,均大于1.000(第5个特征值为0.8270,小于1.000),故取前4个主成分进行分析。前4个主成分的累计贡献率为74.338 %,其中第1主成分贡献率为32.726 %,与叶面积、叶长、叶周长和宽基距相关度较高,主要表征叶片表型的叶大小性状;第2主成分贡献率为20.104 %,与叶面积、叶宽和叶尖角相关度较高,主要表征叶片表型的宽阔程度;第3主成分贡献率为12.721 %,与叶面积、叶长和形状因子相关度较高,主要表征叶片表型的狭长程度;第4主成分贡献率为8.788 %,与叶厚相关度较高,主要表征叶片表型的薄厚程度。
根据主成分分析结果,利用提取的4个主成分对楠木叶片表型性状进行聚类分析。从图1可看出,以欧式距离5为阈值,6个楠木种源可分为3个类群:广西富川种源和江西崇义种源为一类,处于楠木分布范围的南部;福建建瓯种源和湖南沅陵种源为一类,处于楠木分布范围的东北部;重庆江津种源和贵州思南种源为一类,处于楠木分布范围的西部。
3 讨 论
3.1 关于楠木叶片表型性状变异
本研究发现,6个楠木种源的叶片表型性状在种源间和种源内均存在极显著差异,说明楠木叶片表型性状在种源内的变异非常丰富。种源间的变异不仅受遗传多样性影响,也受长期地理阻隔、基因交流受限及地理环境因子变化的共同影响,其表型分化是在长期选择中的适应性进化,比种源内变异更有意义[2]。楠木叶片表型性状分化系数的均值达52.0 %,高于白皮松(Pinusbungeana)(Vst=22.8 %)[23]、复伞房蔷薇(RosabrunoniiLindl.)(Vst=33.1 %)[24]、川西云杉(Piceabalfouriana)(Vst=36.5 %)[25]、大花香水月季(R.odoratavar.gigantea)(Vst=37.3 %)[26]和紫丁香(Syringaoblata)(Vst=43.9 %)[27],小于蒙古栎(Quercusmongolica)(Vst=54.0 %)[28]、中甸刺玫(R.praelucens)(Vst=69.6 %)[29]、峨眉蔷薇(R.omeiensis)(Vst=73.6 %)[30]和云南黄连(Coptisteeta)(Vst=74.4 %)[31]。说明楠木叶片表型性状分化属于中等水平,种源间分化相对种源内稍高,受地理因子的影响较明显。
变异系数可揭示叶片表型性状的变异格局,变异系数越大,说明离散程度越大,性状越不稳定。本研究结果表明,楠木叶片表型性状变异幅度为11.96 %~31.97 %,高于红锥(0.67 %~26.09 %)[9]、水青树(Tetracentronsinense)(4.17 %~26.25 %)[32],低于苦楝(16.48 %~41.26 %)[12],与高州油茶(Camelliagauchowensis)(9.87 %~35.02 %)相近[14]。由于本研究选取的6个楠木种源涉及楠木分布的整个地理分布范围,能较明显反映叶片表型各器官的变异程度,有助于进一步开展叶片性状对楠木生长影响机理的研究。

图1 6个种源楠木的聚类分析结果Fig.1 Cluster diagram of six P. bournei provenances
叶片大小和形状能有效反映植物生存环境的变化状况,通过发生形态变化来调节水分蒸发和热量散失以适应环境[3];植物叶片具有较厚的角质层,一般可反映当地气温高、阳光辐射程度较强;叶柄长决定叶片的空间结构,可反映对光截留的程度。本研究结果表明,叶面积、叶长、叶宽、叶厚、叶柄长、叶基角和叶脉角的表型分化系数均高于50.0 %,说明楠木在不同生境中具有较大的表型性状变异。调查发现,相同种源同一批采集种子培育的3年生苗,其叶片在南宁市广西林业科学研究院苗圃和融水县贝江河林场种苗基地呈现较大的性状分化,南宁市的光热相对融水县充足,叶片表现更宽阔绵薄,进一步表明楠木对环境具有选择性适应性。
3.2 关于楠木的地理变异规律
不同树种的地理变异规律十分复杂,对环境的适应和敏感程度存在差异,随经度和纬度变化具有不同的变异模式,如苦楝的叶面积和叶宽与纬度呈显著正相关[12],高州油茶的叶形指数与经度和纬度呈显著负相关[14],厚朴的叶柄长和叶长与纬度呈显著负相关[13],红锥的叶长、叶宽和叶面积等与经度呈极显著负相关[9]。本研究发现,楠木叶片的表型性状与经度和纬度均呈极显著相关,与张凤良等[9]对红锥、陈丽君等[12]对苦楝的研究结果一致,整体来看,在水平上的渐变特征表现出连续性。
在长期的地理阻隔及基因交流受限下,植物为了适应环境会进行自然选择,由于气候具有区域性特征,植物也表现区域变异特征,通过种源试验可以揭示楠木种源变异情况,为划分种源区提供科学依据。本研究通过聚类分析发现,楠木具有地理来源的南部、东北部和西部3个不同群体特征,即具有区域性;结合相关性分析结果,楠木叶片表型形状变异规律具有地理连续性,说明楠木的地理变异规律属于连续性和区域板块性两种分布模式并存,与舒枭等[13]对厚朴的研究结果相似。
4 结 论
楠木叶片的表型性状在种源间和种源内均存在极显著差异,叶片的表型性状分化属于中等水平,种源间的变异大于种源内变异,受地理因子的影响较大。不同种源地楠木叶片具有从西到东表现变近圆趋势、从南到北表现变狭长趋势、随年降水量的增加表现变小趋势及随≥10 ℃年积温的增加表现变窄趋势的特征,可供楠木品种选育和利用研究参考。
