埃迪卡拉纪末期管状动物的“大辐射”
2020-04-29华洪蔡耀平闵筱柴姝代乔坤
华洪 蔡耀平 闵筱 柴姝 代乔坤


摘要:“寒武纪大爆发”是生物演化史上最重要的事件之一,现生动物的绝大多数门类在短短的两千万年的时间里“突然”出现,这一事件奠定了显生宙生物演化的基础。普遍认为以软躯体为代表的埃迪卡拉生物群是生命进化中“失败”的尝试,与显生宙生物之间没有明确的亲缘关系。文中通过对新元古代管状化石记录的系统梳理,基于对高家山生物群中大量管状动物化石类群如Cloudina,Sinotubulites,Conotubus,Gaojiashania,Shaanxilithes及可能的Anabarites等的较为系统的厘定和形态学复原,提出在埃迪卡拉纪,与典型埃迪卡拉软躯体生物群演化相对应,在新元古代末期,早期动物的进化史上存在着管状动物的大辐射,它们形态复杂,生物构型多样,个体大小差异悬殊,生活方式繁杂,代表了地球生命史上一次重大的生物革新事件,并且其中部分生物成为寒武演化生物群的先驱。该文将这一辐射演化事件称为管状动物的“大辐射”,它开启了显生宙生命演化的序幕。
关键词:埃迪卡拉纪末期;管状动物;大辐射;生物革新事件;高家山生物群
中图分类号:Q915.1
DOI:10.16152/j.cnki.xdxbzr.2020-02-001
"Tubular animal radiation" at the terminal Ediacaran stage
HUA HongCAI YaopingMIN Xiao CHAI ShuDAI Qiaokun1
Abstract: "Cambrian Explosion" is one of the most important events in the history of biological evolution, unparalleled diverse fossil fauna appeared abruptly in an interval not more than 20 million years at the beginning of the Cambrian. This event laid the foundation for the evolution of Phanerozoic organisms. It is generally accepted that the soft-bodied Ediacara biota is a "failed" experiment at the evolution of life, and there is no clear genetic relationship with the Phanerozoic organisms.Based on a comprehensive review of Neoproterozoic tubular fossil records, especialy on a more systematic identification and morphological restoration of a large number of tubular animal fossils in the Gaojiashan biota such as Cloudina, Sinotubulites, Conotubus, Gaojiashania, Shaanxilithes, and possibly Anabarites. It is proposed that in the Ediacaran Period, corresponding to the evolution of the typical Ediacaran soft-bodied biota, there was a large radiation of tubular animals in the evolutionary history of early lifes at the end of late Neoproterozoic. They are complex in morpho-types, diverse in bodyplans, varied in sizes and lifestyles, representing a major biological innovation in Earth′s life, some of which have been pioneers of the Cambrian Evolutionary Fauna. In this paper, the biodivercification event is called the "Tubular animal radiation", which opens the prologue of the evolution of Phanerozoic life.
Key words: terminal Ediacara; tubular animals; radiation; biological innovation event; Gaojiashan biota
傳统观点认为埃迪卡拉生物群与早寒武世复杂多样的小壳化石之间存在着极大的差异,以cloudinids等管状动物化石为代表的骨骼生物与寒武纪底部的多样化的以原牙形类和anabaritids等为代表的小壳化石动物群之间存在着截然的区别[1],埃迪卡拉末期出现的管状骨骼生物在寒武纪-前寒武纪界线附近发生了大灭绝 [2]。随着研究的深入,这一观点遇到了严峻的挑战。
早寒武世小壳化石中最为常见的化石类型是各类管状动物化石,它们是寒武动物群中十分重要的组成部分,但因其形态相对简单,对其属性研究尚显薄弱,至今大部分都被归入有疑问的化石类群(如:似软舌螺类(hyolithelminths),软舌螺类(hyoliths), 阿纳巴管类(anabaritids)等) [3-6]。管状动物的共同特点是均具有中空和细长的圆柱状或圆锥状的身体构型[7-9],它们在现代许多动物类群中都可以找到相应的代表,在系统分类上可以归属完全不同的生物类群,如多毛纲环节动物,昆虫,须腕动物,帚虫动物,轮虫,腹足纲动物和海葵等[10]。近年来不仅在以往缺乏化石证据的扬子地台朱家箐组待补段,陕西宁强宽川铺组碳硅质岩段、哈萨克斯坦Chulaktau组Aksai段发现了cloudinids管状化石(Cloudina, Rajatubulus)以及典型的寒武纪小壳化石如protoconodonts, maikhanellids, siphogonuchitids,及anabaritids和Cambrotubulus等。新组合中的一些管状小壳化石,如Rugatotecha,Anabarites 和Cambrotubulus,与典型的埃迪卡拉纪管状化石(如Sinotublites,Conotubus和Cloudina)在管壁形态和结构方面存在很大的相似性[11]。同时也发现埃迪卡拉纪典型的cloudinids 管状化石和寒武纪典型的Anabarites trisulcatus-Protohertzina anabarica化石带小壳化石存在着叠加混生现象,它们出现的时代早于寒武纪早期碳同位素负异常事件(BACE事件),即混生的化石群出现在寒武纪之前的地层中,寒武纪大爆发开始于前寒武纪,由此认为此前所谓的前寒武纪末期生物大灭绝事件是由于地质记录不完整所造成的假象,寒武纪生物群与前寒武纪晚期埃迪卡拉纪生物群之间具有逐渐过渡的演化关系[12]。
最新研究,Cai等[13]在陕西宁强地区的灯影组中发现了寒武纪早期小壳化石动物群中的重要分子Anabarites以及至少6~8种形态特征与寒武纪早期管状化石相似的类型,从而证实埃迪卡拉纪末期与寒武纪早期的部分骨骼管状化石在演化关系上存在密切的联系。同时表明埃迪卡拉纪末骨骼动物比以前认为的更为多样,埃迪卡拉纪-寒武纪过渡时期生物矿化动物的进化具有连续性[13]。
埃迪卡拉纪也不乏管状化石的报道,并且其类型复杂,生态多样,已被发现于几乎所有的岩性中,包括碳酸盐岩[14]、磷酸岩[15]、页岩[16-17]、粉砂岩[18-20]、砂岩[21]。 它们可能保存为钙质骨骼[14,22]、炭质压膜[16]、硅化或磷酸盐化的管[23-25],有些甚至保存为铸型[21],但多数化石的属性存疑,其中较为可靠的管状动物化石记录可以追溯到埃迪卡拉纪晚期,但它们的生物亲缘性依然存在争议[1]。本文通过对埃迪卡拉纪末期高家山生物群中几类典型管状动物化石的系统研究和分析,结合前人对埃迪卡拉纪管状动物的相关报道,提出在埃迪卡拉纪,伴随着典型的埃迪卡拉模块化构型的软躯体生物组合,存在着以管状动物为主导的演化模式,它始现于埃迪卡拉纪早期的蓝田生物群,在埃迪卡拉生物群白海组合(575Ma)中出现了一定的形态分化,在埃迪卡拉纪末期高家山生物群中其生物构型出现快速多样化,并出现部分寒武纪小壳动物的先驱,延续到早寒武世。本文将其生物辐射事件称为管状动物的“大辐射”(见图1)。
1 新元古代早期淮南生物群(850~720Ma)中管状化石
炭质宏体压膜状管状化石在世界各地新元古代盆地相页岩中广泛存在,它们呈简单的圆柱形管,具横脊、横纹或凸缘的圆柱形管,或具横隔壁的圆柱形管等[16,26-30]。中国新元古代炭质压膜保存的管状化石证据主要来自胶辽徐淮地区新元古代早期辽南生物群、淮南生物群,目前最新研究资料认为它的时代属拉伸纪,约850~720 Ma[31]。由于形态简单,并且呈二维压扁的保存方式,徐淮胶辽地区的具环纹的三类管状化石(Protoarenicola, Pararenicola和Sinosabellidites)的归属一直存在争议。最初将其作为两侧对称动物化石描述,并与环节动物和须腕动物进行对比[32-34]。Dong等[29]对其进行的深入研究,认为虽然其属性仍然存疑,但前人所报道的所谓吸盘状构造是埋藏假象,而固着构造的发现,证实它们可能是底栖固着直立生长的多细胞藻类化石。其中Protoarenicola在管体的一端具有一个球状膨胀结构,Pararenicola则在一端具有一个收缩的开口而另一端呈闭合状,而Sinosabellidites管体的两端都呈现出闭合的半圆状 (参见文献[29]及其同义名解释)。郑文武[35] 最初报道Sinosabellidites时,认为其可能是须腕动物皱节虫属。 Sun等[33]认为Pararenicola和Protoarenicola具有似吸盘状(proboscis-like)或者开口结构,因此它们可能是蠕虫类化石,而Sinosabellidites具有规则的横纹但不具有似鼻状以及开口等类似结构,所以是动物的可能性较小。Xiao等[27]通过对Protoarenicolid化石标本的观察,认为一些 Protoarenicolid 具有类似固着器的结构,进一步大胆地推测Sun等[33] 描述的似鼻状结构可能是保存不好的固着器。Dong 等[29]通过对这些化石的再研究,也支持上述观点,并认为这些化石与印度中部Vindhyan超群中的Tawuia化石相似[36],唯一的区别在于前者具有横纹,但横纹的存在不是动物属性的决定因素。如此看来,这些化石的动物属性的支持证据不足,它们更可能是管状的宏体藻类[37]。
2 埃迪卡拉纪陡山沱期管状化石
2.1 蓝田生物群(635~590 Ma)中管状化石
目前已知最早的可能后生动物是产自安徽蓝田生物群的化石代表,它们由5种形态可与现代刺胞动物或蠕虫类相比较的后生动物所组成,多具有固着器、触手或内部具类似肠道的特征,蓝田生物群的化石产自安徽休宁蓝田组二段。Wan等[38]在Yuan等[39]5个未定名形态种的基础上,将蓝田生物群中可疑的动物化石建立4个新属,5个新种,包括Lantianella laevis,L. annularis, Piyuania cyathiformis, Qianchuania fusiformis和Xiuningella rara。
Lantianella是厘米级、圆锥状化石,具球形的固着构造,顶部具不分枝的触手状结构。圆锥体由纵向的丝状体或纵脊组成,有时具有明显的環带。形态上它与蓝田生物群中十分常见,并解释为宏体藻类的Flabellophyton Chen等,1994,一致[40-42],因此Wan等[38]认为二者具亲缘关系,可能是埋藏所造成的偏差或代表不同系统发育阶段产物;其亲缘关系仍然不明,Lantianella可能是刺胞类水螅类,因为它的触手状结构和水螅类的触手在形态上有着惊人的相似性[38-39]。
Piyuania是一类锥形或杯状的化石,基部具固着构造。内部梭形结构被外部的杯状腔或鞘所包被,内部具有纵向丝状结构,其一端具有明显缩入的触须状构造。Piyuania cyathiformis为锥管状或杯状生物体,与Lantianella一样均具有触手状构造和丝状构造,Piyuania曾与刺胞类的水螅体进行对比[39]。
Qianchuania为纺锤形化石,内部具一个狭窄的纺锤形的内腔。一端具固着器(指定为底端),另一端具一个末端球形的指状构造(指定为远端)。与Lantianella的相似,但前者只有一根触手,推断可能属Lantianella发育的幼年阶段;与Piyuania的相同之处在于二者都具有一个内腔。虽然它们之间可能存在亲缘关系,但Qianchuania以其纺锤状总体形态与Lantianella 和 Piyuania有明显区别。
Xiuningella是管柱状化石,具短而窄的固着茎,球茎稍膨大。柱形管的中心具纵向似中轴的构造。Yuan 等[39]具有类似肠道结构和吸盘的蠕虫状动物,中轴推测代表消化道残余,底部球状构造和柄状构造代表吸盘。Wan 等[38]认为不排除它是底栖藻类化石的可能,球状构造代表固着构造,柄代表茎,圆柱状管代表多细胞丝状藻体。
2.2 埃迪卡拉纪陡山沱期翁安生物群(580~600 Ma)中管状化石
Liu等[28]在前人研究的基础上对埃迪卡拉系翁安生物群中磷酸盐化的管状化石进行了系统研究,鉴定出4个形态属,5个形态种,包括Ramitubus increscens, Ramitubus decrescens, Sinocyclocyclicus guizhouensis, Quadratitubus orbigoniatus, 及Crassitubus costatu。随后Liu等[43]又根据发现于三峡地区陡山沱组上部的硅化类型,建立一新属Yangtzitubus semiteres liu et al.,2009。最新SIMS锆石U-Pb年龄测定瓮安生物群的时代可能为609±5Ma,早于距今580Ma出现的埃迪卡拉动物群[44]。
陡山沱组磷酸盐化或硅化的管状化石中R. increscens, R. decrescens, S. guizhouensis和 C. costatus为圆柱状管,Q. orbigoniatus为四方柱状,上述类型均具有紧密分布的横隔壁, Ramitubus 多数具完整的横隔壁,而其他属种则具有规则交错或不完整的隔壁。R. increscens和R. decrescens具规则二分枝,C. costatus则具有纵向脊[28]。Yangtzitubus semiteres liu et al.,2009则为分枝状具多层状壁的半圆柱状管,内部中空无横隔壁[43]。薛耀松等[45]根据其形态特征将其解释为海百合类的茎和腕枝,随后, Li 等[46]报道了一些类似的微管状化石, 将其解释为小壳动物,Xiao等[15]对此类化石的形态特征及内部构造进行了细致的观察研究, 并对其亲缘关系进行了详细探讨, 根据其分叉方式、内部具横隔板(弯曲, 有时不完整)构造和体腔等特征, 指出此类化石可能属于原始刺丝胞动物, 与已绝灭的床板珊瑚类相似, 而与藻类及其他后生动物有明显的区别。Chen等[47]也指出此类化石属于刺丝胞动物床板珊瑚类,随后,Liu等[28,43,48]则继承了这一认识,但认为Ramitubus 可能为刺胞类基干类群、珊瑚虫纲动物或甚至可能的床板珊瑚,而Ramitubus, Sinocyclocyclicus,Quadratitubus, 和Crassitubus据一些不具分枝特征的微型管状化石在完整横隔板之间发育有不完整隔板、外部发育有具多层结构的外壁、并且一些标本管体内部发育有较宽的间隙等特征,认为这些特征可分别与丝状蓝细菌的细胞裂殖结构、外部胶鞘及藻殖段相对比, 指出一些不具分枝的管状微型化石可能为巨型蓝细菌[28,48]。随后的研究[48]指出具有分枝特征的Ramitubus确为底栖固着生活类型, 目前将其亲缘关系解释为床板珊瑚类是合适的;但不具分枝特征的Crassitubus, Sinocylocyclicus和Quadratitubus的古生态特征尚不清楚, 虽然它们与Ramitubus大小相近, 形态相似, 但现有的形态学特征似乎更支持将此类化石与丝状蓝细菌联系起来。因此,目前对陡山沱组管状化石的生物属性尚无定论。Cunningham等[50]利用最新科研手段对上述化石又进行了系统研究,结果表明Ramitubus和Crassitubus在管的最末端保存有特殊的细胞聚集体,而Sinocyclocyclicus和Ramitubus在横隔壁之间的空腔中充填有细胞组织或亚细胞组织结构,而这些空腔在刺胞类生物中属于以前水螅体生活的位置,后面被放弃掉的部分。上述发现都表明它们可能不是刺胞类生物,更可能是藻类。
2.3 埃迪卡拉纪陡山沱期庙河生物群中管状动物化石
庙河生物群产于陡山沱组上部黑色页岩中, 该化石组合不仅包括形态多样的宏体藻类化石,还有多个管状、锥管状化石被解释为可能的后生动物腔肠动物栖管化石,如:环纹杯状管(Calyptrina striata)、小型原锥虫(Protoconites minor)、简单九曲垴虫(Jiuqunaoella simplicis)、似僧帽管(Cucullus fraudulentus)和标准震旦海绵(Sinospongia typica)等。其中Jiuqunaoella simplicis, Sinospongia typica和Calyptrina striata曾与环节动物或蠕形动物进行过对比[51-53]。然而,后人研究表明Jiuqunaoella simplicis和Sinospongia typica被解釋为分节构造的的横向环纹更可能是化石压实过程中形成的皱褶[16,54-55],而所谓头部构造普遍保存极差无法辨识。曾经作为多孔动物海绵类描述的僧帽管属[54]和震旦海绵属[56], 至今在标本中没有发现任何骨针状构造, 并且它们的形态结构与腔肠动物和绿藻门的蠕藻类(Neomerids)都有类似之处[55],但与华南早寒武世地层中发现的[57-60]和国外的中寒武世布尔吉斯页岩生物群中[61]的海绵动物化石差异很大, 生物属性存疑。
Calyptrina striata: 以具有较为规则、相隔较远的横向有机质环带而与蠕虫或须腕动物进行对比[52,62-63]。Xiao等[16]推测Calyptrina striata为一圆柱形管,通过周期性的增粗在顶端生长,形成环檐构造。这一生长模式与蠕虫管类似,而与藻类不同。Wener[64]认为它应该属刺胞类生物形成的有机质鞘。 Calyptrina目前还发现于俄罗斯地台埃迪卡拉系Redkino层系的最底部层位[65]。王约等[66]则依据在江口翁会发现的锥形碳质压膜化石表面观察到横纹和中脊构造, 确认这类化石为锥管状的底栖后生动物。唐烽等[55]对近于同期的翁会生物群研究观察, 认为所谓的横纹和中脊构造是碳质压膜风化形成的构造假象, 并且该类标本常呈弯曲保存, 尤其在锥状体扩展端具扭曲现象, 并保存有浑圆状末端,认为具多细胞藻类的属性, 锥体薄膜状外壁可能是藻体分泌的黏液膜状保存的结果。
Protoconites:郭俊锋等[67]在早寒武世岩家河生物群中发现了与埃迪卡拉纪庙河生物群中Protoconites形态、大小等几乎一致的化石,并且认为Protoconites与寒武纪早期澄江化石库中的Cambrorhytium(或Archotuba conoidalis)和寒武纪中期的Cambrorhytium major无论在大小、形态,还是生活方式均存在着极大相似性,Cambrorhytium major已被解释成可能的刺丝胞动物[68],或与珊瑚虫出芽的外壳(如Stephanoscyphus[64]) 可以对比[16],暗示着Protoconites在生物演化上可能具有承上启下的作用。
3 埃迪卡拉纪灯影期管状化石
3.1 灯影期石板滩生物群中的管状化石
灯影期(5.5~5.4亿年前)的石板滩生物群因发现与国外埃迪卡拉生物群白海组合和纳玛组合中相同的软躯体动物化石而引人注目[69],该生物群除发现特征的软躯体化石Aspidella,Pteridinium,Rangea,Charnia,Dickinsonia和Hiemalora,以及大量遺迹化石外,其中占主导地位的是一类锥管状化石——雾河管(Wutubus)。近来具复杂分节和两侧对称构型的夷陵虫(Yilingia)的发现再次吸引了世人的目光[70]。
务河管(Wutubus annularis):是一类具有横向环纹的圆锥管状化石,直或微弯,毫米至厘米级,具有圆锥形的顶端和稍微膨胀但大致呈圆柱形的管。 其亲缘关系仍然存疑,但它在体型结构与许多典型的埃迪卡拉类生物类似,是一个单列的模块状建构的生物体。对其生态复原认为它生活在沉积物表面,营直立底栖固着生活。目前,该化石主要见于三峡埃迪卡拉系灯影组,同时在美国内华达Dunfee山埃迪卡拉系Deep Spring组底部也有报道[71]。
夷陵虫(Yilingia):与它的“临终遗迹”共同保存,是一类细长、分节,总体呈链状的两侧对称动物[70]。身体三分,有较为复杂的三叶形身体单元通过模块化重复链接而成,每个模块由一个菱形的中央叶和两个始终朝向身体一端(解释为身体的后端)的近三角形侧叶组成,身体两侧对称,不仅有体环,还有体节,分节数可超过50节。带状的身体向(假定的)前端略变细,并且在后端略变窄。身体分区和头化不明显。身体宽7~25 mm,体长76~608 mm。夷陵虫的归属,推测可能是环节动物或泛节肢动物(包括节肢动物、缓步动物、有爪动物)。这一发现证明,泛节肢动物或者它们的共同祖先在5.5亿年之前已经出现,并已显示初步的多样化,它们已具有两侧对称、分节、定向运动等特征[72]。目前仅在三峡埃迪卡拉系灯影组石板滩段发现。
3.2 埃迪卡拉纪灯影期高家山生物群
高家山生物群是目前新元古代唯一一个以黄铁矿化三维保存的管状和锥管状化石为主导,兼有钙质、磷酸钙质等骨骼生物,以及遗迹化石的多门类生物组合,高家山生物群在研究骨骼生物起源、宏体两侧对称动物的辐射演化等方面具有极为重要的意义。目前已经发现和报道的常见管状化石类群包括:陕西迹(Shaanxilithus)、锥管虫(Conotubus)、高家山虫(Gaojiashania)、克劳德管(Cloudina)、震旦管(Sinotubulites)及许多尚未描述并与早寒武世小壳化石有一定亲缘的微体管状化石,如可能的Anabarites等,是埃迪卡拉纪管状动物大辐射的最典型代表。
3.2.1 陕西迹(Shaanxilithus)
Shaanxilithes最初被描述为由一系列的“皱节”组成(体表的一系列横纹,并非真正的体节或是环节)的条带状构造,“皱节”的间距通常在 0.34 mm,标本的长度范围在 2~3 cm,宽度范围为1~5 mm[73]。
其属种类群包括一系列形态相似的条带状化石,如Shaanxilithes ningqiangensis,Shaanxilithes erodus,Helanoichnus helanensis,Palaeopascichnus等,上述不同类型化石通常保存在同一岩石层面上。近年来的研究成果均倾向于将上述条带状化石解释为Shaanxilithes ningqiangensis的同物异名,而它们之间的存在形态差别也被认为是埋藏差异的结果[74-77]。Mayer等[75]认为Shaanxilithes的保存方式与埃迪卡拉纪Fermeuse型及Burgess页岩型化石保存方式极为相似。
Meyer等[75]将Shaanxilithes三维形态复原为一系列“环状”或“饼状”结构组成的管柱状化石,在后期化石化过程中经常发生压扁、分离,从而保存为不同的形态特征。显然Shaanxilithes应为由一系列相同的结构模块连接形成的管状构造。而王欣[78]依据华北地台新元古代7个化石产地的资料以及大量的化石证据将Shaanxilithes的三维形态复原为 “管中管”结构,即化石由一系列套杯结构形成的内管和具有塑性(具褶皱)的薄膜状外管共同组成,而上述四类带状化石的形态差别由保存原因造成,并非生物的本来面貌。外管原始成分可能为柔软的有机质,具有一定的塑性且易于腐烂,起到包裹并固定内管的作用。内管成分可能是硬度稍大的几丁质,起到支撑和保护作用并形成管体空腔。
Shaanxilithes的生物属性始终存疑,该化石相继被解释为:须腕动物皱节虫类[73],沉积构造[79],遗迹化石[74,80-86],实体化石、可能的钙化藻类碎片[87]以及分类位置不明的后生动物实体化石[75-77,88-89]。特别需要指出的是,Tarhan等[76]对产自印度地区的Shaanxilithes进行拉曼光谱分析,并在化石中检测出成熟的干酪根,从而推定Shaanxilithes是一类具有机质壁的实体化石。王欣[78]的最新研究认为Shaanxilithes与现生深海管状蠕虫类在形态上具有极高的相似性,并倾向于将Shaanxilithes分类位置归为环节动物多毛纲。
对其生态恢复,王欣[78]推测Shaanxilithes生存于浅海环境中,其管体始端插入泥质或者砂质基底中营固着底栖生活。Shaanxilithes的内管由一系列的套杯状结构组成,类似于内骨骼并具有一定的机械强度(可能为几丁质),形成体腔同时起到支撑保护身体的作用。Shaanxilithes的外管由相对柔软的有机质组成,并具有一定的塑性,当管体在水流作用下发生弯曲变形,外管包裹内管保持生物的整体性(防止内管脱节),当外力作用消失后生物又重新恢复成近直管状形态。
Shaanxilithes最早报道于扬子板块西北缘陕南宁强地区晚埃迪卡拉世灯影组高家山段[73]。随后该化石相继在扬子地块云南东部渔户村组旧城段、湖北东南部老堡组、贵州清镇桃子冲组[77,79,87,90-91],以及西北地区相当于罗圈组冰碛岩之上,代表一套趋于正常海环境的地层中,如青海柴达木盆地北缘全吉群皱节山组[88,92];鄂尔多斯盆地西缘贺兰山地区正目观组上部[84,88,93-94]以及华北地台西南缘的陇县、洛南、豫西地区新元古代罗圈组冰碛岩之上的地层中发现[78]。
近年来,Shaanxilithes又相继在西伯利亚地区的Khatyspyt组、Yudoma组,印度的Krol和Tal群以及纳米比亚地区Nama群中有所发现和报道[12,74,76,86,89,95-96],其产出的地层时代均为埃迪卡拉世纪晚期。稳定的化石产出层位,较高的化石丰度,广泛的地理分布都表明Shaanxilithes具有成为晚埃迪卡拉世地层划分和对比的标准化石极大潜力[87]。
3.2.2 高家山虫(Gaojiashania) Gaojiashania化石个体较大,为直或弯曲的近于等粗的长管状,平行层面产出。管体由一系列密集圆环彼此连接在一起形成, 圆环由软组织连接起来,两个圆环之间的软组织发育有3~5条平行于圆环的环纹。圆环的大小基本不变,直径7~10 mm左右, 最大保存长度可达20cm 以上。其初期描述为体似蚕状,直或微弯曲,管壁由密集圆环(或皱脊)彼此连接而成[81-82],并且识别出背部-腹部、口部-尾部的形态学分异[97],Cai等[98]认为上述形态分异实际上是压扁作用、黄铁矿再生长/重结晶、保存不完整等原因造成的假象。陈哲等[99]通过对化石不同埋藏学特征推断,推测Gaojiashania是由一系列坚硬的圆环构成的管体,圆环由软体组织连接起来。Cai等[98]进一步研究发现圆环之间的薄层管体部分相对于圆环来说,厚度较薄,富有韧性,其上发育有横纹。此外,Gaojiashania管体发生的弯曲、伸长、收缩,表明这些发育横纹的薄层管体具有较强的柔韧性。虽然圆环可能未发生矿化或者只发生了轻微的矿化,但是它们相对来说比较坚硬。薄层管壁上发育有平行排列的横纹。在化石化过程中,薄层管壁容易发生腐烂,于是保存潜力很低。一旦薄管壁发生了腐烂,圆环就会发生分散甚至以单个圆环的方式散落在粉砂岩-泥岩层面上并平行岩石层面保存下来。
张录易[82]和林世敏等[81]对Gaojiashania属化石的形态学概貌作了简要的描述。先后建立了一个模式种(Gaojiashania cyclus Yang, Zhang et Lin 1986 (圆环高家山虫))及另外4个种,即C. zonatus Zhang, 1986 (环带高家山虫)、G. annulucosta Zhang, Li et Dong 1992 (环脊高家山虫)、G. caperata Zhang, Li et Dong 1992(起皱高家山虫)、G. haihaoliangensis Zhang, Li et Dong (海壕梁高家山虫)[97]。Cai 等[98]认为,Gaojiashania所有的5个种很大程度上属于埋藏变种,都应该合并为一个种,即林世敏等[81]建立的Gaojiashania模式种——Gaojiashania cyclus。
Gaojiashania在形态上类似于蠕形动物,前人将其作为蠕形动物化石并与环节动物多毛纲Oliveirania Maury, 1927 和Ruedemannella Howell, 1959 进行对比[82,97]。Gaojiashania与现代深海中的多毛类Siboglinidae科[100-101]动物相似。然而,Siboglinidae科动物复杂以及多变的形态结构使G. cyclus与其对比关系依然存疑。
化石保存于灰绿色薄层粉砂岩及灰岩互层中,平躺于层面,未见有斜穿层面的现象。在同一层面上,大小化石相差悬殊,最长可达十几厘米,最小仅几毫米,化石体主体无定向,保存完整。尤其是在层面上呈各个圆环叠置状保存的Gaojiashania化石,各个圆环间的连接部分已经完全腐烂,而各个圆环的相对位置却没有改变,在层面上仍相互联接呈带状分布,未经过搬运。因此,化石埋藏为原地保存,反映了生物原始生活状态,属于浅海底栖的类型。综合形态学、埋藏学、古生态学资料推斷Gaojiashania是一类表栖、平躺在水体-沉积物界面上生活的生物。相对坚硬的圆环提供了充足的管体固着到基底上的接触点,薄层管壁较强的柔韧性保证了G. cyclus可以进行自由的弯曲、伸长、收缩。而这些形态学及生态学特征可能暗示G. cyclus已经具有较强的运动能力,这些重要的形态学-生态学信息不仅指示Gaojiashania为匍匐平躺的底栖生活方式,同时还指示它们可能的真后生动物属性。
主要见于陕西宁强地区的埃迪卡拉系灯影组高家山段,也见于美国内华达Dunfee山埃迪卡拉系Deep Spring组底部及美国内华达Nye县埃迪卡拉系Wood Canyon组下段。近年来,在纳玛生物群主要产出地纳米比亚的Donkergange农场 Hoogland 段云母石英砂岩中中也发现了可能的Gaojiashania[72],并且具有火山灰U-Pb定年547.32+0.65 Ma[102]。其时代与高家山段碎屑锆石的定年值548 ± 8 Ma[103]比较接近。这也表明随着研究的进一步深入,Gaojiashania也可能具有潜在的地层意义。
3.2.3 锥管虫(Conotubus)
Conotubus Zhang and lin 1986最初由张录易和林世敏所命名[81],其特征为中空的扁锥状管体,直或自然弯曲,具明显的背腹之分,背部具有向尾端拱起的皱纹或褶皱,化石通常被黄铁矿化。随后的研究中张录易[82]报道了前端可能似触手状构造的存在,并对其进行了形态复原。Hua等[20]对其特征进行了修订,认为它是由一系列相互嵌套的漏斗所构成的锥管状化石,生物在光滑内管中生活,在生长过程中由于漏斗体管径逐步扩大,管体呈锥形,始端封闭,可能存在一个原始的胎管。与Cloudina的详细比较认为,二者壁壁结构和生活方式具有相似性,推断有机质或弱矿化的Conotubus可能是矿化的Cloudina的前身。Cai 等[104]进一步厘定为:微弯曲的管状化石,一端开口,另一端为圆形的封闭端。封闭端通常钻入下伏岩层中而不易观察到。未观察到特化的固着器等结构,而钝圆的封闭端可能起到固着管体于沉积物上的作用。管体可能未发生矿化并由一系列套叠的桶状管壁构成。桶状管壁具有模糊的纹饰。由于管体具有多层管壁套叠的结构,因此在横切面上能观察到多层管壁。在横切面上,管壁为圆形。新增加的管壁与较老的管壁相互叠压。
目前已经命名的属种有8个,包括Conotubus hemiannulatus Zhang and Lin 1986, C. gaojiashanensis Zhang and Lin, 1986, C. mimicus Zhang and Lin, 1986, C. impolitus Zhang and Lin, 1986, C. hamatus Zhang, 1986, C. cephalotus Zhang, 1986, C. adpressus Zhang, 1986,及C. varius Zhang, Li, and Dong in Li et al., 1992。 Cai 等[104]认为Conotubus的原始鉴定特征主要是基于管体压扁程度和次生黄铁矿化过程中管壁厚度变化等埋藏学变异。因此将其中前7个属种都归入Conotubus. hemiannulatus Zhang and Lin 1986,而C. varius Zhang, Li, and Dong in Li et al., 1992.由于主要来自岩石薄片,并且其壳壁明显由碳酸盐岩所组成,其特征与Cloudina riemkeae一致。
一些遗迹化石在后续的厘定中也被认为是Conontubus hemiannultus的同物异名。包括:Arthrophycus sp.[81],Intrites cf. punctatus[81],Intrites cf. punctatus[82],Neonereites ichnosp.[81],Neonereites ichnosp[82],palaeopascichnus cf. delicatus[81],Planolites annularis[82],Planolites beverleyensis[82],Planolites ichnosp.[81],Planolites sp.[82],Skolithos isolatus[82]。雖然过去把它们归为遗迹化石,但是它们展示出以下这些特征:具有和Conontubus hemiannulatus相似的个体大小、模糊的纹饰(Palaeopascichnus cf. delicatus, Planolites ichnosp.,[81];Planolites annularis, Planolites beverleyensis, Planolites ichnosp.[82], )或者脊(Arthrophycus sp.,[81];Neonereites ichnosp.[82], ),桶状外形(之前被解释为垂直潜穴;Intrites cf. punctatus, [81];Skolithos isolatus, Intrites cf. punctatus[82])、以及较短的黄铁矿柱体(Neonereites ichnosp.[81])。这些特征暗示它们是纹饰保存较差或者保存不完整的Conotubus hemiannulatus[104]。
Conotubus hemiannulatus的亲缘关系还未知。在没有软体结构被保存的情况下,很难对它的亲缘关系做出解释。和其他埃迪卡拉类化石一样,缺乏后埃迪卡拉纪可类比的化石同样给C. hemiannulatus的亲缘关系归属带来难度。此外,C. hemiannulatus的亲缘关系归属还受到化石保存方式的影响:与磷酸盐化作用不同,黄铁矿化作用、碳质压膜、黏土矿物很难保存软体结构。但是,锥管状形态、开口端与封闭端的分异、套叠状管壁结构、以及古生态学方面的特征共同表明C. hemiannulatus应该是后生动物。Conotubus和C. hemiannulatus最初被解释为两侧对称的蠕虫动物[81-82]。然而,据笔者等研究,C. hemiannulatus背部-腹部的分异只是压实过程中产生的一种埋藏假象。一些现生生物 (例如:pogonophorans中的Lamellisabella, Polybrachia以及vestimentiferan; 见文献[105])也显示出套叠状的管壁结构,这与C. hemiannulatus的很相似。然而,pogonophorans中更复杂、更多变的形态结构使二者的对比存疑。
高家山生物群中的Conotubus化石具有原地埋藏特征,其始端 1/3 处常具有明显的弯转或折断特征,该化石的古生态学特征也被推测为始端管体插入泥质基底中营固着底栖生活[104-105]。
3.2.4 克劳德管(Cloudina)
Cloudina管体常呈不同程度弯曲,始端封闭,末端呈喇叭状开口,未见任何附着痕迹。Germs[22]将Cloudina的形态复原为由一系列截切的锥体套合而成的管状体, Grant[14]则在大量薄片观察的基础上对此类化石作了系统总结,并对该化石重新作了形态复原。他认为Cloudina具有锥管套锥管状构造,即每一套合的锥体在底部都是封闭的,生物只生活在最上面那个锥体中,因而类似于珊瑚等,可能属于同一级别的刺丝胞类生物,他的观点得到了绝大多数学者的认同。Hua等[24]通过磷酸盐化三维保存的化石研究认为Cloudina管体是由一系列漏斗状单元偏心状套合而形成。除了始端单元,其他单元体都没有封闭的底,因此所谓的横隔壁在Cloudina管体中并不存在。漏斗体在始端逐渐汇聚形成共同的内壁,构成表面光滑的内部中央管;末端张开形成喇叭状环檐,或多个环檐互相愈合形成环脊,漏斗体外表面常见粗细不等的环纹或皱褶,环纹间近于平行。横断面各漏斗体通常近圆形。
Cloudina的研究虽然历经30余年,但对其形态复原和属性归类一直颇富争议。Germs[22]对产自非洲西南部Nama群中的化石首次做了描述,并建立新属Cloudina Germs,1972。根据管体直径的大小将其分为Cloudina hartmannae和Cloudina riemkeae两个种,认为可能属于Cribricyathea,且与龙介类有着一定的关系。Yochelson和Herrera[107]将产自于阿根廷Serrade Villiam的管状化石定名为Cloudina? borrelloi,并将其归于Cloudina。Glaessner[108]在认同Germs观点的基础上进一步提出Cribricyatheans以及Cloudina完全可以与龙介类多毛类(Polychaetes)相比较。Hahn等[109]在重新研究了产于阿根廷的Cloudina? borrelloi的基础上,建立了新属Acuticloudina,将Cloudina? borrelloi归入Acuticloudina,并且将与Cloudina相似结构的管状化石重新建立新科Cloudinidae Hahn and Pflug, 1985,其中就包括Cloudina和Acuticloudina两个属。这一观点随后也得到了Conway Morris等[110]的赞同。张录易等[97]将陕南宁强地区李家沟剖面发现的相似化石定名为Cloudina sinensis和Cloudina lijiagouensis,并且将其归入Cloudinidae科。陈哲等[111]根据大量较为完整的孤立标本研究,建立了新的管体构造复原模式,并且确认了Cloudina管体具有两种结构类型,即具有共同外壁的A型和不具外壁锥管裸露的B型,而两种管体结构的区别就在于是否具有外壁的共壁包裹。Cai等[112]根据Cloudina管体纹饰方向和管体大小区别了李家沟剖面和纳米比亚的Cloudina,并划分了新种,Cloudina ningqiangensis Cai et al.,2017和Cloudina xuanjiangpingensis Cai et al.,2017。同时认为有些属种可能是由于保存或埋藏学变异所导致的同物异名,如C. sinensis和C. lijiagouensis可能是C. hartmannae。
Germs[22]最早注意到Cloudina存在出芽生殖现象, Gaucher等[18]在乌拉圭发现了更多出芽生殖的证据,支持其可能为刺丝胞类基干类群。Pernet[113]认为无性繁殖分叉后子管体管径急剧增大,与多毛类管体特征不符,但与刺胞类,尤其是珊瑚虫相似。随着研究的深入,笔者等[24]修订了Cloudina的结构,认为它具有漏斗套漏斗的结构,漏斗底部在内部愈合,形成统一的内部共管,横向的底壁并不存在,同时首次在克劳德管中发现了共管及出芽繁殖的现象,子体在母管体内部以出芽方式形成新的管体,其分枝方式与一些多毛类,如Salmacina和Filograna的无性繁殖方式十分相似,证明两者间存在一定的亲缘关系。从而为长期悬而未决的克劳德管归属问题找到了可以直接对比的论据。Vinn等[114]認为Cloudina管壁的微颗粒状超微结构与环节动物不符,后者或为球粒状(缨腮虫类),或为球柱状(缨腮虫类和丝鳃虫类),或呈拉长的棱柱状(多毛类),他们的观点得一些研究者的响应,Zhuravlev 等[1]提出Cloudina的管内二分枝生殖方式与多毛类在口部出芽生殖不符,更类似刺丝胞类的插入式出芽生殖,并且管体底部封闭也与刺丝胞类相似。
Germs[22]曾在切片中发现C. hartmannae具有出芽生殖的现象,但并未得到Grant[14]的肯定,张录易等[97]曾描述了一枚在管体中部有一球状的壳体,被多层壳壁半包裹。随后Hua等[24]发现了Cloudina的分枝形式,“芽状”分枝和“共管”型分枝化石,前者在母管体口部分出芽枝,但随后其中一个管体停止生长,呈芽状,另一个管体继续生长;后者母管体在距内一分为二,两个子管体近于等大,两个个体的横切面为近半圆形,在两管接触部形成共壁,外为同一个外管所包围。Min等[115]根据近期发现的Cloudina大量不同类型无性繁殖的资料,提出了Cloudina四种无性繁殖模式,而其中的“逃生舱”模式和“口部出芽”模式均是现生多毛类主要的无性繁殖分枝方式,同时指出虽然多毛类以释放大量配子的模式主要行有性繁殖,但在一些特殊环境下,尤其在聚集生活过程中,多毛类会采取无性裂殖的方式,母体将身体裂成两段或多段,由于母体在生活过程中通常不能完全离开自己栖居的管体,而它又堵住了管体中惟一的出口,幼体发育到一定程度,为独立生活,会在母管体上采取开孔的方式,在开孔处逐步分泌出子管体,因此无论是“逃生舱”模式、“口部出芽”模式、“内部共管”模式及“Y型”分枝模式,其目的只有一个:子体可以脱离母管体行独立生活。因此认为Cloudina的多种分枝模式支持其为多毛类的可能。
目前正式报道的Cloudina属种共有下列9个:Cloudina lucianoi,Cloudina hartmanae Germs,Cloudina riemkeae Germs,Cloudina waldei Hahn & phlug,Cloudina lijiagou,Cloudina sinensis,Cloudina carinata,Cloudina ningqiangensis Cai et al.,2017和Cloudina xuanjiangpingensis Cai et al.,2017[14,22,110,116]。
虽然Cloudina和伊迪卡拉可疑软躯体化石可以在多个地区的同期地层中相间出现,但从未发现在同一层面上共生的现象,这表明二者在微环境上存在差异。在许多Cloudina中由各漏斗状套管所形成的脊宽窄不一,反映生物体存在异速生长现象。Seilacher是第一个对Cloudina生活习性做出相对合理解释的学者。主流观点认为该化石管体始端可能插入(或固着于)微生物席营滤食生活[106,117-118],活动能力有限。Seilacher[117]认为Cloudina锥形的管体形态、各套管底部封闭的特征,符合“mat-sticker”的生活习性,其基部黏附或半埋在微生物藻席中,伴随着沉积物的加积其壳体向上垂直生长,像珊瑚等刺丝胞类生物一样随着沉积速率调整管体的生长速度,分泌出新的套管,而管体的生长速率与沉积速率相吻合。其软体部分在壳体中也不断上移,而它的软体只在最上部的那个套管内。而Cloudina管体的膝状弯曲也可以简单地解释为管体由于水流、扰动或其他因素所导致的倒伏[117]。在这一模式下,Cloudina具有滤食生物的特征,它向上的生长主要是为了维持滤食器官始终在底面沉积物之上,同时Cloudina的软体部分不需要随之拉长,它仅需要抬升身体位置即可,其生活方式与许多珊瑚类似。
对磷酸盐化三维立体保存的Cloudina化石的管体生长形态进行系统观察,一个明显的特征是这些管体表现出多变的生长方向和复杂的弯曲形态。这很可能反映了这些固着生物对生态环境变化或物种间生态空间竞争而产生的响应。在许多Cloudina中由各漏斗状套管所形成的脊宽窄不一,反映生物体存在异速生长现象。综合这些研究资料之后,Cai等[118]认为陕南地区的Cloudina至少有3种固着生活方式:①管体封闭端固着到发育微生物席的碳酸盐岩基底上;②部分管体埋在基底中生活;③部分管体平躺在基底上,同时部分管体直立伸入水体中生长[118]。
Cloudina具有较为稳定的时空分布,除华南地区外, Cloudina已在几乎所有大洲都有报道,包括西南非洲的纳米比亚[22,119], 南美洲的阿根廷[107]、巴西[109] 及巴拉圭Paraguay[18,120-123], 北美洲的美国内华达和加利福尼亚[124-125]及加拿大[126], 欧洲的西伯利亚地台[127-129], 西班牙中部[14,129-140],及阿曼南部 Salt Basin[2,110]。Cloudina被认为是新元古代末期最好的标志性化石[14,20,106,110,141-142]。
3.2.5 多锥管(Multiconotubus) 该属由Cai等[112]所新建,用来区分那些具有若干个深嵌套的锥形层组成的多层管壁、直或弯的锥形生物矿化管,其始端封闭,末端开口。在横切面上呈圆形,套锥体通常不在横切面和纵切面的中心。
与Conotubus[104] 和 Cloudina[118]的生活方式相似,Multiconotubus可能也是通过锥形管的始部与基底的锚定,营固着生活方式。Multiconotubus具有明显的中央管,表明管层可能是逐渐向内分泌形成的。动物的软体部分可能生活在中央管中,通过内部锥管的分泌起到了提升软体部分的作用,从而占据了更有利的生态位。其锥管形态和相关的古生态策略与早寒武世的一些物种(如锥管状小壳化石)相似,从而支持了它们在埃迪卡拉纪-寒武纪过渡期古生物学和古生态学上的进化连续性。该化石目前仅报道于陕西宁强地区。
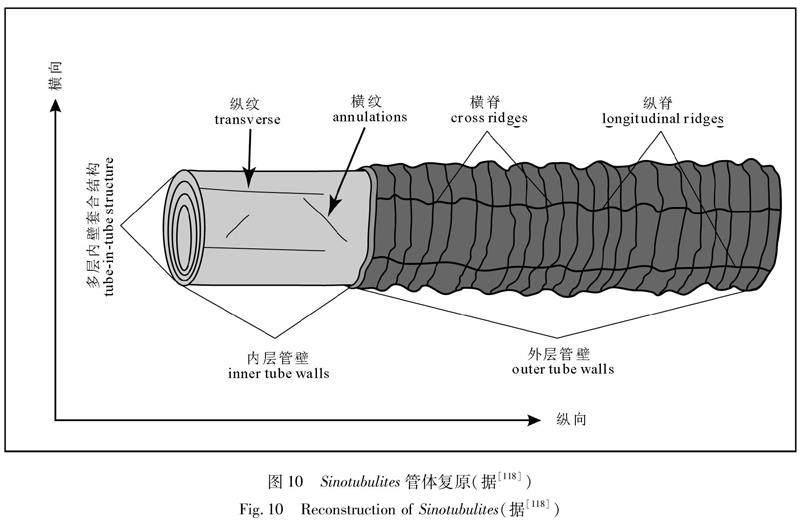
3.2.6 震旦管(Sinotubulites) Sinotubulites为管体直或微弯,两端开口的管状化石。长5~28 mm、直径2~5 mm。管体通常为等宽的圆柱状,由内外两部分构成:外部管体部分通常由多层不规则的褶皱状管壁构成,在横向和纵向上可见不规则的横脊和纵脊;内部管体部分由相对平滑的多层管壁组成。相对靠近管体外部的管壁套合相邻的、相对靠近管体内部的管壁,最终形成多层套管结构[111,143]。管壁总体光滑,表面发育稀疏的横纹和纵纹。相邻的管壁之间具有一定的间距。从管体横切面上可以观察到多层管壁的横切面构成的多个圆环,这些圆环通常具有一定的偏心率。外层管壁的纹饰密度不均一相对内层管壁的要粗大和显著的多。
陈孟莪等[144]最早在湖北三峡地区埃迪卡拉系灯影组白马沱段的白岩中发现了Sinotubulites,但当时的研究将其归入Cloudina sp.进行描述,还根据管体的直径尺寸将此管体分为大(4~5 mm)、中(1.5~3 mm)、小(0.4~1 mm)3种类型,将其归入环节动物门多毛纲龙介虫科。随后对其的重新研究[23]认为由于化石壳壁微细构造保存较差增长方式不明未见共壁或芽生现象不易与Cloudina作比较,因此暂定新属Sinotubulites,有疑问地归属于环节动物门龙介虫科。McMenamin[145]对发现于墨西哥Sonora西北部Caborca地区下寒武统La Cienega组及California的White Inyo地区可能属于下寒武统Tommotian阶的Deep Spring组的Sinotubulites進行了报道,把 Sinotubulites归入?Coleolidae 科并建立了新种Sinotubulites cienegensis McMenamin, 1985。此外,McMenamin [145]认为陈孟莪等[23]报道的大、中、小型3种管体均为同一个种:Sinotubulites baimatuoensis,同时他对Sinotubulites鉴定特征进行了补充,指出管表具有明显的条纹、条带甚至横脊,并且由纵向脊连接或切割。岳昭[146]将陕西宁强李家沟剖面磷酸盐化保存的具有清晰管壁构造的Sinotubulites分为3个种:Sinotubulites baimatuoensis,S. cienegensis和S. shaanxiensis并与 Cloudina,Rugatotheca以及Coleolella进行了对比研究,认为Sinotubulites可能为一种具口盖的虫管而并非动物体的外壳。张录易等[97]考虑到Sinotubulites模式标本的管壁全部被硅化、保存较差、未能查明其壳壁结构等原因,于是重新建立新属Qinella以示区别。华洪等[147]将Qinella属的模式种Q. shaanxiensis Zhang, Li et Dong, 1992 归入Sinotubulites Chen, Chen et Qian, 1981,但保留了Qinella,用来作为管壳外表面光滑的多层壁套管式结构类型Qinella levis。陈哲等[111]将 Qinella Zhang, Li et Dong, 1992归入Sinotubulites Chen, Chen et Qian, 1981, 并对属征进行了补充,同时依据管体大小和表面纹饰特征分为S. shaanxiensis和S. levis两个种,并对管体形态结构、成分、生态、归属等作了详细的研究。
Sinotubulites管体外壁上的横脊具有以下特征:横脊在外层管壁上的分布密度很不均一。在横脊密集的区域,横脊排列紧密,相邻横脊之间的间距相对较小,单个横脊相对高凸尖锐;在横脊稀疏的区域,横脊排列稀松,相邻横脊之间的间距相对较大,单个横脊相对中凸平缓。横脊与管体的长轴方向的夹角变化较大,大部分情况下横脊近于跟管体长轴方向垂直,少部分情况下横脊与管体长轴方向以低角度斜交。
Sinotubulites管体外壁上的纵脊具有以下特征:纵脊的条数变化很大,而且每条纵脊往往不连续。目前所观察到的、在整个管体上都连续发育的纵脊条数为3,5,6,8,大部分情况下为6条[148]。Sinotubulites管体的横切面常呈多边形,多边形断面的边数对应着纵脊的条数[143]。虽然在少部分情况下可以见到同一条纵脊沿着整个管体分布,在大致平行于管体长轴的方向上弯弯曲曲地延伸,但是在绝大部分情况下同一条纵脊在平行管体长轴的方向上往往不连续。最近Cai等[149]根据陕西宁强李家沟灯影组磷酸盐化保存的材料,对具有3,5,6辐射的类型,对应建立了3个新种:S. triangularis, S. pentacarinalis,和S. hexagonus Chen等[143]根据管体两端开口,管体上发育有纵脊以及管体缺乏固着结构等,认为Sinotubulites是一种表栖平躺的生物[111,143]。随后孙勃等[148]通过对Sinotubulites管体的生长特征以及不规则褶皱状外壁的形态变化研究进一步补充和支持Sinotubulites为表栖固着生物的形态学及功能学证据。Sinotubulites内部管体由多层管壁套合而成,这些特征表明管体不具有能发生自由弯曲的能力,而强烈弯曲的管体应该反映了Sinotubulites的生长特征,即:管体在活着的时候发生了弯曲生长。横脊的形态变化往往随着管体的弯曲而发生相应的形态变化。横脊之间的一小段纵脊往往会发生弯曲,这表明相邻的横脊之间的间距可以发生改变。同一管体上横脊的密度以及横脊与管体长轴之间的夹角也可以发生变化,这些特征都支持管体可以平躺在基底上,通过调整横脊之间的距离来达到伸展和收缩。横脊和纵脊的形态变化保证了管体的伸展和收缩。而横脊和纵脊的形态變化正好反映了Sinotubulites不规则褶皱状外壁相对较强的活动性。这些不规则的横脊和纵脊可以保证管体平躺在基地上,不至于受到水流或其他原因引起的扰动而发生翻转[143,149]。
3.2.7 高家山生物群中其他管状化石类群
高家山生物群中同时发现了大量与小壳化石可以对比的类群,如可能的Anabarites,Paragloborilus,Tiksitheca,Conotheca及Cambrotubulus等,显示出相当的分异度水平[13],目前尚有大量管径相对粗大,但表面缺乏纹饰的管状化石没有描述和研究。
可能的阿纳巴管(Anabarites sp.):根据Anabarites Missarzhevsky的鉴定标准[150],它的典型特征是 “钙质的管(鞘)或铸型,具有三辐射对称的纵向单元” (同时参考[151])。 高家山生物群中所发现的化石具有明显的三辐射状结构,表现为三条纵沟,与Anabarites的鉴定特征相符。
拟球管螺(未定种)(Paragloborilus sp.) (图13,A~C):管体有两部分组成,下部为一个球形封闭的始部,可能为胎管,一个顶部开口的锥状管,二者间被横向环沟所分隔的,与Paragloborilus Qian, 1977的鉴定特征吻合。
提克西螺(未定种)(Tiksitheca sp.) (图13,D~E).Tiksitheca Missarzhevsky的特征是具有三角形或亚三角形横截面的细长、光滑、圆锥形管 (参考[151])。目前的标本具有三角形的横截面,但目前标本为圆柱状而非Tiksitheca的椎管状。
寒武管螺(未定种)(Cambrotubulus sp.) (图13,L):Cambrotubulus Missarzhevsky的原始鉴定特征为具有不同形态及扩张速率的锥管,与anabaritids的区别在于在不同生长阶段其横截面都为圆形,但二者的管壁具有相似的微结构,并且均具有尖缩的始部和末端的开口[1511。目前标本具有近圆形的横截面,尖缩状的始部,和末端开口,可以暂时归入Cambrotubulus sp.。
Coleolella sp. (图13,O):Coleolella Missarzhevsky是一类锥管状化石,具有圆或亚圆形横截面,管体表面具横肋。目前发现的化石具锥管状形态,表面横肋十分发育,可以归入此类。
4 埃迪卡拉纪其他管状动物化石
世界其他地区的埃迪卡拉纪管状化石,主要来自几个方面:①出现在埃迪卡拉生物群组合中,并与典型软躯体化石组合相伴生,如加拿大纽芬兰阿瓦隆组合中的Thectardis,南澳Nilpena Rawnsley石英岩埃迪卡拉段(白海组合)中的Somatohelix、Funisia及Plexus;加拿大Mackenzie山Blueflower组(纳玛组合,同位素年龄553~541 Ma)的Sekwitubulus annulatus和Annulatubus flexuosu。②与埃迪卡拉生物组合交替出现,但不共生,代表二者生活环境的差异,如纳玛组合中的管状化石Cloudina,只出现在碳酸盐岩相中,与碎屑砂岩相中保存的软躯体组合明显有别③出现在一些与高家山生物群相类似,没有典型埃迪卡拉软躯体化石的地区,如乌拉圭、巴西、美国内华达等地区的埃迪卡拉系上部地层中,包括常见的Cloudina,Sinotubulites,也有如Corumbella,Soldadotubulus和Waltheria等特有类型,近期碎屑岩中也有Wutubus, Gaojiashania和Conotubus的报道。
埃迪卡拉生物群因最初发现于南澳大利亚埃迪卡拉山前寒武纪晚期的庞德砂岩而得名,是已知最古老的具有复杂体型结构的生物组合之一,主要生活在5.75亿年前至5.41亿年前的海洋中,是宏体生物的演化过程中最为独特的化石生物群,它们为软躯体,化石以印痕或铸型方式保存于碎屑岩中。埃迪卡拉生物化石形态独特,以状、叶状、管状等为特征,其大小不一,从几厘米到超过一米。关于埃迪卡拉生物的分类归属一直争论很大,多数学者认为它们可能是后生动物的早期代表,是后生动物出现后的第一次适应性辐射,其所采取的是与现代大多数动物截然不同的形体结构,是为适应在元古宙末期低氧环境。目前的证据表明,埃迪卡拉生物群的组成十分复杂,但其属性多数存疑,传统观点认为绝大多数的埃迪卡拉生物可以和现生类群对比,如辐射对称和海笔状的类型常归入刺胞类,近于两侧对称的分节类群被归入环节动物和节肢动物[152-153]。由于缺乏可与现代生物类群明确对比的形态特征,加之软躯体化石常在中颗粒砂岩中呈铸型或铸膜保存,对其分类属性的认识目前仍众说纷纭,包括动物进化中“失败的实验品”——文德动物(“Vendobionts”)[154]、原生动物[155]、地衣[156]、原核生物聚合体[157]及真菌[158]等。近年来,较为统一的认识是它既包括一些后生动物的基干或冠群类群,也包括大量已绝灭的生物类群[159-161]。
Waggoner[162]根据埃迪卡拉生物群的演化特征及时空分布,将其划分为3个近于连续的演化生物群组合.:即阿瓦隆组合 (575~559 Ma)、白海组合(558~550 Ma)、纳玛组合 (549~541 Ma),但随后的研究发现核心的化石组合可能代表了不同的环境分异(如对水深的生态响应[163-164])、埋藏学偏差[165] 等,不同组合在时间和生态上甚至都可能存在叠加性[166]。虽然对3个组合的相互关系仍有争议,但必须指出3个化石组合的主要特征还是比较明确的。最早的阿瓦隆组合软躯体生物构型相对简单,以 rangeomorph 和 arboreomorph 类群组成[167-168],代表深水边缘斜坡和盆地相组合类群。这些组合通常以“Conception型”保存在火山灰层之下[165,169],或深水砂岩包卷层和浊积层之内[166]。它的实体构型相对简单,以分形方式,通过模块化在二维平面上增加表面积来生长,通过体表渗透从水体中吸收养分,即通过渗透营养(osmotrophy)方式生活。白海组合(558~550 Ma)主要由Bilateralomorph, Dickinsoniomorph, 和 Kimberellomorph等不同身体构型的生物类群所组成,通常发现在浅海陆架环境中,微生物席占主导的环境中,以“Flinders型”的铸模和铸型保存[164-165,170-171],以出现较为丰富的遗迹化石为特征;最年轻的纳玛组合(549~541 Ma)主要发现于浅的陆架-海岸边缘环境中的,生物身体构型趋于 erniettomorph 和 rangeomorph 分类群落,通常以三维铸型的形式保存在风暴沉积的沙层和河道充填沉積物中,最典型特征是出现大量骨骼生物,如全球性分布的管状骨骼生物Cloudina等[122]及大量尚待深入研究的管状化石(如Carbone等[173]所报道的)。
4.1 阿瓦隆生物组合(575~565 Ma)中的管状化石
Thectardis报道于纽芬兰埃迪卡拉纪Conception 群(575~565 Ma)[174],是一类厘米级的三角形印痕,边缘突出,内部无特征或仅有轻微的横向脊。复原后认为化石为锥管状化石,个体较大(直径最大超过 80 mm,高可达150 mm),根据形态及埋藏学特征推断它是一类食悬浮的‘mat sticker,以其锥顶插入藻席覆盖的沉积物中直立生长。
4.2 白海组合中的管状化石
三类产自南澳Nilpena Rawnsley Quartzite砂岩 Ediacara 段(约570 Ma),原定为遗迹化石的类型,经重新研究厘定,被归入实体管状化石进行了详细的描述,分别定名为Somatohelix sinuosus,Funisia dorothea和Plexus ricei[8,175-176]。有学者因而指出必须对前寒武纪大量原定为遗迹化石的类型进行更为深入的研究,其中许多化石可能是实体管状化石。
Somatohelix sinuosus Sappenfield et al.,2011:为一类砂岩层面上正玄曲线状弯曲的管子,在层面上呈规则对称的波状沟或脊,由至少两个以上管体直径恒定的正弦波所组成。 正弦曲率保持恒定并且与管径大小无关。 横截面为圆形,其直径在整个化石体范围内大小一致。 平行于化石轴线的侧缘整齐而弯曲,没有内部或外部的装饰。Glaessner [177]最初将其作为遗迹化石描述,Sappenfield 等[175]对其重新研究发现Somatohelix sinuosus与围岩界线明显,并且可以完整的从围岩中取出,证明它更可能是一类实体管状化石。进一步观察发现,其整体的形态是松散的螺旋管状。Sappenfield等[175]将其解释为底栖固着的类型,螺旋状的结构形态有利于其生活时保持直立,其底部部分管体与基质接触并埋藏在基质内,上部螺旋状管体竖立于水体中并行可能的滤食生活。
Funisia dorothea Droser and Gehling 2008:是另一类原解释为遗迹化石的管状实体化石。管体长可超过30cm,管体直径在2~12 mm之间,并且在同一层面上同一属种、个体大小一致的管体常聚集在一起,显示聚集生活的特点。管体由模块化的,纵向分成6~8 mm长的系列原盘或串珠状构造单元所组成,各单元由与管轴垂直或倾斜的收缩纹所显现。个体通过在管状体上添加连续的单元和通过管子的分枝来生长。固着构造由直径1~8 mm的层面上内陷的突起所呈现。 该化石最早被认定为动物的粪球粒,但它们的结构和定向紧密排列的特征意味着可能与多孔类或刺胞动物存在亲缘[8]。因此,单个生物体通过在管中添加一列列的模块单元及通过管体的分裂来生长,并通过同时产生大量的雌性和雄性生殖细胞,通过有性繁殖来扩散繁殖。在现生生物中,能同时吐出大量的配子的繁殖方式几乎毫无例外是有性繁殖的结果,但极少与无性繁殖相关。Funisia dorothea被复原为一类用藻席层下的固着器及垂直生长的群居管状生物类型。
Plexus ricei Joel et al.,2014:化石体则由两个部分组成: 一个坚硬的中间管状结构和一个脆弱的外管状壁。化石体长5~80 cm,宽5~20 mm,P. ricei对称的体构显示可能属两侧对称动物,但化石体前后无法确认,因此生物属性不明[176]。从化石描述和化石复原可以初步判断Plexus ricei是由单列的串珠状模块连接而成。
4.3 埃迪卡拉生物群纳玛组合中的管状化石
纳玛组合以出现大量骨骼生物为特征,其中包括具有全球分布的最早骨骼动物化石Cloudina, 同时还有许多尚待描述的管状动物类型,其中Cloudina已在文中作了详细讨论,其他同时期的管状化石的报道分别来自加拿大 Mackenzie山Blueflower组(同位素年龄553~541 Ma)的Sekwitubulus annulatus和Annulatubus flexuosu[173],俄罗斯地台北部Onega 河中游的文德系上统的Vendoconularia triradiata[172],乌拉圭埃迪卡拉系Arroyo del Soldado群中的Waltheria 和Soldadotubulus[18],以及产自俄罗斯中部和西北部地区埃迪卡拉纪晚期至寒武纪早期地层中的管状化石Sabellidites Yanishevsky,1926[178-180]。下面就上述几类管状化石进行较为详细的描述和讨论。
Sekwitubulus annulatus Carbone et al.,2015:为直的,具有一定刚性的小型圆柱形的管(管径通常小于5 mm),管壁薄,管表面装饰有均匀分布的环脊,脊间距约1mm。通过与管直径相同的固着盘附着在基底上生活。以上特征表明Sekwitubulus很可能是一类垂直生长的管,并通过一个小的固着器固定在基质上。在形态结构上与现代多毛类蠕虫管相似,Sekwitubulus类似于管状多毛类蠕虫的管子,因为它有坚硬的管状外壳,光滑的管腔,具外部环状脊(可能是生长环或口缘),缺少锥度[114]。 不管化石大小有何差异,环脊的厚度保持恒定,说明Sekwitubulus可能是通过在末端增加环来生长的,这也类似于多毛类蠕虫的管子。然而,这种结构上的相似性并不意味着Sekwitubulus和现代多毛类蠕虫之间存在亲缘关系[173]。
Annulatubus flexuosus Carbone et al.,2015:是波曲状,厘米级的圆柱形管,表面具横向或稍呈新月形的环状脊,脊间距均匀分布,约5 mm,具新月形而不是横向的环突证明该化石具一定柔韧性,易于压缩的,未见固着构造。 Annulatubus在大小上接近于Wutubus annularis,但后者为锥形的管[69], Annulatubus直径恒定,具环纹及呈波曲状的特征与Grazhdankin等 (95-2008)描述的产自西伯利亚北部的有规则间隔的环纹物极为相似,但后者呈炭质膜状保存[173]。
Vendoconularia triradiata Ivantsov and Fedonkin,2002:化石产自俄罗斯地台北部Onega 河中游的文德系上统Ust′-Pinega组下部碎屑岩中,共生的化石包括曾在纳玛群中发现的铸型保存的软躯体化石Swartpuntia Narbonnes,地方性属种Ventogyrus Ivantsov and Grazhdank及类似庙河生物群中的Calyptrina状管状化石,以及许多尚未描述的化石[172]。
Vendoconularia为细长锥管状,具有6个相同的面,每个面有两排交错的横脊。有纵脊沿面与面间的纵向角沟分布。其总体锥管状的形态,放射状排列的由两侧分布的横脊组成的面,特征都与锥石类相似。不同仅在于目前标本可能是有机质的,而非磷酸盐化的体壁,总体呈现六辐或一般为三辐射对称(锥石一般为四辐对称)。
Waltheria 和Soldadotubulus:在乌拉圭埃迪卡拉系Arroyo del Soldado群Yerbal組的条带状粉砂岩中发现了多种类型的管状和锥管状化石,除比较常见的Cloudina riemkeae,还有较为特殊的两类管状化石,Waltheria marburgensis Gaucher and Sprechmann, 1999和Soldadotubulus siderophoba Gaucher and Sprechmann,1999。
Waltheria marburgensis Gaucher and Sprechmann, 1999:为直管状,两头尖缩,管内具隔壁状构造,在隔壁处出现管径收缩。管体各分节长度不均一。管壁具三层式结构:内层隔壁层;中间层具螺旋结构;外层通常被石英交代。 管体一端为基部圆柱形凸起的球形腔,另一端可能开口。管体为一系列具组织结构的亚球形单元形成的单列构型,但不乏有多束排列的现象。管体常见分枝。可能代表繁殖现象。管体直径变化很大,10 μm~1 mm,长度变化在0.8~8 mm。原始成分可能为磷酸钙质。隔壁没有穿孔以及多层管壁排除其为有孔虫类可能,从其较为复杂的结构构造,作者推测为原始后生动物[18]。
Soldadotubulus:为直或微弯,不分枝但分节的管,向两端尖缩。口部直径大于管径的圆筒状,在管体中部或甚至下部还会出现一类似的圆筒状结构;管体具规则的分节,在分节处管体有明显收缩现象,管体表明常见垂直与长轴发现的脊状纹饰。管体内径30~67 μm。管长0.2~0.6 mm。Soldadotubulus与Waltheria都为分节的管状构造,可能存在一定亲缘[18]。
Corumbella werneri:Corumbella仅局限分布于亚马逊(Amazonas)和旧金山(San Francisco)克拉通,包括巴西、巴拉圭[122,181]及劳伦西亚大陆,如,美国、墨西哥等地[19,182]。Hahn 等[181]和Walde等[183]重建的Corumbella werneri为圆形的管,具有近端‘柄区和远端的‘头区,具有双列排列的‘息肉状构造。这一解释表明它可能与刺胞类钵水母纲的冠状动物(scyphozoan coronates),并与埃迪卡拉生物群中典型的charniid类化石具有一定可比性[181-183]。而Babcock等[184]则推断Corumbella werneri具有四角形横截面的管,不存在近端和远端的区分;可能与锥石类关系紧密,并与刺胞类钵水母纲具亲缘关系。Pacheco[185] 在其博士论文中对Corumbella werneri进行了较为详细的研究,重建的管状体中线具四方截面,内部具横隔板及与锥石类似的具有乳突和小孔的有机质多边形板片,认为属真正的刺胞动物锥石类[122,185-187]。虽然,Erdtmann 等[188-190]注意到扁平的C. werneri管与cloudinids一样可能属具软躯体的埃迪卡拉动物[187],Walde等[191]的最新研究否定了Pacheco 等[186]所描述的管体四方截面特征及内部横隔板的存在,柱状而非锥管状,圆或椭圆形横截面,未见附着构造,可以排除其为刺胞类锥石的可能,认为C. werneri可能属早期环节动物或类似的具部分矿化(可能是文石质),或完全有机质的体外管,尤其他表面特征的不规则状皱褶及二分叉的横脊,与Sinotubulites baimatuensis的特征完全一致。笔者对C. werneri和高家山生物群中发现的磷酸盐化Sinotubulites观察对比支持Walde等[191]的观点,认为二者可能存在一定亲缘。
Sabellidites Yanishevsky,1926(皱节虫):是一类产自埃迪卡拉纪晚期至寒武纪早期地层中的管状化石,目前关于该化石的研究报道主要见于俄罗斯中部和西北部地区[178-180]。Sabellidites化石呈笔直条带状,宽度范围在0.2~3.0 mm,最大长度超过16 cm。表面具有密集的横纹,横纹或互相平行,或以低角度斜交[180]。不同于其他晚埃迪卡拉纪管状化石的保存模式(例如钙化、磷酸盐化、黄铁矿化和黏土矿物交代),Sabellidites通常呈有机质形式保存[180]。Yanishevsky[178]推测Sabellidites成分可能为几丁质[180],同时依据形态特征将Sabellidites归为须腕动物门(现等同于环节动物门多毛纲西伯加虫科(Siboglinidae Caullery, 1914)[192])。也有不同观点认为,Sabellidites应归为分类位置不明的化石类群[179]。Moczydlowska等[180]发现Sabellidites中有机质管的微细结构是由多层的,镶嵌在有机基质中,不同排列方向的纤维组成的离散层。高度有序和特殊的纤维排列模式证实,Sabellidites与现生深海环节动物Siboglinids具有较近的亲缘关系。
5 埃迪卡拉纪管状动物化石的时空分布
通过对新元古代管状化石记录的系统梳理,我们发现管状生物在埃迪卡拉纪存在两个明显的演化阶段:
1)管状化石的萌发阶段(600~551 Ma),其时限约等于埃迪卡拉纪陡山沱期。新元古代大冰期结束后不久,最早可能的管状化石就已经出现了,它始现于较深水陆棚相的黑色泥岩中,以炭质压膜保存,主要发现于陡山沱期蓝田生物群、庙河生物群中。在同为深水相的埃迪卡拉生物群阿瓦隆组合中也出现了少量可疑的以铸型或铸膜保存的类型。其主要特点:化石主要为锥管状,细长圆柱管状为主,类型单调,多数具有固着构造,部分可能具触手,但缺乏明显的管体纹饰;分布十分局限,仅在中国华南及加拿大纽芬兰个别产地有报道,并且多数为地方性属种。近年来在澳大利亚南部浅海相的Rawnsley Quartzite砂岩中,在埃迪卡拉生物群白海组合中发现了两类较为复杂的类型,Funisia dorothea及Plexus ricei,它们均呈细长管状,管体具有并与典型埃迪卡拉软躯体化石共生,但目前仅在一个产地发现。由于湖北三峡地区庙河生物群[53],贵州江口桃映庙河生物群[193] 和澳大利亚南部Rawnsley Quartzite砂岩中[194]都出现了被解释为栉水母类的干群类型的Eoandromeda octobrachiata[194-195],表明廟河生物群与澳大利亚南部的埃迪卡拉生物群在时代相同。
2)管状化石快速辐射演化期(551~541Ma),其时限基本相当于埃迪卡拉纪灯影组沉积期,与埃迪卡拉生物群纳玛组合的时限基本一致,对应华南地区两个著名的生物群,石板滩生物群和高家山生物群。该时期管状动物化石的特点是:生物类型多样,除最早管状骨骼矿化生物Cloudina外,还有Shaanxilithus,Conotubus,Gaojiashania,Sinotubulites,Corumbella,Sabellidites等,更出现了许多尚未描述并与早寒武世小壳化石有一定亲缘的微体管状类型,同时还出现大量地方性属种,如Sekwitubulus annulatus,Annulatubus flexuosu,Vendoconularia triradiata,Waltheria 和Soldadotubulus等;其形态复杂,它可能呈圆管状(Gaojiashania,Sekwitubulus,Annulatubus)或棱管状(Corumbella),可能不具任何纹饰,或具有环纹或皱褶 (如Cloudina,Corumbella 和 Sinotubulites),或为管套管模式(Sinotubulites),或为锥套锥构型(Conotubus 和Cloudina),有些内部中空(如Cloudina和Sinotubulites),有些具横隔壁(如Waltheria),还有少量分枝类型(Cloudina 和 Waltheria);管壁成分多样,大部分为有机质壁管,但有些可能具矿化或弱矿化的壁 (Cloudina, Sinotubulites 和 Waltheria);分布更趋广泛,其中的Sinotubulites,Cloudina和Shaanxilithes等在世界许多地区埃迪卡拉纪晚期地层均有发现[76,140,149], 显示出它们具有成为区域和洲际埃迪卡拉纪晚期地层对比标准化石的潜力[20]。近来,一些原来的地方性属种也逐步扩大了其分布范围,如原仅见于华南的Wutubus, Gaojiashania, Conotubus等在美国内华达、纳米比亚等都有发现。上述管状动物可能具有不同的起源,但共同组成了埃迪卡拉纪复杂多样的生物群面貌[28]。
6 埃迪卡拉纪管状动物化石构型演化
根据管状化石形态差异、管体结构构型等,埃迪卡拉纪末期管状化石可以分为以下几种形态类群:
1) 简单管状或锥管状;为锥管状,细长圆柱管状管体,多数具有固着构造,缺乏明显的管体纹饰,个别具触手构造。包括蓝田生物群的Lantianella laevis,L. annularis, Piyuania cyathiformis, Qianchuania fusiformis. 和 Xiuningella rara;庙河生物群中的Calyptrina striata和Protoconites minor,以及埃迪卡拉生物群阿瓦隆组合中的Thectardis,白海组合中的Somatohelix sinuosus等。
2) 简单模块化构型;由形态、结构基本一致的构造单元通过单列(或偶为双列)模块化组成。如Shaanxilithes是由一系列特征一致的圆或月牙形构造组合而成的特殊生物类群,在后期化石化过程中经常发生压扁、分离,从而保存为不同的形态特征。而著名的Funisia dorothea, 管体由模块化的,一系列圆盘或串珠状构造单元通过在管状一端不断添加新的模块单元或通过管子的无性分枝来生长。
3) 有一定形态分异的复杂模块化构型;有一系列具有不同结构单元或一定形态分异的模块组合而成的管状或锥管状构造,如 Gaojiashania类直管状构型,管体由一系列圆环组成,圆环由软组织连接而成,两个圆环之间的软组织发育有3—5条平行于圆环的环纹;cloudinids类锥管状构型,包括Conotubus,管體由一系列套叠的桶状管壁构成,管壁成分可能是有机质或弱矿化,Cloudina的漏斗状套合,具矿化壁; sinotubulitid类管套管状多层管构型,多层套管结构;内部管壁光滑,外部管体褶皱强烈;管体横截面可以看到具3-,5-,6-,8-辐射对称特征。
4) 与早寒武世管状动物化石可能存在亲缘关系的管构型:构型虽趋于简单,但表现在矿化管壁结构的复杂化。
推测的埃迪卡拉纪管状动物构型演化模式(但不代表生物系统进化关系)见图18。
7 结 论
通过对新元古代末期已有化石资料的系统梳理,基于对高家山生物群的较为系统的研究,初步认为在寒武纪大爆发之前、与软躯体埃迪卡拉生物群几乎同期,存在着管状动物的大辐射,其形态复杂,生物构型多样,个体大小差异悬殊,生活方式繁杂,多数类群与现生生物类群对比困难,代表了地球生命史上一次重大的生物革新事件。这次大辐射的特点可以总结为以下几个方面:
生物类群多样化。从埃迪卡拉纪早期仅有少量地方性属种,至埃迪卡拉纪末期化石属种已超过30余种,并且从原来基本是一属单种发展到一属多种,如Cloudina,目前已见诸报道的属种有9个。
管体构型复杂化。埃迪卡拉纪的管状动物构型存在着从简单的圆柱形、锥形,到简单的模块化重复单元建造发展到有不同分异的模块化单元建造,最终进化到矿化结构单元的演化。同时同一类型化石其结构液呈现多样化的趋势,如Cloudina,呈现出套合疏密性极大差异,壳面纹饰繁杂,有光滑,上部具纹饰,整个小锥体具环纹,纵向具皱褶;对于Sinotubulites,更呈现出较为复杂的对称性,3-,5-,6-,8-辐射对称。
新的生态方式产生。管状动物已从单一的底栖固着食悬浮,至埃迪卡拉纪末期,呈现底栖固着食悬浮、底栖游移食碎屑(Gaojiashania),底栖平躺固着食碎屑(Sinotubulites)、小型礁体等丰富多样的生活方式。
生殖方式多样化。在同一个化石类型中出现了有性繁殖与无性繁殖有性交替的现象。有性繁殖可能便于快速繁殖和扩散、无性繁殖有利于形成小型礁体和抵御环境变化。
同时在进化过程中伴随着分布范围的逐步扩大,广布性属种逐步增多,至埃迪卡拉纪末期以Cloudina为代表的骨骼管状动物已在全球20余个化石产地发现,成为新元古代末期最好的标志性化石。
致谢:感谢审稿专家和编辑部在本文修改过程中提出的建设性意见和建议!
参考文献:
[1]ZHURAVLEV A Y, LIN E, VINTANED J A G, et al. New finds of skeletal fossils in the terminal neoproterozoic of the Siberian platform and Spain [J].Acta Palaeontologica Polonica,2012,57(1):205-224.
[2]AMTHOR J E, GROTZINGER J P, SCHRDER S,et al.Extinction of Cloudina and Namacalathus at the Precambrian-Cambrian boundary in Oman [J].Geology,2003,31(5):431-434.
[3]KOUCHINSKY A V. Skeletal microstructures of hyoliths from the early Cambrian of Siberia[J].Alcheringia:an Australasian Journal of Palaeontology,2000,24(2):65-81.
[4]KOUCHINSKY A V, BENGTSON S. The tube wall of Cambrian anabaritids[J].Acta Palaeontologica Polonica,2002,47:431-444.
[5]MALINKY J M, SKOVSTED C B. Hyoliths and small shelly fossils from the Lower Cambrian of northeast Greenland [J].Acta Palaentologica Polonica,2004,49(4):551-578.
[6]SKOVSTED C B, PEEL J S.Hyolithellus in life position from the Lower Cambrian of North Greenland [J].Journal of Paleontology,2011,85,1:37-47.
[7]JENSEN S.The Proterozoic and earliest Cambrian trace fossil record; patterns, problems and perspectives[J].Integr Comp Biol,2003, 43:219-228.
[8]DROSER M L, GEHLING J G.Synchronous aggregate growth in an abundant new Ediacaran tubular organism[J].Science,2008,319(5870):1660-1662.
[9]COHEN P A, BRADLEY A, KNOLL A H, et al.Tubular compression fossils from the Ediacaran Nama Group, Namibia[J].J Paleontology, 2009,83(1):110-122.
[10]SIGNOR P W, MCMENAMIN M A S. The early Cambrian worm tube onuphionella from California and Nevada[J].Journal of Paleontology,1988,62(2):233-240.
[11]YANG B, STEINER M,ZHU M Y, et al. Transitional Ediacaran-Cambrian small skeletal fossil assemblages from South China and Kazakhstan: Implications for chronostratigraphy and metazoan evolution[J].Precambrian Research,2016,285:202-215.
[12]ZHU M, ZHURAVLEV A Y, WOOD R A, et al. A deep root for the Cambrian explosion: Implications of new bio-and chemostratigraphy from the Siberian Platform[J].Geology,2017,45(5):459-462. doi:10.1130/G38865.1.
[13]CAI Y P, XIAO S H, LI G, et al. Diverse biomineralizing animals in the terminal Ediacaran Period herald the Cambrian explosion[J]. Geology, 2019,47(4):380-384.
[14]GRANT S W F.Shell structure and distribution of Cloudina, a potential index fossil for the Terminal Proterozoic[J]. Am J Sci,1990,290-A:261-294.
[15]XIAO S. YUAN X, KNOLL A H. Eumetazoan fossils in terminal Proterozoic phosphorites?[J].Proceedings of the National Academy of Sciences of the United States of America,2000, 97(25):13684-13689.
[16]XIAO S H, YUAN X L, STEINER M, et al. Macroscopic carbonaceous compressions in a terminal Proterozoic shale: A systematic reassessment of the Miaohe biota, South China[J].Journal of Paleontology,2002,76(2):347-376.
[17]DONG L, XIAO S H, SHEN B,et al. Silicified Horodyskia and Palaeopascichnus from Upper Ediacaran cherts in South China: Tentative phylogenetic interpretation and implications for evolutionary stasis[J]. Journal of the Geological Society,2008,165(1):367-378.
[18]GAUCHER C, SPRECHMANN P. Upper Vendian skeletal fauna of the Arroyo del Soldado Group, Uruguay[J].Beringeria, 1999, 23:55-91.
[19]HAGADORN J W, WAGGONER B M. Ediacaran fossils from the southwestern Great Basin, United States[J]. J Paleontol,2000,74(2):349-359.
[20]HUA H, CHEN Z, YUAN X L.The advent of mineralized skeletons in Neoproterozoic Metazoa:New fossil evidence from the Gaojiashan Fauna[J].Geol J,2007,42(3/4):263-279.
[21]DROSER M L, GEHLING J G, JENSEN S R. Assemblage palaeoecology of the Ediacara biota: the unabridged edition?[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2006,232(2/3/4):131-147.
[22]GERMS G J B.New shelly fossils from the Nama Group, south west Africa[J].Am J Sci, 1972, 272(8):752-761.
[23]陈孟莪,陈忆元,钱逸.峡东区震旦系-寒武系底部的管状动物化石[J]. 中国地质科学院天津地质矿产研究所所刊, 1981, 3:117-124.
[24]HUA H, CHEN Z, YUAN X L, et al.Skeletogenesis and asexual reproduction in the earliest biomineralizing animal Cloudina[J].Geology, 2005,33(4):277-280.
[25]HUA H, PRATT B R, ZHANG L Y. Borings in cloudina shells: Complex predator-prey dynamics in the terminal neoproterozoic[J]. Palaios, 2003,18(4/5):454-459.
[26]SOKOLOV B S.Vendian and Early Cambrian Sabelliditida (Pogonophora) of the USSR[C]∥Proceedings, 23rd International Geological Congress, Prague, 1968:79-96.
[27]XIAO S H, DONG L. On the morphological and ecological history of Proterozoic macroalgae[M]∥Topics in Geobiology. Dordrecht Netherlands:Springer,2006:57-90.
[28]LIU P J, XIAO S H, YIN C Y, et al. Systematic description and phylogenetic affinity of tubular microfossils from the Ediacaran Doushantuo Formation at Weng′an, South China[J].Palaeontology, 2008, 51(2):339-366.
[29]DONG L, XIAO S H, SHEN B,et al.Restudy of the worm-like carbonaceous compression fossils Protoarenicola, Pararenicola, and Sinosabellidites from early Neoproterozoic successions in North China[J].Palaeogeography, Palaeoclimatology,Palaeoecology,2008,258(3):138-161.
[30]COHEN P A, BRADLEY A, KNOLL A H, et al.Tubular compression fossils from the Ediacaran Nama Group, Namibia[J].Journal of Paleontology,2009,83(1):110-122.
[31]牛绍武,辛后田,刘欢,等.论中国拉伸纪淮南系的建立[J].地质调查与研究,2018,41(4):241-257.
NIU S W, XIN H T, LIU H, et al. Establishment of the Huainan System (Tonian Period) in Northen China platform[J].Geological Survey and Research,2018,41(4):241-257.
[32]汪贵祥.安徽淮南晚期前寒武纪环节和须腕动物化石[J].中国地质科学院天津地质矿产研究所所刊,1983(6):9-22.
[33]SUN W G, WANG G X, ZHOU B H. Macroscopic worm-like body fossils from the upper Precambrian(700~900 Ma), Huainan district,Anhui, China and their stratigraphyic and evolutionary significance[J].Precambrian Research, 1986,31(4):377-403.
[34]CHEN J. Precambrian metazoans of the Huai River drainage area (Anhui, E. China): Their taphonomic and ecological evidence[J].Senkenbergiana Lethaea,1988,69(3/4):189-215.
[35]鄭文武. 皖北震旦系中Chuaria等化石的发现及其地质意义[J].中国地质科学院天津地质矿产研究所分刊,1980,1(1): 49-69.
[36]KUMAR S. Mesoproterozoic megafossil Chuaria-Tawuia association may represent parts of a multicellular plant, Vindhyan Supergroup, Central India[J].Precambrian Res,2001,106(3/4):187-211.
[37]叶琴. 华南神农架-黄陵地区晚新元古代宏体藻类化石研究[D].武汉:中国地质大学(武汉),2018.
[38]WAN B, YUAN X L, CHEN Z, et al.Systematic description of putative animal fossils from the early Ediacaran Lantian Formation of South China[J].Palaeontology,2016, 59(4):515-532.
[39]YUAN X L, CHEN Z, XIAO S H,et al. An early Ediacaran assemblage of macroscopic and morphologically differentiated eukaryotes[J].Nature, 2011,470(7334):390-393.
[40]YUAN X L, LI J, CAO R J. A diverse metaphyte assemblage from the Neoproterozoic black shales of South China[J].Lethaia,2007,32(2):143-155.
[41]WAN B, YUAN X, CHEN Z,et al. Quantitative analysis of Flabellophyton from the Ediacaran Lantian Biota, South China: Application of geometric morphometrics in Precambrian fossil research[J]. Acta Geologica Sinica (English Edition),2013,87(4):905-915.
[42]XIAO S H, DROSER M, GEHLING J G,et al.Affirming life aquatic for the Ediacara biota in China and Australia[J].Geology, 2013, 41(10):1095-1098.
[43]LIU P J, XIAO S H, YIN C Y,et al.Silicified tubular microfossils from the upper Doushantuo Formation (Ediacaran) in the Yangtze Gorges area, South China[J].Journal of Paleontology,2009,83(4):630-633.
[44]ZHOU C M, LI X, XIAO S H, et al. A new SIMS zircon U-Pb date from the Ediacaran Doushantuo Formation: Age constraint on the Weng′an biota[J]. Geological Magazine, 2017,154(6):1193-1201.
[45]薛耀松,唐天福,俞從流.贵州晚震旦世陡山沱期具骨骼动物化石的发现及其意义[J]. 古生物学报,1992,31(5):530-539.
XUE Y S,TANG T F,YU C L.Discovery of oldest skeletac fossics from upper sinian Donshantuo Formation in Weng′an Guizhou,and its significance[J].Acta Palaeontologica Sinica,1992,31(5):530-539.
[46]LI G X,XUE Y S,ZHOU C M.Late Proterozoic tubular fossils from the Doushantuo Formation of Weng′an, Guizhou, China[J].Palaeoworld, 1997,7:29-37.
[47]CHEN J Y, OLIVERI P, GAO F et al. Precambrian animal life: Probable developmental and adult cnidarian forms from southwest China[J].Developmental Biology, 2002, 248(1):182-196.
[48]LIU P J, YIN C Y, TANG F.Microtubular metazoan fossils with multi-branches in Weng′an Biota[J].Chinese Science Bulletin, 2006, 51(5):630-632.
[49]刘鹏举,尹崇玉,陈寿铭,等.华南埃迪卡拉纪陡山沱期管状微体化石分布、生物属性及其地层学意义[J].古生物学报,2010, 49(3):308-324.
LIU P J,YIN C Y,CHEN S M,et al. Affinity,distribution and stratigraphic significance of tubular microfossils from the Ediacaran Doushantuo Formation in South China[J].Acta Palaeontologica Sinica,2010,49(3):308-324.
[50]CUNNINGHAM J A, VARGAS K, LIU P J,et al.Critical appraisal of tubular putative eumetazoans from the Ediacaran Weng′an Doushantuo biota[J].Proceedings of the Royal Society B:Biological Sciences, 2015,282(812):20151169.
[51]陈孟莪,肖宗正.峡东区上震旦统陡山沱组发现宏体化石[J].地质科学,1991,26(4):317-324,407-412.
CHEN M,XIAO Z. Discovery of the macrofossils in the Upper Sinian Doushantuo Formation at Miaohe, eastern yangtze gorges[J].Chinese Journal of Geology(Scientia Geologica Sinica),1991(4):317-324.
[52]陳孟莪,肖宗正,袁训来.晚震旦世的特种生物群落——庙河生物群新知[J]. 古生物学报,1994,33(4):319-403,525-528.
CHEN M,XIAO Z,YUAN X,et al.A new assemblage of megafossics:Miaohe Biota from upper sinian Doushantuo Formation,Yangtze Gorges[J].Acta Palaeontologica Sinica,1994,33(4):319-403,525-528.
[53]丁莲芳,李勇,胡夏嵩,等. 震旦纪庙河生物[M].北京:地质出版社,1996.
[54]STEINER M.Die neoproterozoishen Megaalgen SudChinas [J].Berliner Geowissrnschaftliche, Abhandlungen,1994,15(E): 1-146(in German with English abstract).
[55]唐烽,尹崇玉,刘鹏举,等.华南伊迪卡拉纪“庙河生物群”的属性分析[J].地质学报,2008,82(5):601-610.
TANG F,YIN C Y,LIU P J,et al.Morphological Comparison of the Ediacaran Miaohe Biota from South China: Implications for their affinities and ecology[J].Acta Geologica Sinica,2008,82(5):601-610.
[56]陈孟莪, 萧宗正.峡东震旦系陡山沱组宏体生物群[J].古生物学报, 1992, 31 (5):513-529.
CHEN M,XIAO Z.Macrofossil Biota from Upper Sinian Doushantuo Formation in eastern Yangtze Gorges,China[J].Acta Palaeontologica Sinica,1992,31(5):513-529.
[57]CHEN J, ZHOU G, ZHU M Y, et al.The Chengjiang Biota:A Unique Window of the Cambrian Explosion[M].Taichung:National Museum of Natural History,1996:1-222.
[58]CHEN J, ZHOU G. Biology of the Chengjiang fauna[J]. Bulletin of the National Museum of Natural Science (Taipei), 1997(10):11-105.
[59]袁训来, 肖书海, 尹磊明,等.陡山沱期生物群——早期动物辐射前夕的生命[M].合肥:中国科学技术出版社, 2002:1-171.
[60]杨兴莲, 赵元龙, 朱茂炎.贵州下寒武统牛蹄塘生物群中海绵新材料[J].古生物学报,2005, 44(3):454-463.
YANG X L,ZHAO Y L,ZHU M Y.New sponges from the lower cambrian of Guizhou[J].Acta Palaeontologica Sinica,2005,44(3):454-463.
[61]BRIGGS D E, ERWIN D H, COLLIER F J. The Fossils of the Burgess Shale[M].Washington DC:Smithsonian Institution Press,1994.
[62]SOKOLOV B S. Drevneyshiye pogonofory [The oldest Pogonophora][J].Doklady Akademii Nauk SSSR,1967,177(1):201-204 (English translation page 252-255).
[63]SOKOLOV B S. Essays on the Advent of the Vendian System[M].Moscow:KMK Scientific Press, 1997.
[64]WERNER B. Stephanoscyphus (Scyphozoa, Coronatae) und seine direkte Abstammung von den fossilen Conulata[J].Helgolnder Wissenschaftliche Meeresuntersuchungen, 1966,13(4):317-347.
[65]FEDONKIN M A. Paleoichnology of Vendian Metazoa[M].Heidelberg:Springer-Verlag,1990.
[66]王约, 王训练, 黄禹铭. 华南伊迪卡拉系陡山沱组Protoconites的分类位置与生态初探[J]. 地球科学,2007,32 (增刊):1-10.
[67]郭俊锋, 李勇, 韩健,等.原锥虫属(Protoconites Chen et al., 1994)在湖北三峡地区纽芬兰统(Terreneuvian)岩家河组的发现[J].自然科学进展,2009,19(2):180-184.
[68]CONWAY MORRIS S, ROBISON R A.More soft-bodied animals and algae from the Middle Cambrian of Utah and British Columbia[J].The University of Kansas Paleontological Contributions,1988,122:23-84.
[69]CHEN Z, ZHOU C M, XIAO S H, et al.New Ediacara fossils preserved in marine limestone and their ecological implications[J].Scientific Reports,2015,4:4180.
[70]CHEN Z, ZHOU C M, YUAN X L,et al. Death march of a segmented and trilobate bilaterian elucidates early animal evolution[J]. Nature, 2019,573(7774):412-415.
[71]SMITH E F, NELSON L L, STRANGE M A, et al.The end of the Ediacaran: Two new exceptionally preserved body fossil assemblages from Mount Dunfee, Nevada, USA[J].Geology,2016,44(1):911-914.
[72]华洪.5.5 亿年前动物“临终遗迹”的发现将分节动物的祖先推前了一千万年[J]. 科学通报,2019,64(36): 3781-3783.
HUA H.550 Ma-old body fossils and their mortichnium traces rolls back the origins of segemented animals[J].Chinese Science Bulletin,2019,64(36):3781-3783.
[73]陈孟莪,陈祥高,劳秋元. 陕南震旦系上部地层中的后生动物化石及其地层意义[J].地质科学,1975,10(2):181-190,197-198.
[74]ZHURAVLEV A Y, VINTANED J A G, IVANTSOV A Y.First finds of problematic Ediacaran fossil Gaojiashania in Siberia and its origin[J].Geological Magazine,2009,146(5):775-780.
[75]MEYER M, SCHIFFBAUER J D, XIAO S H,et al.Taphonomy of the Upper Ediacaran enigmatic ribbonlike fossil Shaanxilithes[J]. Palaios, 2012, 27(5):354-372.
[76]TARHAN L, HUGHES N, MYROW P,et al. Precambrian-Cambrian boundary interval occurrence and form of the enigmatic tubular body fossil Shaanxilithes ningqiangensis from the Lesser Himalaya of India[J].Palaeontology,2014,57(2):283-298.
[77]張志亮,华洪,张志飞.埃迪卡拉纪疑难化石Shaanxilithes在云南王家湾剖面的发现及地层意义[J].古生物学报,2015,54(1):12-28.
ZHANG Z L,HUA H,ZHANG Z F.Problematic Ediacaran fossil Shaanxilithes from the Jiucheng member of Wangjiawan Section in Jinning,Yunnan Province[J].Acta Palaeontologica Sinica,2015,54(1):12-28.
[78]王欣.晚埃迪卡拉世管状化石陕西迹的形态学、生物地层学及埋藏学研究[D].西安:西北大学,2019.
[79]罗惠麟,蒋志文,武希彻,等.云南东部震旦系-寒武系界线[M].昆明:云南人民出版社,1982:1-200.
[80]邢裕盛,岳昭.中国震旦系-寒武系界线专号[J].中国地质科学院地质研究所所刊,1984,10:111-125.
[81]林世敏,张运芬,陶喜森,等.陕南震旦系上统高家山组发现的后生动物、遗迹化石和宏观藻类化石[J].陕西地质,1986,4(1):9-17,103-104.
[82]张录易.陕西宁强晚震旦世晚期高家山生物群的发现和初步研究[J].西安地质矿产研究所所刊,1986,13:67-88.
[83]丁莲芳,张录易,李勇,等.扬子地台北缘晚震旦世-早寒武世早期生物群研究[M].北京:科学技术出版社,1992:1-156.
[84]李日辉,杨式浦,李维群.中国震旦系-寒武系界线过渡层遗迹化石研究[M].北京:地质出版社,1997:1-99.
[85]WEBER B, STEINER M, ZHU M Y. Precambrian-Cambrian trace fossils from theYangtze Platform (South China) and the early evolution of bilaterian lifestyles[J].Palaeogeography, Palaeoclimatology, Palaeoecology,2007,254(1/2): 328-349.
[86]ZHURAVLEV A Y, VINTANED J A G, IVANTSOV A Y. Discussion of ′First finds of problematic Ediacaran fossil Gaojiashania in Siberia and its origin′ (Reply)[J].Geological Magazine,2011,148(5):329-333.
[87]华洪,陈哲,张录易.Shaanxilithes在贵州的发现及其意义[J].地层学杂志,2004,28(3):265-269,291.
HUA H,CHEN Z,ZHANG L Y. Shaanxilithes from taozichong formation of Guizhou Province and its significance[J].Journal of Stratigraphy,2004,28(3):265-269,291.
[88]SHEN B, XIAO S H, DONG L,et al. Problematic macrofossils from ediacaran successions in the North China and Chaidam blocks: Implications for their evolutionary roots and biostratigraphic significance[J]. Journal of Paleontology, 2007, 81(6): 1396-1411.
[89]DARROCH S A F, BOAG T H, RACICOT R A, et al.A mixed Ediacaran-metazoan assemblage from the Zaris Sub-basin, Namibia[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2016, 459(3): 198-208.
[90]赵银胜.鄂东南的震旦系及其微古植物[J].湖北地质,1995,9(1):21-30,108.
[91]顾鹏,钟玲,张国栋,等.华南埃迪卡拉(震旦)系顶部地层划分及与寒武系界线FAD分子的选择[J]. 地质学报,2018,92(3):449-465.
GU P,ZHONG L,ZHANG G D,et al. The division of the late Ediacaran-Cambrian boundary interval stratigraphy and new options of index fossil FAD in South China[J].Acta Geologica Sinica,2018,92(3):449-465.
[92]王云山,庄庆兴,史从彦,等.柴达木盆地北缘的全吉群[M].天津:天津科学技术出版社,1980:214-230.
[93]杨式傅,郑昭昌.寧夏贺兰山震旦纪正目关组遗迹化石[J].地球科学,1985,10:9-18.
[94]YANG J, ZENG Z, CAI X, et al.Carbon and oxygen isotopes analyses for the Sinian carbonates in the Helan Mountain, North China[J]. Chinese Science Bulletin, 2013, 58(32): 3943-3955.
[95]GRAZHDANKIN D V, BALTHASAR U, NAGOVITSIN K E, et al.Carbonate-hosted Avalon-type fossils in arctic Siberia[J].Geology, 2008, 36(10):803-806.
[96]WOOD R A, ZHURAVLEV A Y, SUKHOV S S, et al.Demise of Ediacaran dolomitic seas marks widespread biomineralization on the Siberian Platform[J].Geology,2017,45(1):27-30.
[97]张录易,董军社,田淑华,等,高家山生物群[M]∥丁莲芳,张录易,李勇,等.扬子地台北缘晚震旦世—早寒武世生物群研究.北京:科学技术文献出版社,1992:33-63.
[98]CAI Y, HUA H, ZHANG X. Tube construction and life mode of the late Ediacaran tubular fossil Gaojiashania cyclus from the Gaojiashan Lagersttte[J].Precambr Res,2013,224:255-267.
[99]陈哲,孙卫国,华洪.陕南晚震旦世Gaojiashania的保存特征及形态解释[J].古生物学报,2002,41(3):448-454.
CHEN Z, SUN W G, HUA H.Preservation and morphol ogic interpretati on of late sinian gaojiashania from southern shaanxi[J].Acta Palaeontologica Sinica,2002,41(3):448-454.
[100]HALANYCH K M. Molecular phylogeny of siboglinid annelids (a.k.a. pogonophorans):A review[M]∥Developments in Hydrobiology.Brlin/Heidelberg:Springer-Verlag,2005:297-307.
[101]SOUTHWARD E C, SCHULZE A, GARDINER S L.Pogonophora (Annelida): form and function[M]∥Developments in Hydrobiology.Brlin/Heidelberg:Springer-Verlag,2005:227-251.
[102]GROTZINGER J P, BOWRING S A, SAYLOR B Z, et al.Biostratigraphic and geochronologic constraints on early animal evolution[J].Science,1995,270:598-604.
[103]CUI H, KAUFMAN A J, XIAO S, et al.Environmental context for the terminal Ediacaran biomineralization of animals[J].Geobiology,2006,14:344-63. doi: 10.1111/gbi.12178.
[104]CAI Y P, SCHIFFBAUER J D, HUA H,et al.Morphology and paleoecology of the late Ediacaran tubular fossil Conotubus hemiannulatus from the Gaojiashan Lagersttte of southern Shaanxi Province, South China[J].Precambrian Research,2011,191(1/2):46-57.
[105]BRUSCA R C, BRUSCA G J. Invertebrates, 2nd ed[M].Sunderland, MA:Sinauer Associates, Inc., 2003.
[106]華洪,陈哲,蔡耀平,等.最早骨骼生物Cloudina——埃迪卡拉纪晚期一类重要的标准化石[J].西北大学学报(自然科学版),2009,39(3):522-527.
HUA H,CHEN Z, CAI Y P,et al.The earliest biomineralizing animal Cloudina:An important index fossil in the Late Ediacaran[J]. Journal fo Northwest University(Natural Science Edition),2009,39(3):522-527.
[107]YOCHELSON L, HERRERA H E. Un fossil enigmatico del Cambrico Inferior de Argentina[J]. Ameghiniana, 1974, 11: 283-294.
[108]GLAESSNER M F. Early phanerozoic annelid worms and their geological and biological significance[J].J Geol Soc,1976,132(3):259-275.
[109]HAHN G, PFLUG H D. Cloudinidae n. fam., calcareous tubes from the Vendian and Lower Cambrian[J].Senckenberg Biol,1985,65:413-431.
[110]CONWAY MORRIS S, MATTES B W, et al.The early skeletal organism Cloudina: new occurrences from Oman and prossibly China[J].Amer Jour Sci, 1990, 290A:245-260.
[111]陈哲,孙卫国.陕南晚震旦世后生动物管状化石Cloudina和Sinotubulites[J].微体古生物学报,2001,18(2):180-193,199-202.
CHEN Z, SUN W G.Late sinian(tubular) metazoan fossils:Cloudina and sinotubulites from southern Shaanxi[J].Acta Micrapalaeont Ologica Sinica,2001,18(2):180-193,199-202.
[112]CAI Y P, CORTIJO I, SCHIFFBAUER J D,et al. Taxonomy of the late Ediacaran index fossil Cloudina and a new similar taxon from South China[J].Precambrian Research, 2017,298:146-156.
[113]PERNET B. Escape hatches for the clonal offspring of Serpulid polychaetes[J]. Biol Bull,2001,200(2):107-117.
[114]VINN O, ZATON M. Inconsistencies in proposed annelid affinities of early biomineralized organism Cloudina (Ediacaran): Structural and ontogenetic evidences[J].Carnets de Goélogie, 2012,C,G2012-A03: 39-47.
[115]MIN X, HUA H, CAI Y,et al. Asexual reproduction of tubular fossils in the terminal Neoproterozoic Dengying Formation, South China[J].Precmbrian Research, 2019, 322:18-23.
[116]ADORNO R R, DO CARMO D A,GERMS G, et al. Cloudina lucianoi (Beurlen & Sommer, 1957), Tamengo Formation, Ediacaran, Brazil: Taxonomy, analysis of stratigraphic distribution and biostratigraphy[J]. Precambrian Research,2017, 301:19-35.
[117]SEILACHER A. Biomat-related lifestyles in the Precambrian[J]. Palaios,1999, 14:86-93.
[118]CAI Y P,HUA H,SCHIFFBAUER J D,et al.Tube growth patterns and microbial mat-related lifestyles in the Ediacaran fossil Cloudina,Gaojiashan Lagersttte,South China[J].Gondwana Res,2014,25:1008-1018.
[119]BRAIN C K. Some observations on Cloudina, a terminal Proterozoic index fossil from Namibia[J].Journal of African Earth Sciences,2001,33(3/4):475-480.
[120]GAUCHER C, SIAL A N, BLANCO G,et al.Chemostratigraphy of the lower arroyo del soldado group (Vendian,Uruguay) and palaeoclimatic implications[J].Gondwana Research,2004,7(3):715-730.
[121]BOGGIANI P C, GAUCHER C.Cloudina from the Itapucumi Group (Vendian, Paraguay): Age and correlations[C]∥In:1st Symposium on Neoproterozoic - Early Paleozoic Events in SW-Gondwana, 2004:13-15.
[122]WARREN L V, FAIRCHILD T R, GAUCHER C,et al. Corumbella and in situ Cloudina in association with thrombolites in the Ediacaran Itapucumi Group, Paraguay[J]. Terra Nova,2011,23(6):382-389.
[123]YOCHELSON E L, STUMP E. Discovery of early Cambrian fossils at Taylor Nunatak, Antarctica[J]. Journal of Paleontology, 1977,51:872-875.
[124]SIGNOR P W, MCMENAMIN M A S, GEVIRTZMAN D A,et al.Two new pre-trilobite faunas from western North America[J].Nature,1983,303:415-418.
[125]SIGNOR P W, MOUNT J F, ONKEN B R.A pre-trilobite small shelly fauna from the White-Inyo region of eastern California and western Nevada[J].Journal of Paleontology,1987,61(3):425-438.
[126]HOFMANN H J, MOUNTJOY E W. Namacalathus-Cloudina assemblage in Neoproterozoic Miette Group (Byng Formation), British Columbia: Canada′s oldest shelly fossils[J]. Geology,2001,29(12):1091-1094.
[127]GROTZINGER J P, WATTERS W A, KNOLL A H.Calcified metazoans in thrombolite-stromatolite reefs of the terminal Proterozoic Nama Group, Namibia[J].Paleobiology,2000, 26(3):334-359.
[128]WOOD R A. Proterozoic modular biomineralized metazoan from the Nama Group, Namibia[J].Science,2002,296(5577):2383-2386.
[129]ZHURAVLEV A Y, LIN E,GMEZ VINTANED J A G,et al.New finds of skeletal fossils in the terminal Neoproterozoic of the Siberian platform and Spain[J].Acta Palaeontologica Polonica,2012,57(1):205-224.
[130]LIN E, PALACIOS T, PEREJN A. Precambrian-Cambrian boundary and correlation from southwestern and central part of Spain[J].Geological Magazine,1984,121:221-228.
[131]LIN E, GOZALO R, PALACIOS T,et al. Cambrian[M]∥The Geology of Spain, London:The Geological Society, 2002:17-29.
[132]PALACIOS MEDRANO T. Microfósiles depared orgnicadel Proterozoico Superior (región central de la Península Ibérica)[J].Memorias del Museo Paleontológico de la Universidad de Zaragoza,1989,3:1-91.
[133]VIDAL G, PALACIOS T, GMEZ VINTANED J A,et al.Neoproterozoic-early Cambrian geology and palaeontology of Iberia[J]. Geological Magazine,1994,131(6):729-765.
[134]VIDAL G,PALACIOS T,MOCZYDOWSKA M,et al.Age constraints from small shelly fossils on the early Cambrian terminal Cadomian Phase in Iberia[J].Geologiska Freningens Stockholm Frhandlingar,1999,121:137-143.
[135]GMEZ VINTANED J A. Therío Husosection. In: LIN E, GMEZ VINTANED J A, GOZALO R.(eds.), II Field Conference of the Cambrian Stage Subdivision Working Groups[C]∥Internattonal Subcommission on Cambrian Stratigraphy. Spain, Field Trip Guide and Abstracts, Zaragoza:Universidad de Zaragoza,1996:28-31.
[136]FERNNDEZ-REMOLAR D C. Latest Neoproterozoic to Middle Cambrian body fossil record in Spain (exclusive of trilobites and archaeocyaths) and their stratigraphic significance[J].Geologiska Freningens i Stockholm Frhandlingar, 2001,123(2):73-80.
[137]FERNNDEZ-REMOLAR D C, GARCA-HIDALGO J F, MORENO-EIRIS E. Interésdel registrode losprimeros organismosen elArcaico y Proterozoico[J]. Boletín de la Real Sociedad Espaola de Historia Natural (Sección Geológica), 2005, 100: 177-209.
[138]ZHURAVLEV A Y,LIN E, GMEZ VINTANED J, et al.New finds of the oldest skeletal fossils in the terminal Neoproterozoic of the Siberian Platform and Spain[C]∥In:21 ème Réunion des Sciences de la Terre, Dijon,Joint Earth Sciences Meeting, 43. Société géologique de France, Paris,2006.
[139]JENSEN S, PALACIOS T, MART MUS M. A brief review of the fossil record of the Ediacaran-Cambrian transition in the area of Montes de Toledo-Guadalupe, Spain[J].Geological Society, Special Publications,2007,286(1):223-235.
[140]CORTIJO I, MART MUS M, JENSEN S, et al. A new species of Cloudina from the terminal Ediacaran of Spain[J]. Precambrian Research, 2010,176(1/2/3/4):1-10.
[141]SUN W G. Early multicellular fossils[M].Procedings of the 84th: Bengston S, eds. Early life on Earth. Nobel Symposium 84. New York: Columbia University Press, 1994, 369.
[142]GU P, ZHANG G, TANG F, et al. The Division of the Late Ediacaran-Cambrian Boundary Interval Stratigraphy and New Options of Index Fossil FAD in South China[J]. Acta Geologica Sinica, 2018(3):449-465.
[143]CHEN Z, BENGTSON S, ZHOU C M,et al.Tube structure and original composition of Sinotubulites: shelly fossils from the late Neoproterozoic in southern Shaanxi, China[J].Lethaia,2008,41(1):37-45.
[144]陳孟莪, 王义昭, 峡东区上震旦统灯影组中段的管状化石[J].科学通报,1977,22( Z1):219-221.
[145]MCMENAMIN M A S. Basal Cambrian small shelly fossils from the La Cienega Fromation,Northestern Sonora,Mexico[J].J Paleont,1985,59(6):1414-1425.
[146]岳昭.扬子地台前寒武系一寒武系过渡层骨骼微化石生物地层学及古生物学[D].北京:中国地质科学院,1990.
[147]华洪,张录易,张子福,等.晚震旦世高家山生物群化石新材料[J].古生物学报,2000,39(3):381-390.
HUA H,ZHANG Y L,ZHANG Z F,et al.New fossil evidences from latest neoproterozoic Gaojiashan biota,South Shaanxi[J].Acta Palaeontologica Sinica,2000,39(3):381-390.
[148]孙勃, 华洪, 蔡耀平. 陕西宁强埃迪卡拉系顶部管状化石Sinotubulites形态学与古生态学新知[J]. 古生物学报,2012,51(1):107-113.
SUN B,HUA H,CAI Y P.Morphology and palaeoecology of the late Ediacaran tubular fossil Sinotybulites[J].Acta Palaeontologica Sinica,2012,51(1):107-113.
[149]CAI Y, XIAO S, HUA H,et al.New material of the biomineralizing tubular fossil Sinotubulites from the late Ediacaran Dengying Formation, South China[J]. Precambrian Research, 2015, 261:12-24.
[150]VORONOVA L G,MISSARZHEVSKY V V.Finds of algae and worm tubes in the Cambrian and Precambrian boundary strata on the north of the Siberian Platform [in Russian][J].Doklady Akademii Nauk SSSR,1969,184:204-210.
[151]KOUCHINSKY A,BENGTSON S,FENG W M,et al.The lower Cambrian fossil anabaritids: affinities, occurrences and systematics[J].Journal of Systematic Palaeontology,2009,7(3):241-298.
[152]GLAESSNER M F. The dawn of animal life-A biohistorical study[M].Cambrige: Cambrige University Press,1984:1-241.
[153]GEHLING J G.The case for Ediacaran fossil roots to the Metazoan tree[J].Geological Society of India Memoir,1991,20:181-224.
[154]SEILACHER A.Vendobionta and Psammacorallia: Lost constructions of Precambrian evolution[J].Geol Soc London,1992, 149(4):607-613.
[155]ZHURAVLEV A Y. Were Ediacaran Vendobionta multicellulars?[J].Neues Jahrb Geol Palaontol Abh,1993,190(2):299-314.
[156]RETALLACK G J. Were the Ediacaran fossils lichens?[J].Paleobiology,1994,20(4):523-544.
[157]STEINER M, REITNER J. Evidence of organic structures in Ediacara-type fossils and associated microbial mats[J]. Geology, 2001, 29(12):1119-1122.
[158]PTERSON K J, WAGGONER B, HAGADORN J W. A fungal analog for newfoundland ediacaran fossils?[J]. Integr Comp Biol, 2003, 43(1):127-136.
[159]LAFLAMME M, DARROCH S A F, TWEEDT S M,et al. The end of the Ediacara biota: extinction, biotic replacement, or Cheshire Cat?[J].Gondwana Res,2013,23(2):558-573.
[160]BUDD GE, JENSEN S.The origin of the animals and a ′Savannah′ hypothesis for early bilaterian evolution[J].Biol Rev,2015, 92(1):446-473.
[161]ERWIN D H. Early metazoan life: Divergence, environment and ecology[J].Philos Trans R Soc B,2015,370(1684):20150036.
[162]WAGGONER B. The Ediacaran biotas in space and time[J].Integr Comp Biol,2003,43(1):104-13.
[163]GRAZHDANKIN D V.Patterns of distribution in the Ediacaran biotas: Facies versus biogeography and evolution[J].Paleobiology,2004,30(2):203-221.
[164]GEHLING J G, DROSER M L. How well do fossil assemblages of the Ediacara biota tell time? [J]. Geology, 2013, 41(4):447-450.
[165]NARBONNE G M. The Ediacara biota: Neoproterozoic origin of animals and their ecosystems[J]. Annual Reviews of Earth and Planetary Sciences, 2005,33(1):421-442.
[166]NARBONNE G M, LAFLAMME M, TRUSLER P W,et al. Deep-water Ediacaran fossils from Northwestern Canada: Taphonomy, ecology, and evolution[J].Journal of Paleontology, 2014,88(2):207-223.
[167]DARROCH S A F, LAFLAMME M, CLAPHAMM E. Population structure of the oldest known macroscopic communities from Mistaken Point, Newfoundland[J].Paleobiology, 2013, 39(4):591-608.
[168]LIU A G, KENCHINGTON C G, MITCHELL E G. Remarkable insights into the paleoecology of the Avalonian Ediacaran macrobiota[J].Gondwana Research, 2015,27(4):1355-1380.
[169]KENCHINGTON C G,WILBY P R.Of time and taphonomy:Preservation in the Ediacaran[C]∥LAFLAMME M, SCHIFFBAUER J D,DARROCH S A F.Reading and Writing of the Fossil Record: Preservational Pathways to Exceptional Fossilization.Paleontological Society Papers. Paleontological Society Short Course, 2014, 20:101-122.
[170]MARTIN M W, GRAZHDANKIN D V, BOWRING S Aet al. Age of Neoproterozoic bilaterian body and trace fossils, White Sea, Russia: Implications for Metazoan evolution[J].Science, 2000,288(5467):841-845.
[171]ZAKREVSKAYA M. Paleoecological reconstruction of the Ediacaran benthic macroscopic communities of the White Sea (Russia)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 410:27-38.
[172]IVANTSOV A Y, FEDONKIN M A.Conulariid-like fossil from the Vendian of Russia: A metazoan clade across the Proterozoic/Palaeozoic boundary[J].Palaeontology,2002,45(6):1219-1229.
[173]CARBONE C A, NARBONNE G M, MACDONALD F A et al. New Ediacaran fossils from the uppermost Blueflower Formation, northwest Canada:Disentangling biostratigraphy and paleoecology[J].Journal of Paleontology, 2015, 89(2):281-291.
[174]CLAPHAM M E, NARBONNE G M, GEHLING J G et al. Thectardis avalonensis:A new Ediacaran fossil from the Mistaken Point biota, Newfoundland[J]. Journal of Paleontology, 2004, 78(6):1031-1036.
[175]SAPPENFIELD A, DROSER M L, GEHLING J G. Problematica, trace fossils, and tubes within the Ediacara Member (South Australia): Redefining the Ediacaran trace fossil record one tube at a time[J]. Journal of Paleontology, 2011,85(2):256-265.
[176]JOEL L V, DROSER M L, GEHLING J G. A New Enigmatic, Tubular Organism from the Ediacara Member, Rawnsley Quartzite[J]. South Australia.Journal of Paleontology, 2014, 88(2):253-262.
[177]GLAESSNER M F.Trace fossils from the Precambrian and basal Cambrian[J].Lethaia,1969, 2(4):369-393.
[178]YANISHEVSKY M. Ob ostatkah trubchatyh cherveiiz kembriyskoy sineygliny[J].Ezhegodnik Russkogo Paleontologicheskogo Obchestva, 1926, 4: 99-112.
[179]FEDONKIN M A, GEHLING K, GREY GM, et al. The Rise of Animals Evolution and Diversification of the Kingdom Animalia[M]. Washington: The Johns Hopkins University Press, 2007:1-326.
[180]MOCZYDLOWSKA M, WESTALL F, FOUCHER F. Microstructure and biogeochemistry of the organically Preserved Ediacaran Metazoan Sabellidites[J].Journal of Paleontology, 2014,88(2):224-239.
[181]HAHN G HAHN R, LEONARDOS O H,et al.Krperlich erhaltene Scyphozoen-Reste aus dem Jungprkambrium Brasiliens[J]. Geologica et Paleontologica,1982,16:1-18.
[182]SMITH E F, NELSON L L, TWEEDT S M,et al.A cosmopolitan late Ediacaran biotic assemblage:New fossils from Nevada and Namibia support a global biostratigraphic link[J].Proceedings Royal Society B,2017,284(1858):20170934.
[183]WALDE D H G, LEONARDOS O H, HAHN G,et al.The first Precambrian megafossil from South America, Corumbella werneri[J]. Anais da Academia Brasileira de Ciencias,1982,54:461.
[184]BABCOCKA L E, GRUNOW A M, SADOWSKIC G R,et al. Corumbella, an Ediacaran-grade organism from the late neoproterozoic of brazil[J].Palaeogeography, Palaeoclimatology, Palaeoecology,2005,220(112):7- 18.
[185]PACHECO M L A F. Reconstituio Morfológica e Anlise Sistemtica de Corumbella werneri Hahn et al. 1982 (Formao Tamengo, Ediacariano, Grupo Corumb), Mato Grosso do Sul, Brasil: Implicaes paleoecológicas e tafonómicas[D].Tese de Doutorado, Instituto de Geociências, Universidade de So Paulo, 2012:140.(Unpublished).
[186]PACHECO M L A F, LEME J, MACHADO A. Taphonomic analysis and geometric modelling for the reconstruction of the Ediacaran metazoan Corumbella werneri Hahn et al. 1982 (Tamengo Formation, Corumba Basin, Brazil)[J]. Journal of Taphonomy, 2011, 9:269-283.
[187]PACHECO M L A F, GALANTE D, RODRIGUES F. Insights into the skeletonization, lifestyle, and affinity of the unusual Ediacaran fossil Corumbella[J].PLoS ONE, 2015,10: e0114219.
[188]ITEN H V,MARQUES A C, LEME J,et al.Origin and early diversification of the phylum Cnidaria Verrill: Major developments in the analysis of the taxon′s Proterozoic-Cambrian history[J].Palaeontology,2014,57(4):677-690.
[189]ERDTMANN B D, WALDE D, CARMO DO D A. New data and observations on Ediacaran (Latest Precambrian) fossils from the Paraguai Belt in SW Brazil[C]∥20th Colloquium on Latin American Earth Sciences, Kiel, Germany,2007:206-207.
[190]ERDTMANN B D.The Infra-Cambrian bioradiation event: A review from the perspective of the Yangtze Plate in South China and correlation with coeval strata in Brazil[J]. Revista Brasileira de Paleontologia, 2004,7(2):223-230.
[191]WALDE D, WEBER B, ERDTMANN B D,et al. Taphonomy of Corumbella werneri from the Ediacaran of Brazil: Sinotubulitid tube or conulariid test?[J]. Alcheringa: An Australasian Journal of Palaeontology, 2019,43(3):335-350.
[192]ROUSE G W. Cladistic Analysis of Siboglinidae Caullery, 1914 (Polychaeta, Annelida): Formerly the phyla Pogonophora and Vestimentifera[J]. Zoological Journal of the Linnean Society, 2001,132:55-80.
[193]TANG F, YIN C, BENGTSON S,et al.Octoradiate Spiral Organisms in the Ediacaran of South China[J].Acta Geol Sin, 2008,82:27-34.
[194]ZHU M Y, GEHLING J G, XIAO S H, et al. Eight-armed Ediacara fossil preserved in contrasting taphonomic windows from China and Australia[J].Geology, 2008,36(11):867-870.
[195]TANG F, BENGTSON S, WANG Y,et al. Eoandromeda and the origin of Ctenophora[J].Evol Dev,2011,13(5):408-414.
(編 辑 亢小玉)
作者简介:
华洪,博士,2006年入选教育部“新世纪优秀人才支持计划”,国家基金委创新研究群体、科技部重点领域创新团队骨干成员。西北大学地质学系教授,博士生导师。主要研究领域:前寒武纪古生物学和地层学,尤其是“寒武大爆发”前的生物群面貌及其环境变化,骨骼生物的起源及早期演化、化石埋藏学及中元古代新元古代叠层石和生物地层等。在Nature, Science, Nature Communication, Geology, Chemical Geology, Gondwana Research,Precambrian Research, Palaios,Geological Joural,科学通报,中国科学,古生物学报,地层学杂志等权威及核心刊物上发表论文100余篇。主持完成国家自然科学基金重点项目、面上项目、基地基金项目等13项,作为骨干成员参与国家基金委重大项目、科技部“973”项目、中国科学院先导性项目等多项。参与完成的埃迪卡拉纪“蓝田生物群”研究获2011年度中国十大地质科技进展, 2013年度获江苏省科学技术奖一等奖。目前担任“微体古生物学报”副主编、“生物进化”编委,兼任中国古生物学会副理事长、中国地质教育学会副会长、中国地层委员会委员、中国藻类学会理事、陕西省古生物学会理事长。
收稿日期:2020-03-09
基金项目:国家自然科学基金资助项目(41672025, 41572012,41621003,41890844);中科院先导专项项目(XDB26000000) ; 国家重点研发项目(2017YFC0603101)
作者简介:华洪,男,江苏无锡人,教授,博士生导师,从事古生物学与地层学研究。
