灰葡萄孢BcPDR1基因差异表达基因的筛选和鉴定
2020-04-23刘晓颖吴艳丽袁雪梅曹宏哲邢继红董金皋
刘晓颖,吴艳丽,李 白,袁雪梅,曹宏哲,张 康,邢继红,董金皋
(1.河北农业大学 河北省植物生理与分子病理学重点实验室/真菌毒素与植物分子病理学实验室,河北保定 071000;2.河北省正定县农业农村局,河北 正定 050800)
灰葡萄孢(Botrytis cinerea)是一种重要的植物致病菌,在世界温带地区分布较为广泛。灰葡萄孢能够引起200多种植物的灰霉病,包括大多数蔬菜和水果作物、树木和花卉[1-3]。它可以侵染许多植物的组织器官,如叶片、茎、花和果实等,作为一种死体营养型植物病原菌,经常在农产品收获后造成严重的损失。目前,灰葡萄孢已经成为死体营养型植物病原真菌的模式真菌[4-5]。因此,深入发掘灰葡萄孢的生长发育和致病相关基因,并对其功能和机制进行深入研究,可为灰霉病的防治以及其他死体营养型植物病原真菌的研究提供理论依据。
灰葡萄孢的主要致病机制是产生有助于杀死寄主细胞的酶类,帮助病菌的菌丝或分生孢子入侵,并从寄主细胞中获取营养[6]。果胶是植物细胞壁的主要成分,可被一系列真菌果胶酶降解。其中,根据病菌侵染的阶段和侵染寄主的不同,内多聚半乳糖醛酸酶(Endopolygalacturonase, PG)基因(BcPG1-6)表现出不同的表达模式,表明这些基因具有果胶分解活性的特殊功能[7]。其中,BcPG1基因的破坏导致病原菌对番茄叶片和果实以及苹果的毒力降低[8];BcPG2基因的敲除可显著降低病原菌对番茄和蚕豆的毒力,突变体的病斑形成延迟、病斑扩展速率降低50%~80%[9]。研究表明,BcPG2通过破坏位于背斜细胞壁的果胶网络参与病菌的穿透[8],而BcPG1则需要在定殖过程中通过中膜破坏果胶网络[8]。果胶甲基酯酶(Pectin methylesterase, PME)BcPME1被认为通过将果胶去甲基化为果胶酸来促进BcPGs的作用,且灰葡萄孢Bd90菌株中BcPME1基因的失活导致灰葡萄孢对几个植物寄主的毒力大大降低[10-11]。非果胶降解的细胞壁降解酶(CWDEs),如纤维素酶或半纤维素酶,也促进了灰葡萄孢的侵染。如,灰葡萄孢木聚糖酶(Endo-b-1,4-xylanas, Xyn11A)基因失活导致其对番茄叶片和葡萄浆果的致病性降低[12-13]。
实验室前期研究获得了一株致病力完全丧失的突变体BCt89,明确了其突变基因为BcPDR1,推测BcPDR1基因参与灰葡萄孢的致病过程。利用基因敲除与功能互补回复技术,获得了BcPDR1基因的敲除突变体ΔBcPDR1和互补回复突变体CE,通过表型和致病力分析,明确了BcPDR1参与病菌的生长、发育和致病过程[14],但其调控病菌致病力的分子机制尚不清楚。本研究利用数字基因表达谱技术(Digital Gene Expression Profiling, DGE),筛选BcPDR1基因影响的差异表达基因,对差异表达基因进行GO富集分析,探讨BcPDR1基因影响病菌致病力的机制;同时,筛选差异表达的细胞壁降解酶基因,并利用Real-time PCR技术检测细胞壁降解酶基因在灰葡萄孢野生型BC22和BcPDR1基因突变体中的表达情况,为阐明BcPDR1基因调控灰葡萄孢致病力的分子机制奠定基础。
1 材料和方法
1.1 试验材料
灰葡萄孢野生型菌株BC22、BcPDR1基因的T-DNA插入突变体BCt89、敲除突变体ΔBcPDR1、回复突变体CE,均由河北农业大学真菌毒素与植物分子病理学实验室保存并提供。
1.2 数字基因表达谱(DGE)实验流程
分别提取灰葡萄孢野生型菌株BC22、BcPDR1基因的T-DNA插入突变体BCt89、敲除突变体ΔBcPDR1、回复菌株CE的总RNA,反转录获得各菌株的cDNA。将各菌株的cDNA进行末端修复、加碱基A、加接头处理和目的片段的扩增、回收后,即完成了文库的构建。文库构建好后需进一步用Illumina平台进行测序,测序策略为SE50。
1.3 数字基因表达谱(DGE)的数据分析
测序完成后的原始数据以FASTQ格式存储,对这些数据进行筛选,得到高质量序列。这些高质量序列还要进行质控检查,合格的数据进行表达量分析和比对信息统计。为保证得到准确而完整的数据,对不合格的样品需要重新测序。
1.4 差异表达基因的筛选
利用DGESeq软件,对灰葡萄孢野生型菌株BC22、BcPDR1基因的T-DNA插入突变体BCt89、敲除突变体ΔBcPDR1和回复菌株CE的12个样本中获得的数据进行差异表达分析,利用Benjamini和Hochberg方法使假阳性在一定范围内,即P_value≤0.05时认为筛选得到的基因为差异表达基因。对比分析各菌株的表达谱数据,得到上下调基因个数。
1.5 差异表达基因的GO富集分析
通过GO富集分析得到每个GO term的基因数目,再利用超几何检验可以得到和整个基因组对比在差异表达基因中显著富集的GO term。这些GO term只有满足校正后的P_value≤0.05才被认为是需要的数据。
1.6 细胞壁降解酶基因的热图分析
根据数字基因表达谱数据,获得细胞壁降解酶基因在灰葡萄孢野生型菌株BC22、BcPDR1基因的T-DNA插入突变体BCt89、敲除突变体ΔBcPDR1、回复菌株CE中的表达谱数据,通过HemI软件绘制热图。
1.7 Real-time PCR分析
利用Real-time PCR,对细胞壁降解酶基因在灰葡萄孢野生型菌株BC22、BcPDR1基因的T-DNA插入突变体BCt89、敲除突变体ΔBcPDR1、回复菌株CE中的表达水平进行分析。以Tubulin基因为内参,用基因特异性引物(表1)与SYBR Green I荧光染料进行荧光定量PCR检测。Real-time PCR反应体系:模板cDNA 2.0 μL、Mix 7 μL、引物各0.5 μL。反应程序:95 ℃ 5 min,94 ℃ 10 s,55 ℃ 15 s,72 ℃ 30 s,循环数为30次,每个样品重复3次。

表1 Real-time PCR引物设计Table 1 Real-time PCR primers design
2 结果与分析
2.1 差异表达基因的筛选
采用DESeq进行基因差异表达分析,并选取|log2Ratio|≥1和q<0.05的基因作为差异表达基因。结果发现,与灰葡萄孢野生型BC22对比,BCt89突变体中1 493个基因上调表达,850个基因下调表达;ΔBcPDR1突变体中1 561个基因上调表达,848个基因下调表达;CE中940个基因上调表达,723个基因下调表达(图1-A)。将BCt89、ΔBcPDR1和CE突变体分别与野生型BC22比较获得的差异表达基因进行overlap分析,筛选出1356个差异表达基因;将BCt89、ΔBcPDR1突变体分别与野生型BC22和CE突变体比较获得的差异表达基因进行overlap分析,筛选出1116个差异表达基因;将获得的1 116个基因再与野生型BC22、回复菌株CE共同包含的差异基因进行过滤,筛选出933个差异表达基因(图1-B)。
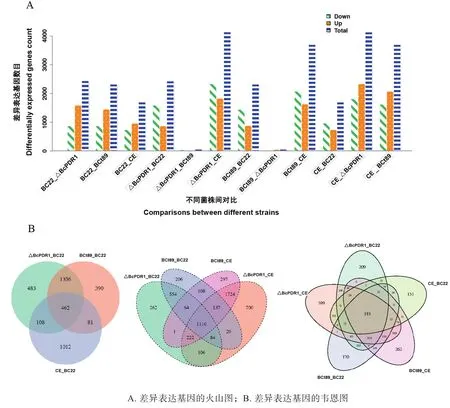
图1 差异表达基因的筛选Fig.1 Screening of differentially expressed genes
2.2 差异表达基因的GO富集分析
对获得的933个差异表达基因进行GO功能富集分析,结果发现,这些差异基因主要集中在生物过程(biological process)中的细胞过程(cellular process)、代谢过程(metabolic process)和单一生物过程(single-organism process),细胞组分(cellular component)中的细胞部分(cell part)、细胞器(organelle)和膜部分(membrane part),分子功能(molecular function)中的催化(catalysis)和结合(binding)功能(图2)。

图2 差异表达基因的GO功能富集分析Fig.2 GO enrichment analysis of differentially expressed genes
2.3 细胞壁降解酶基因的热图分析
利用数字基因表达谱数据,筛选获得了14个差异表达的胞壁降解酶基因(见图3)。

图3 细胞壁降解酶基因的热图分析Fig.3 Heat map analysis of cell wall degrading enzyme genes
热图分析发现,14个胞壁降解酶基因在突变体BCt89和ΔBcPDR1中的表达水平与在野生型BC22和回复突变体CE中的表达水平呈现比较明显的差别,其中BcPG5、BcPGX1、BcAMS1、BcMNL1基因在突变体BCt89和ΔBcPDR1中的表达水平明显低于野生型BC22和回复突变体CE,而BcPME2、BcPG1、BcPG3、BcPME1、BcPG6基因在突变体BCt89和ΔBcPDR1中的表达水平明显高于野生型BC22和回复突变体CE。
2.4 Real-time PCR分析
利用Real-time PCR技术,对14个胞壁降解酶基因在野生型BC22和BcPDR1基因突变体中的表达水平进行分析。结果发现,突变体BCt89和ΔBcPDR1中各胞壁降解酶基因的表达水平与在野生型BC22和回复突变体CE中的表达水平呈现明显的差别,并且与数字基因表达谱的结果基本一致。其中,BcMNS1、BcROT2、BcPME2、BcPG1、BcPG3、BcPME1、BcPG6、BcPG4基因在突变体BCt89和ΔBcPDR1中的表达水平明显高于野生型和回复菌株 CE,而BcPG5、BcCUTA、BcPGX1、BcAMS1、BcMNL1、BcCUTB在突变体BCt89和ΔBcPDR1中的表达水平明显低于野生型和回复菌株CE(图4)。

图4 胞壁降解酶基因的表达分析Fig.4 Analysis of expression of cell wall degrading enzyme genes
3 结论与讨论
灰葡萄孢(B.cinerea)为子囊菌门、核盘菌科、孢盘菌属的一种重要的植物病原真菌,能够引起数百种双子叶植物的灰霉病[15]。作为死体营养型植物病原真菌的模型,其生长发育和致病相关基因的挖掘及机制研究,不仅可为灰霉病的防治提供理论依据,还可为其他死体营养型病原菌相关基因的功能研究提供参考。本实验室前期研究获得了灰葡萄孢致病相关基因BcPDR1,明确了BcPDR1参与病菌的生长、发育和致病力过程[14],但其调控病菌致病力的分子机制尚不清楚。本研究利用数字基因表达谱技术(DGE),筛选获得了933个受BcPDR1基因影响的差异表达基因,GO富集分析发现差异表达基因主要集中在细胞过程、新陈代谢过程、细胞组分和催化作用等方面。研究结果将为进一步阐明BcPDR1调控灰葡萄孢生长发育和致病力的分子机制奠定基础。
作为一种死体营养型植物病原真菌,灰葡萄孢可以产生多种细胞壁降解酶(CWDEs),通过这些酶破坏植物细胞壁,并将其作为营养来源。本研究利用数字基因表达谱,获得了14个差异表达的胞壁降解酶基因,热图分析和Real-time PCR分析发现,14个胞壁降解酶基因在突变体BCt89和ΔBcPDR1中的表达水平被明显上调或下调,表明这些胞壁降解酶基因在BcPDR1基因引起的致病过程中可能发挥重要作用。果胶甲基酯酶催化半乳糖醛酸酯的去甲基化,从而使果胶可被多半乳糖醛酸酶和果胶酸裂解酶降解。研究发现,灰葡萄孢Bd90菌株中BcPME1基因的敲除突变体对苹果、葡萄和拟南芥叶片的毒性较弱[10],而菌株B05.10中BcPME1、BcPME2的敲除突变体的毒力均没有表现出任何减弱[11]。表明BcPME1、BcPME2基因在不同菌株中可能具有不同的功能。本研究中,发现BcPME1、BcPME2基因在突变体BCt89和ΔBcPDR1中均具有较高水平表达,表明BcPDR1基因负调控BcPME1、BcPME2基因的表达,推测BcPME1、BcPME2基因在BcPDR1调控病菌致病过程中具有重要的作用。
有文献报道,灰葡萄孢基因组包含至少6个BcPG基 因[16],ten Have等 研 究 了 6个BcPG(BcPG1~BcPG6)在番茄、蚕豆、苹果和西葫芦4种寄主植物上的表达,结果发现,这些BcPG基因的表达模式明显不同。其中,BcPG1在所有组织中均有表达,BcPG2在除苹果外的所有植物感染早期均有表达,而BcPG3和BcPG5仅在苹果组织中有表达[7]。本研究利用数字基因表达谱技术,获得了差异表达的BcPG1、BcPG3、BcPG4、BcPG5和BcPG6基因,发现BcPG1、BcPG3、BcPG6基因在突变体BCt89和ΔBcPDR1中高水平表达,BcPG4、BcPG5基因在突变体BCt89和ΔBcPDR1中低水平表达。表明BcPDR1负调控BcPG1、BcPG3、BcPG6基因的表达,正调控BcPG4、BcPG5基 因 的 表 达, 推 测BcPG1、BcPG3、BcPG4、BcPG5和BcPG6基因在BcPDR1调控灰葡萄孢致病过程中可能发挥不同的作用。
