组蛋白去乙酰化酶3对恒定型自然杀伤性T细胞发育和功能的调控作用
2020-04-20郑全辉袁鸿儒马剑楠刘兆基刘倚博张爱红郑爱华李会婷
郑全辉,雷 冰,袁鸿儒,马剑楠,刘兆基,张 郡,刘倚博,张 冰,张爱红,郑爱华,张 颖,李会婷,田 枫
(1.河北省慢性疾病重点实验室 河北省唐山市慢性病基础研究重点实验室 华北理工大学基础医学院免疫学系,河北 唐山 063210; 2.河北医科大学第三医院骨外科,河北 石家庄 050051; 3.河北省唐山市工人医院ICU,河北 唐山 063000; 4.北京大学医学部实验动物科学部,北京 100191)
恒定型自然杀伤性T细胞(invarient nature killer T cells, iNKT)产生于胸腺, 是一群既表达有限T细胞受体α链和β链,同时也表达NK细胞受体(NK1.1)的固有淋巴细胞亚群[1]。根据其细胞表面分子CD44和NK1.1的表达差异,iNKT在胸腺的发育和成熟可分为3个阶段:最初阶段非成熟iNKT细胞表现为CD44-NK1.1-;随iNKT细胞成熟程度的增加,CD44表达首先增加,表现为CD44+NK1.1-;成熟阶段iNKT细胞NK1.1表达明显增加,表现为CD44+NK1.1+[2]。由于iNKT细胞在活化后能够产生大量Th1和Th2型细胞因子,如干扰素γ(interferon-γ,IFN-γ)和白细胞介素4(interleukin-4, IL-4)等,因此在感染和肿瘤等多种疾病中发挥作用[3-5]。已有研究[6-9]发现:组蛋白去乙酰化酶3(histone deacetylase 3, HDAC3)在传统T细胞、B细胞、巨噬细胞和树突状细胞的发育和功能调控中发挥促进或抑制作用,而HDAC3对iNKT细胞发育和功能的影响目前尚不完全清楚。本研究采用T细胞特异的hdac3基因敲除小鼠,探讨HDAC3对iNKT细胞发育和功能的影响。
1 材料与方法
1.1 实验动物、主要试剂和仪器C57BL/6背景的hdac3floxP转基因小鼠(Stock No:024119)和CD4启动子/增强子/沉默子-Cre转基因小鼠(Stock No:022071)购自美国Jackson实验室,并在华北理工大学SPF级小鼠房饲养、繁殖,动物合格证号:SCSK(京)2014-0004。hdac3floxP小鼠和CD4启动子/增强子/沉默子-Cre小鼠交配、繁殖子代后经细胞基因组PCR检测鉴定可获得T细胞特异的hdac3基因敲除小鼠(hdac3floxP+CD4Cre+,HDAC3KO),将hdac3基因正常小鼠作为野生型正常对照(hdac3floxP+CD4Cre-,WT)。实验选取4~8周龄及性别匹配的HDAC3KO和WT小鼠进行研究。实验操作按华北理工大学实验动物管理委员会规定进行。细胞DNA提取试剂盒购自天根生物科技(北京)有限公司,荧光素标记的抗小鼠TCRβ(H57-597)、NK1.1(PK136)、CD44(IM7), CD122(5H4)、CD69 (H1.2F3)、DX5(HMα2)、CD4(RM4-5)、CD8(53-6.7)、IL-4(11B11)、IFN-γ(XMG1.2)、生物素标记抗小鼠CD3(17A2)抗体及淋巴细胞固定/打孔试剂盒购自美国BD或eBioscience公司,抗生物素磁珠购自德国美天旎生物技术有限公司,半乳糖神经酰胺(α-Galcer)及荧光素标记的CD1d-α-Galcer四聚体购自日本麒麟公司,大鼠抗小鼠FcR单克隆抗体(2.4G2)取自2.4G2杂交瘤细胞培养上清,RPMI1640培养液、胎牛血清、HEPES、青-链霉素、谷氨酰胺、莫能菌素、红细胞裂解液及小鼠IFN-γ和IL-4 ELISA检测试剂盒购自北京达科为生物技术公司,引物由生工生物工程(上海)股份有限公司合成。低温冷冻离心机购自德国Beckan公司,FACSAria Ⅱ流式细胞仪购自美国BD公司。
1.2 小鼠基因型鉴定采用FACSAriaⅡ流式细胞仪分别分选HDAC3KO和WT小鼠胸腺CD4+CD8+双阳性T细胞(DP)、CD4单阳性T细胞(CD4SP)、CD8单阳性T细胞(CD8SP)以及脾脏CD4+T细胞(CD4)和CD8+T细胞(CD8)。各取1×106个细胞,按试剂盒操作说明书提取DNA并进行PCR扩增。所用上游引物:5′-CCCAGGTTAGCTTTGAACTCT-3′,下游引物:5′-CCACTGGCTTCTCCTAAGTTC-3′。HDAC3KO小鼠PCR扩增生成211 bp DNA片段,WT小鼠PCR扩增生成935 bp DNA片段。
1.3 流式细胞术分析HDAC3KO小鼠iNKT 细胞数量和发育表型分别分离HDAC3KO和WT小鼠(每组3~5只)胸腺、脾、肝脏和淋巴结并制成单细胞悬液,采用含2%胎牛血清的PBS染色缓冲液洗涤2次,加入2.4G2封闭,4℃孵育30 min。然后直接加入10 μL (1∶10~1∶100稀释)荧光素标记的单克隆抗体,4℃避光孵育30 min,用PBS染色缓冲液洗涤2次,采用FACSAriaⅡ流式细胞仪收集标本,采用FlowJo software或CELLQuest Pro(BD Biosciences) 软件进行数据分析。实验至少重复3次。
1.4 骨髓混合嵌合体小鼠制备采用6~8周龄CD45.1+的B6.SJL小鼠作为受体,以剂量为900 Gy的γ射线照射杀死受体鼠骨髓细胞。在无菌条件下分别提取周龄和性别相同B6.SJL和CD45.2+的HDAC3KO小鼠(每组每种小鼠各取4~6只)骨髓细胞,裂解红细胞后采用生物素标记抗CD3抗体和抗生物素磁珠分选、剔除骨髓中存在的成熟CD3+T细胞。采用无菌PBS缓冲液洗涤并重悬剩余B6.SJL和HDAC3KO骨髓细胞,调整细胞浓度为1×107mL-1。 按1∶1比例混合后经尾静脉注射γ射线照射的B6.SJL小鼠(每只100 μL)。细胞注射8周后采用流式细胞术检测不同骨髓来源iNKT细胞在受体B6.SJL小鼠胸腺的产生和发育情况。实验重复3次。
1.5 iNKT细胞活化和细胞内染色分析将α-Galcer用二甲基亚砜溶解,浓度为100 mg·L-1,取2 μg α-Galcer溶于100 μL无菌PBS中,经尾静脉分别注入HDAC3KO和WT小鼠(每组每种小鼠各取3~5只)体内。4 h后分离小鼠脾细胞,红细胞裂解液处理去除红细胞后每种样本各取2×106个细胞重悬于RPMI 1640培养液(含10%胎牛血清、2 mmol·L-1谷氨酰胺和10 mmol·L-1HEPES),同时加入GolgiStop(1 mg·L-1),于37℃、5%CO2条件下培养2 h。收集培养细胞首先经细胞表面染色,然后经冷PBS洗涤后用新鲜配置的固定/打孔液重悬细胞,加入2.4G2,4℃封闭10 min,直接加入抗小鼠IFN-γ和IL-4抗体进行细胞内染色,4℃避光孵育30 min,用固定/打孔液洗涤细胞2次,流式细胞仪分析iNKT细胞内IFN-γ和IL-4水平。实验至少重复3次。
1.6 酶联免疫吸附实验(ELISA) 检测HDAC3KO小鼠血清细胞因子水平采用结合缓冲液(0.1 mol·L-1Na2HPO4,pH 9.0)稀释抗小鼠IFN-γ和IL-4抗体至2 mg·L-1,按每孔100 μL加入至相应ELISA板,4℃包被过夜。移除包被液,加入封闭缓冲液(1% BSA),每孔200 μL,室温孵育2 h,采用PBS/Tween20洗涤3次后加入1∶50稀释α-Galcer处理的HDAC3KO和WT小鼠(每种小鼠各取3~5只)血清以及倍比稀释的标准血清,每孔100 μL,室温孵育4 h。采用PBS/Tween20洗涤3次后加入底物显色,在450 nm处测量吸光度(A)值,并根据标准曲线计算血清中IFN-γ和IL-4水平(μg·L-1)。实验至少重复3次。

2 结 果
2.1 HDAC3敲除小鼠基因型鉴定对流式细胞术分选的HDAC3KO和WT小鼠胸腺和脾脏T细胞亚群进行基因型鉴定,来自WT小鼠的胸腺DP和CD4SP细胞产生935 bp的 DNA 片段, 而来自HDAC3KO小鼠的胸腺DP、CD4SP、CD8SP细胞和脾脏CD4、CD8 T细胞均产生211 bp 的DNA 片段,说明HDAC3KO小鼠基因型鉴定正确。见图1。

图1 PCR技术鉴定HDAC3KO和WT小鼠胸腺和脾脏T细胞亚群基因型
Fig.1 Identification of genetypes of T cell subtypes of HDAC3KO and WT mice by PCR
2.2 HDAC3敲除小鼠胸腺、脾脏、肝脏和淋巴结中iNKT细胞数量采用抗TCR-β抗体和荷载糖脂抗原α-Galcer的CD1d四聚体(CD1d-Tetramer)染色,流式细胞术分析各组织器官中iNKT细胞比例和数量。结果显示:与WT小鼠比较,HDAC3KO小鼠胸腺、脾脏、肝脏和淋巴结中iNKT细胞比例和数量均明显降低(P<0.05)。见图2、表1和表2。

图2 流式细胞术检测HDAC3KO和WT小鼠胸腺、脾脏、肝脏和淋巴结中iNKT细胞水平
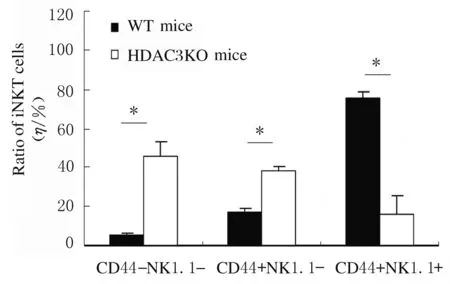
2.3 HDAC3敲除小鼠胸腺中iNKT细胞发育的变化采用抗CD44、NK1.1、 CD122、CD69和DX5抗体染色,流式细胞术检测iNKT细胞发育表型的结果显示:与WT小鼠比较,HDAC3KO小鼠处于非成熟阶段CD44-NK1.1-和CD44+NK1.1-iNKT细胞比例明显增加(P<0.05),而成熟阶段CD44+NK1.1+细胞比例明显降低(P<0.05)。见图3和图4。与WT小鼠比较,HDAC3KO小鼠iNKT细胞中CD122、CD69和DX5表达水平明显降低(P<0.05)。见图5和图6。
表1 HDAC3KO和WT小鼠胸腺、脾脏、肝脏和淋巴结中iNKT细胞比例
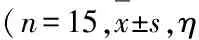

GroupRatio of iNKT cellsThymusSpleenLiverLymph nodesWTmice0.28±0.081.07±0.5514.92±6.380.14±0.04HDAC3KOmice 0.02±0.01∗0.01±0.01∗0.08±0.04∗0.01±0.01∗
*P<0.05 compared with WT mice.
表2 HDAC3KO和WT小鼠胸腺、脾脏、肝脏和淋巴结中iNKT细胞数量
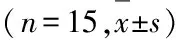

GroupNumber of iNKT cellsThymus(×105)Spleen(×105)Liver(×104) Lymph nodes(×104)WT mice3.79±1.0611.98±4.8824.25±10.371.16±0.28HDAC3KO mice0.21±0.06∗0.15±0.07∗0.22±0.10∗0.06±0.04∗
*P<0.05 compared with WT mice.
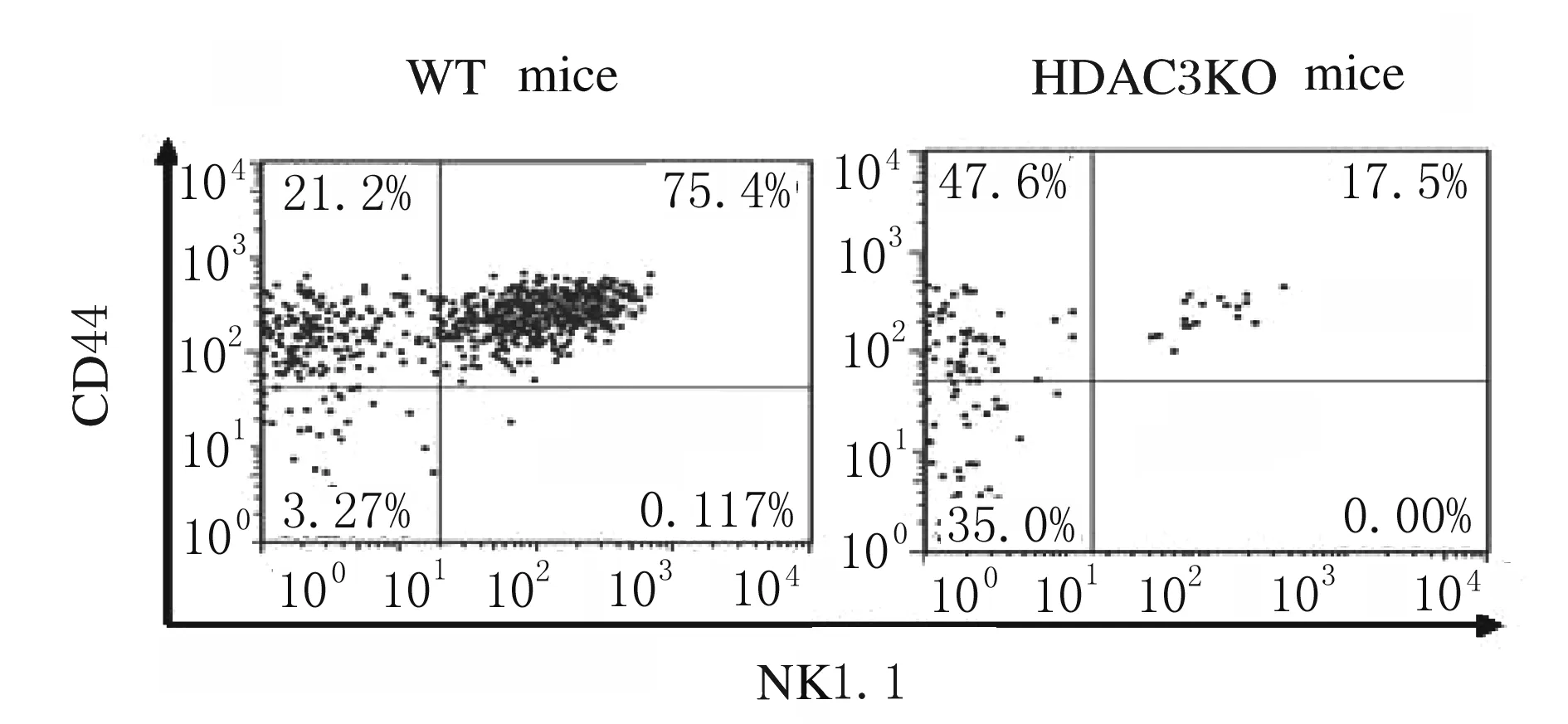
图3 流式细胞术检测HDAC3KO和WT小鼠胸腺iNKT细胞中CD44和NK1.1的表达
Fig.3 Expressions of CD44 and NK1.1 in thymus iNKT cells of HDAC3KO and WT mice detected by flow cytometry
*P<0.05 compared with WT mice.
图4 HDAC3KO和WT小鼠胸腺不同发育阶段iNKT细胞比例
Fig.4 Ratios of iNKT cells in thymus in different development stages of WT and HDAC3KO mice
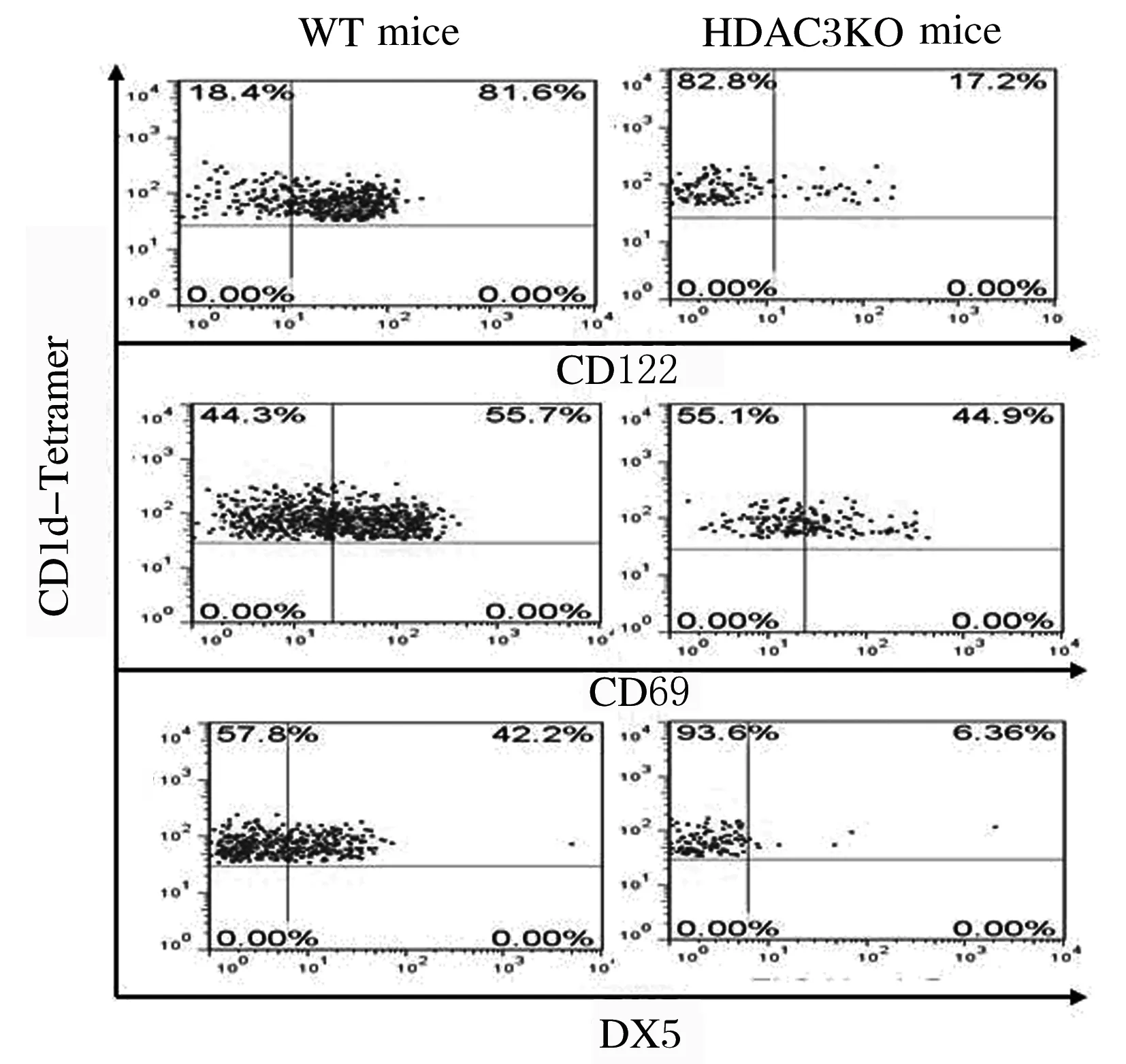
图5 流式细胞术检测HDAC3KO和WT小鼠胸腺iNKT细胞中CD122、CD69和DX5表达
Fig.5 Expressions of CD122,CD69 and DX5 in thymus iNKT cells of HDAC3KO and WT mice
2.4 HDAC3通过内源性途径对iNKT细胞发育和成熟的调控作用流式细胞术检测HDAC3KO小鼠DP细胞中CD1d表达结果显示:与WT小鼠比较,HDAC3KO小鼠DP细胞中CD1d表达无明显变化。见图7。采用骨髓混合嵌合体方法检测受者小鼠胸腺iNKT细胞的产生和发育结果显示:与来源于正常小鼠的骨髓细胞比较,来源于HDAC3KO小鼠的骨髓细胞产生的iNKT细胞数量明显减少(P<0.05)。发育表型检测结果显示:与来源于正常小鼠的骨髓细胞比较,来源于HDAC3KO小鼠骨髓的iNKT细胞主要表现为非成熟CD44-NK1.1-和CD44+NK1.1-表型,而成熟CD44+NK1.1+表型细胞比例明显降低(P<0.05)。见图8~11。
2.5 HDAC3敲除小鼠iNKT细胞和血清中IFN-γ和IL-4水平细胞内染色结果显示:α-Galcer体内刺激4 h后,与WT小鼠比较,HDAC3KO小鼠iNKT细胞中IFN-γ和IL-4水平明显降低(P<0.05)。见图12和图13。ELISA法检测显示:与WT小鼠比较,HDAC3KO小鼠血清中IFN-γ和IL-4水平明显降低(P<0.05)。见图14。

*P<0.05 compared with WT mice.
图6 HDAC3KO小鼠胸腺iNKT细胞中CD122、CD69和DX5表达水平
Fig.6 Expression levels of CD122, CD69 and DX5 in thymus iNKT cells of HDAC3KO mice

图7 流式细胞术检测HDAC3KO和WT小鼠DP细胞中CD1d的表达
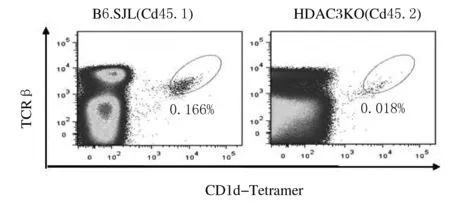
图8 流式细胞术检测来源于HDAC3KO和B6.SJL小鼠骨髓细胞的iNKT细胞的产生
Fig.8 Production of iNKT cells from bone marrow cells of HDAC3KO and B6.SJL mice detected by flow cytometry
3 讨 论
本研究结果显示: HDAC3对于iNKT细胞的发育和细胞因子的产生发挥关键作用。HDAC3敲除导致中枢和外周免疫器官中iNKT细胞数量明显减少,iNKT细胞在胸腺的发育受阻,同时iNKT细胞因子水平也明显降低。

*P<0.05 compared with B6.SJL mice.
图9 来源于HDAC3KO和B6.SJL小鼠骨髓细胞的iNKT细胞比例
Fig.9 Ratios of iNKT cells from bone marrow cells of HDAC3KO and B6.SJL mice
本文作者以往研究[10]显示:CD4Cre酶介导的HDAC3敲除不影响传统T细胞在胸腺的发育和成熟,但导致外周CD4+T细胞和CD8+T细胞出现活化诱导的细胞凋亡,表现为细胞核DNA复制加快但同时伴随着细胞凋亡的明显增加。另外,增殖能力旺盛的造血干细胞缺失HDAC3引起多能淋巴祖细胞凋亡增加,淋巴细胞生成明显减少[11]。鉴于HDAC3在T淋巴细胞活化增殖中的重要性,HDAC3已被作为靶点用于T细胞淋巴瘤的临床治疗[12]。另外,iNKT细胞由胸腺DP细胞识别邻近细胞CD1d提呈的糖脂类抗原后经阳性选择产生,在其发育和成熟过程中,iNKT细胞中CD44和CD69等T细胞活化分子表达水平明显升高,增殖活性也明显增强。本研究结果显示:HDAC3敲除主要导致CD44+NK1.1+成熟iNKT细胞比例明显降低,而非成熟CD44-NK1.1-和CD44+NK1.1-iNKT细胞比例升高,提示T细胞活化增殖越明显,其活性对HDAC3的依赖性越强。另外,THAPA等[13]采用PLZFCre酶介导的基因敲除方法,在iNKT细胞发生阳性选择后敲除HDAC3,也观察到iNKT细胞数量在胸腺、脾脏和肝脏中明显减少。然而,CD4Cre酶介导的hdac3基因敲除导致iNKT细胞数量在胸腺和外周免疫器官中明显降低,表明HDAC3对于iNKT细胞的阳性选择以及整个发育和成熟过程必不可少。

图10 流式细胞术检测来源于HDAC3KO和B6.SJL小鼠骨髓细胞的iNKT细胞中CD44和NK1.1的表达
Fig.10 Expressions of CD44 and NK1.1 in iNKT cells from bone marrow cells of HDAC3KO and B6.SJL mice detected by flow cytometry
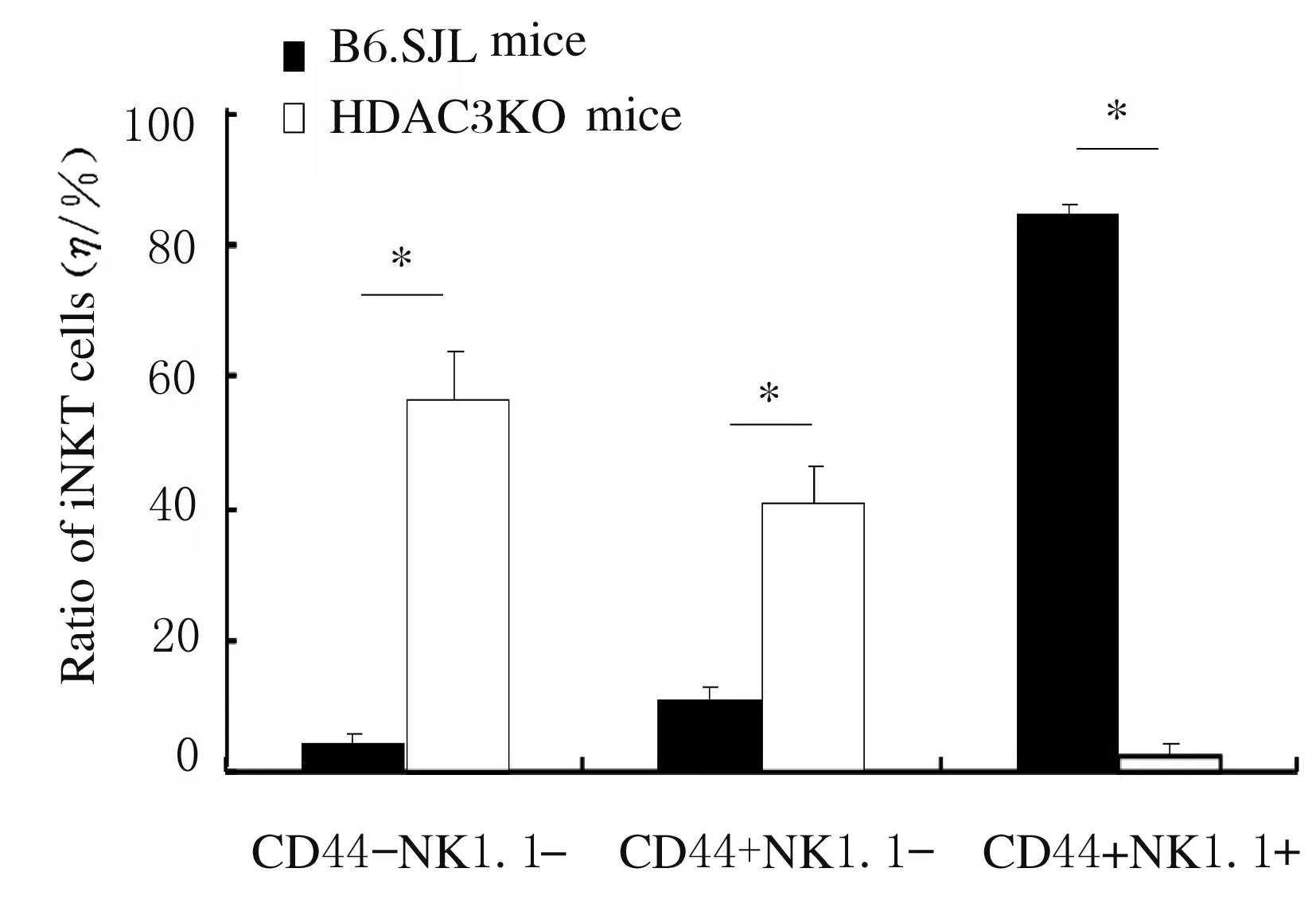
*P<0.05 compared with B6.SJL mice.
图11 来源于HDAC3KO和B6.SJL小鼠骨髓细胞的不同表型iNKT细胞比例
Fig.11 Ratios of iNKT cells with different phenotypes from bone marrow cells of HDAC3KO and B6.SJL mice

图12 细胞内染色检测HDAC3KO和WT小鼠iNKT细胞中IFN-γ和IL-4表达
Fig.12 Expressions of IFN-γ and IL-4 in iNKT cells of HDAC3KO and WT mice detected by intracellular staining

*P<0.05 compared with WT mice.
图13 HDAC3KO和WT小鼠iNKT细胞中IFN-γ和IL-4表达水平
Fig.13 Expression levels of IFN-γ and IL-4 in iNKT cells of HDAC3KO and WT mice
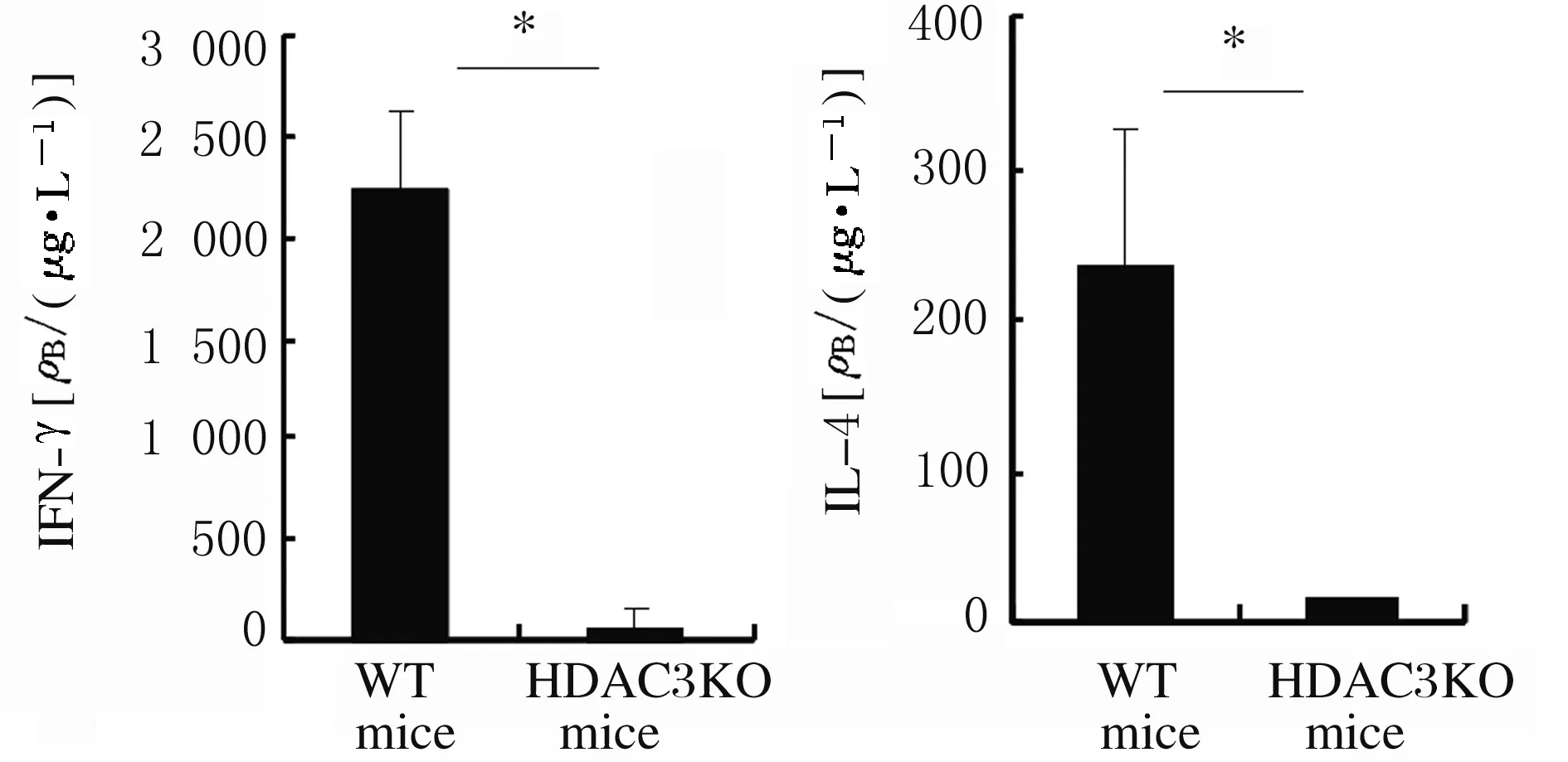
*P<0.05 compared with WT mice.
图14 ELISA法检测HDAC3KO和WT小鼠血清中IFN-γ和IL-4表达水平
Fig.14 Expression levels of IFN-γ and IL-4 in serum of HDAC3KO and WT mice detected by ELISA method
iNKT细胞在胸腺的产生和发育既依赖于胸腺微环境,也依赖于其细胞自身。胸腺DP细胞表面CD1d对糖脂类抗原的有效提呈在iNKT细胞阳性选择过程中发挥重要作用,CD1d敲除小鼠iNKT细胞几乎完全缺失[14]。然而,本研究结果显示:HDAC3敲除并不影响胸腺DP细胞表面CD1d的表达,提示HDAC3主要通过内源性途径调控iNKT细胞在胸腺的产生、发育和成熟,这也被骨髓混合嵌合体实验结果进一步证实。这一结果表明:以HDAC3为靶点的干预措施可直接对iNKT细胞发育和功能发挥调控作用,进而可以调节iNKT相关疾病的发生发展。
目前对HDAC3调控iNKT细胞产生、发育的信号通路仍不完全明确。CD122是IL-15受体β亚单位,本研究结果显示:CD122在HDAC3敲除小鼠iNKT细胞中表达水平明显降低,表明HDAC3敲除引起的iNKT细胞产生和发育缺陷与IL-15信号通路受阻有关[15]。ZHU等[16]报道IL-15信号途径缺陷导致iNKT细胞自噬功能下降,细胞生存能力明显降低。THAPA等[17]研究显示:与正常iNKT细胞比较,HDAC3缺失iNKT细胞自噬标记物Cyto-ID染色阳性率和LC3A/B表达降低,进一步证实HDAC3可通过细胞自噬途径调控iNKT的生存和发育。另外,HDAC3与NKAP和Runx1均可发生细胞内相互作用,而NKAP和Runx1缺失同样导致iNKT细胞生成明显减少,提示HDAC3也可通过NKAP和Runx1相关信号途径调控iNKT的产生和发育[18-19]。
在受到TCR信号刺激时,iNKT细胞能在短时间内产生大量Th1和Th2型细胞因子,进而发挥强大的免疫调节功能[20]。研究[21]显示:非成熟CD44-NK1.1-和CD44+NK1.1-iNKT细胞主要产生IL-4,而成熟CD44+NK 1.1+ iNKT细胞主要产生IFN-γ。本研究结果显示:HDAC3敲除导致iNKT细胞和机体IL-4和IFN-γ的产生均明显降低,表明HDAC3可总体影响iNKT细胞的功能。一方面,HDAC3缺失引起的iNKT及外周CD4+和CD8+T细胞数量的明显减少可导致α-Galcer活化iNKT细胞和机体细胞因子产生明显减少;另一方面,HDAC3也可能通过对IL-4和IFN-γ及其转录因子如GATA-3和T-bet的直接或间接表达调控作用影响以上细胞因子的产生[8]。
综上所述, HDAC3内源性地调控iNKT细胞的发育和成熟,HDAC3缺失导致iNKT细胞数量明显减少、细胞成熟障碍和细胞因子产生功能的明显降低。而对于HDAC3调控iNKT细胞产生、发育和功能发挥的信号通路目前仍不明确,需要在后续实验中进一步探讨。
