湖相介形类壳体微量元素在古环境重建中的应用
2020-04-20姜高磊刘林敬毛欣
姜高磊,刘林敬,毛欣
1. 中国地质科学院水文地质环境地质研究所,石家庄 050061
2. 中国地质科学院第四纪年代学与水文环境演变重点实验室,石家庄 050061
介形类是海洋、湖泊、河流等水域中广泛存在的一种节肢类小型甲壳动物,以浅海和静水湖泊中最为丰富[1-3]。湖泊中的介形类一般营底栖生活,栖息于沉积物表面、沉积物内部(钻掘深度通常<1 cm)或者水草丛中,取食各种有机质,包括藻类、原生动物、植物残体等[1-2]。
大多数介形类的寿命短于一年,多数为几周至几个月。介形类以蜕壳的方式进行生长,由卵孵化成幼虫称为第一龄期,当身体生长对壳体来说已经过大时,坚硬的双壳脱落,随之身体迅速增长和发育,同时形成新的壳并钙化,其完整的生命周期要经历8 个这样的龄期,蜕壳8 次,在沉积物中留下9 对钙质外壳[1,4]。
介形类壳体大小多数为0.5~2 mm,主要由低镁方解石组成,且壳体的组成物质几乎全部来自于其生存的水体[5]。介形类每次蜕壳,新壳壳体的钙化时间特别短,一般为几个小时至几天时间[6],在钙化过程中介形虫吸收宿生水体中的离子进行壳体的钙化,介形虫除了吸收壳体主要组成物质Ca 离子、碳酸根离子之外,还会吸收一些其他微量元素,如Mg、Sr、Ba 等(图1),且这些元素的含量及其之间的比值受水温、水化学参数等因素的影响[7-10]。由于不同物种具有特定的生存微环境,且对其宿生水体的水化学条件的变化极其敏感,而且低镁方解石相对比较稳定,可以在沉积物中长期保存,不易受后期成岩作用的影响,介形类壳体中的微量元素及其比值通常被作为重建古环境、古水文的重要指标[11-19]。

图1 介形类壳体微量元素比值(中心介形类为意外湖花介左壳;据文献[8]修改)Fig.1 Trace element ratio of an ostracod shell (a left valve of Limnocythere inopinata in the center; modified from reference[8])
湖湘介形类壳体微量元素在重建古环境中的应用始于20 世纪80 年代[20-23],在应用过程中出现了一些问题,方法也在不断地完善和修正。本文在文献研读的基础上,尝试总结前人利用介形类壳体微量元素(主要指Mg 和Sr)定量重建古环境工作中的成果和遇到的问题,对未来研究工作进行展望。
1 研究历史及应用原理
在20 世纪80 年代,Chivas 等[20]提出非海相介形类壳体中的Mg、Sr 和Ba 微量元素与其宿生水体中对应的微量元素成比例,壳体中的微量元素含量可以用来进行定量恢复其宿生水体的微量元素组成。之后,Chivas 等[21]利用澳大利亚东南部的Keilambete 湖泊中现生介形类Australocypris robusta壳体中Sr/Ca 比值和湖水中Sr/Ca 比值计算出其壳体的Sr 分配系数Kd为0.082。计算公式如下:

式中:Sr/Ca 比值为摩尔比。利用计算得出的分配系数和湖泊沉积物中的介形类壳体中的Sr/Ca 比值,定量恢复了Keilambete 湖一万年以来湖水盐度变化。但在之后一年,Chivas 等[22-23]在介形类培养试验和野外调查研究过程中发现,介形类壳体中的Mg 的分配不仅与宿生水体的Mg/Ca 比值有关,而且受温度和个体生长阶段的影响,而Sr 的分配与宿生水体中的Sr/Ca 和盐度有关[7,24]。之后越来越多的学者注意到其他地区介形类壳体中Mg 的含量也受宿生水体温度的影响[25-27],而且温度对介形类壳体中Mg/Ca 比值的影响量很大,20 ℃的温度变化可以引起Mg/Ca 比值50%的变化[28]。因此,学者们将介形类壳体中Mg 的分配系数计算公式表达如下:

式中:Mg/Ca 比值为摩尔比。当介形虫宿生水体中的Mg/Ca 比值较为恒定时,壳体中的Mg/Ca 比值可以反映宿生水体的温度变化,这也是利用海水介形类壳体中Mg/Ca 比值定量重建湖水[29]或海水[30-31]温度的基本原理。
利用上述介形类壳体中微量元素与宿生水体中的微量元素的关系可以定性或者定量地进行古环境重建。定性重建古环境主要是利用介形类壳体中微量元素比值M/Ca 随着湖泊沉积物的深度(时间)序列的变化趋势,定性地描述湖泊演化过程中湖泊水化学的变化趋势。如,Zhai 等[15]在呼伦湖利用介形类(意外湖花介Limnocythere inopinata)壳体中的Mg/Ca 和Sr/Ca 比值结合其他指标定性地重建了11 000 年以来湖泊水文演化历史。定量重建古环境的基本流程如下:首先通过调查现生湖泊中介形类壳体中M/Ca(M 指Mg 和Sr)比值与湖水的M/Ca 比值确定现生介形类壳体的分配系数Kd[M];与此同时需要调查研究现代湖水中的M/Ca 比值与古环境指标(多指古盐度)的关系,建立定量关系式。然后利用沉积物中的介形类壳体化石中的M/Ca 比值和已经获得的分配系数Kd[M],推算得出古湖水中的M/Ca 比值,代入M/Ca 比值与古环境指标的关系式进行定量重建[16]。依据以上原理,在之后的二三十年里,这种方法在很多地区广泛使用进行古环境的定量重建,如牙买加的Wallywash Great Pond[32]、澳大利亚的中部Frome 湖泊[33]、意大利南部的西西里岛湖泊[17]以及我国的内蒙古岱海[11]、新疆博思腾湖[34]。
随着研究的深入,在利用介形类壳体微量元素进行古环境重建的过程中,部分学者发现重建的结果与其他指标存在较大的偏差。如在云南鹤庆盆地古环境研究中发现,当沉积物中有自生碳酸盐出现时,介形类壳体中的Sr/Ca 比值不能很好地响应环境变化[35]。在苏干湖的研究中发现,当介形类宿生水体中的Mg/Ca>40 时,壳体中的Mg/Ca 比值较为复杂[36]。利用介形类壳体微量元素重建古环境的影响因素较为复杂,许多学者通过室内培养和野外调查研究,具体影响因素见下文。
2 影响因素
根据介形类壳体微量元素重建古环境的原理,将其影响因素分为两类:影响介形类宿生水体中Mg/Ca、Sr/Ca 比值的因素以及影响介形类壳体微量元素分配系数的因素。
2.1 宿生水体中M/Ca 比值变化的影响因素
从上述原理可以看出,利用介形类重建古环境主要是通过壳体的M/Ca 比值来推断或计算湖泊水体的M/Ca 比值,进而推断环境变化。但在实际过程中有些影响因素可以造成湖泊的一些水文条件的变化并不能通过水体的M/Ca 比值反映出来,或者介形类壳体的M/Ca 比值变化并不是环境变化导致的。主要有以下因素:
2.1.1 季节变化
通过逐月采集北美大平原上湖泊中介形虫(Candona rawsoni)进行介壳种Mg、Sr 含量测定,结果发现一年内的单个个体之间介壳Mg、Sr 元素含量的变化竟然可以达到35%,湖水中Mg、Sr 浓度随着季节的变化直接导致了介壳中Mg/Ca、Sr/Ca 比值随季节的变化[37]。在对地中海地区一处湖沼研究时发现,水体的碱度、Ca2+浓度、Sr/Ca 和Mg/Ca比值均具有季节变化,变化幅度约为10%,而且能体现在介形类壳体的微量元素组成信号上[38]。因此,在利用介壳中Mg/Ca、Sr/Ca 比值重建古环境时,同一层位上分散多采样品,避免由于季节变化造成的差异[37]。近年来,由于技术的发展,在单壳上进行Mg/Ca、Sr/Ca 比值测试得以实现,这种方法能够让我们更精确地分析介形类壳体上微量元素的分布[39],但在重建古环境时也需要注意季节变化对单个个体的影响[40]。
2.1.2 生存微环境差异
在进行云南鹤庆盆地的古环境恢复时,胡广等[35]发 现 利 用Ilycypris microspinata 和Lineocypris jiangsuensis 两个属种的重建结果具有很好的协同性,但是二者之间存在稳定的差值,分析认为二者生活水域的差异,水体微环境化学组成可能导致生活其间的壳体化学组成不同,以底栖方式生活的L. jiangsuensis 的壳体中的Mg/Ca、Sr/Ca 比值较I.microspinata 的值高。
2.1.3 湖泊演化过程(矿物沉淀)
在重建湖水古盐度的过程中,会将恢复的水体的M/Ca 比值与盐度联系起来,但是在许多湖泊演化过程中湖水的离子浓度和比值与盐度并不是简单的线性关系[41](图2),只有在特殊的环境下,如钙亏损的湖泊,水体中的M/Ca 与盐度才具有正相关关系[42]。如对地中海地区的一处湖沼进行逐月采样分析发现,介形虫Herpetrocypris intermedia 壳体中的Sr/Ca 和Mg/Ca 比值随着盐度的增加而降低[38]。关于湖泊水体演化路径可以查阅文献[43],湖泊演化过程对利用介形类重建古环境的影响在文献[41]中做了详细的论述。当湖水咸化沉积物中出现文石沉淀时,水体中的Sr/Ca 比值会急剧下降,导致介形类壳体中的Sr/Ca 比值不能真实地反映湖泊环境变化[35--36]。黏土矿物的沉淀,尤其是含Mg 的蛭石和蒙脱石会选择性地吸收水体中的Mg,使得水体中的Mg 含量降低,也影响到重建结果[44]。
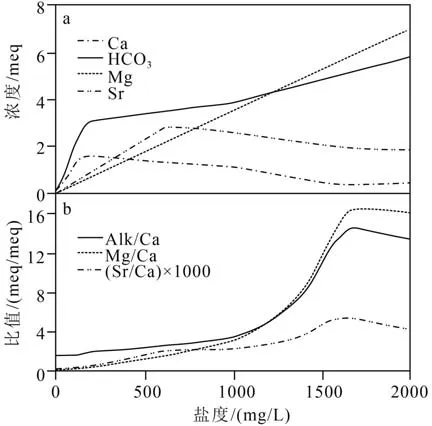
图2 湖泊水体中离子浓度和比值与盐度关系(据文献[41]修改)Fig.2 Ion concentration and ratio change with salinity in lake water (modified from reference[41])
2.2 影响分配系数的因素
从利用介形类壳体微量元素定量重建古环境的原理和过程可以看出,分配系数是十分重要的。多数学者在根据现代湖泊调查或者室内培养实验得出分配系数后将其应用到古环境的定量重建[11,17],但在研究过程中发现介形类的微量元素分配系数并不是恒定的,而是受其生存环境影响会发生变化,影响因素主要有以下几种:
2.2.1 宿生水体中的M/Ca 比值
在一定的范围内,介形类壳体的M/Ca 比值会随着宿生水体中的M/Ca 比值增大,这也是利用介形类重建古环境的基础,但是当水体中的M/Ca 比值过大或者过小的时候,壳体的Kd[M] 会出现偏差。如随着水体中的Mg/Ca 比值增加,介形类壳体中的Mg/Ca 比值增加的速度会减缓以至于不增加,也就是Kd[M]值会变小(图3a),认为介形类在高镁水体中分泌形成低镁方解石壳体时,壳体吸收的Mg(和Sr)的量是有阀值的,过多的Mg 会造成介形类生理上的排斥[26,37]。Zhang 等[36]在利用介形类重建苏干湖古环境演化时,发现当水体Mg/Ca 比值>45 时,Eucypris inflata 的Kd[Mg] 值为0.000 3,要比先前其他属种的研究中Kd[Mg] 值低一个数量级。当水体中Mg/Ca 比值过低时(<2),介形类壳体Kd[Mg]会随着水体中Mg/Ca 比值的降低迅速增加(图3b),Kd[Sr]十分分散(图3c)。从晶体学和介形类壳体钙化过程的角度来说,Mg2+和Ca2+的离子半径相差较大,Mg 不能以完全类质同像的方式进入方解石壳体,当水体中Mg/Ca 较高时,一般湖水的盐度也较大,水体中的Ca2+离子充足,介形类在壳体生长过程中会更倾向于更易吸收的Ca2+,导致壳体中的Mg/Ca 比值偏低;当水体中Mg/Ca 比值较低时,湖水盐度也较低,两种离子的浓度均较低,介形类为了形成新的壳体,会将Mg 以类质同像的方式吸收,导致壳体中的Mg/Ca 比值偏高[35]。而介形类壳体Kd[Mg]随着水体中Mg/Ca 比值增加而降低是介形类自生在高Mg 环境中在生理上对Mg 的吸收能力和在低Mg 环境中吸收Mg 的热动力学平衡之前的一个权衡,也就是说介形类会在尽量消耗低能量的情况下完成壳体的分泌、钙化[45]。
2.2.2 温度
一定的温度、盐度范围内,介形类壳体Kd[Mg]值 随着温度的升高而增加[7,23, 25-26,47]。早期研究[23]发现,在11~25 ℃时,温度每升高1 ℃,Mytilocypris hericae 壳体的Mg/Ca 比值增加0.0015,导致壳体Mg的分配系数升高。Chivas 等[21,23]在1985 和1986 年的论文中均认为介形类壳体Kd[Sr]不受温度变化的 影 响,但De Deckker 等[7]通 过 介 形 类Cyprideis australiensis 培养实验,认为宿生水体的盐度并不影响壳体对Mg、Sr 的吸收,而温度不仅与壳体Kd[Mg]具有正相关,而且对Kd[Sr]有一定的影响。Dettman等[9]在总结前人资料的过程中,将Chivas 等[21,23]的数据进行了整理,发现在水化学条件相同的水体中,壳体中的Sr/Ca 比值是随着温度的升高而升高的,因此,他们认为温度对介形类壳体Kd[Sr]虽然影响较小,但还是有一定的影响(图4)。
2.2.3 碱度
Gouramanis 等[45]在对澳大利亚一系列的咸水、超咸水湖泊中现生介形类研究发现,高碱度(碳酸氢根离子活度)对介形类壳体的Kd[Sr]影响很大,较高的碱度会导致Kd[Sr]显著下降。
以上几种影响因素是通过直接影响介形类壳体的分配系数来影响古环境的重建,这些影响因素最终是影响了介形类壳体分泌、钙化过程中的生命-化学过程,是利用介形类定量重建古环境的重要障碍,只有通过足够多的现代过程研究,充分了解不同环境下介形类壳体的分泌、钙华过程,才能保证古环境重建结果的可靠性。值得一提的是,以上有关影响因素和古环境重建过程中所用的介形类均是成体,不同发育阶段的介形虫壳体中的微量元素组成是不同的,一般发育早期的壳体中Mg 的含量偏高[22, 27, 48],钙化程度较弱的壳体中的Sr 含量偏低[49]。
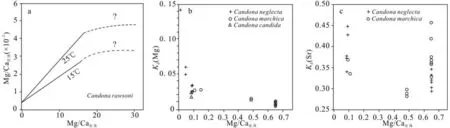
图3 介形类壳体Mg/Ca 比值、Kd [Mg]与湖水Mg/Ca 比值关系(a 据文献[37]修改;b 和c 据文献[46]修改)Fig.3 The relationship between Mg/Ca ratio, Kd[Mg] of shell and Mg/Ca ratio of lake water(a modified from reference[37]; b and c modified from reference[46])
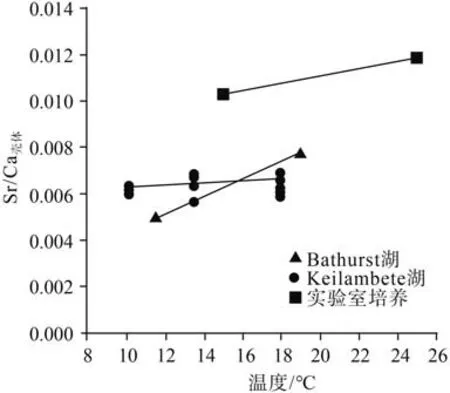
图4 介形类壳体Sr/Ca 与温度的关系(据文献[9]修改)Fig.4 The relationship between Mg/Ca ratio and temperature (modified from reference[9])
由于上述众多影响因素,介形类壳体的微量元素分配系数变化很大,即使是同一属种在不同的环境中也存在很大的差异。下面以我国西北和北方常见属种意外湖花介(Limnocythere inopinata)为例进行讨论,意外湖花介在我国西北和北方干旱、半干旱地区的现代湖泊和新生代地层中大量出现[50-54],该种的生活习性研究也较多[4,55-56]。在利用意外湖花介壳体Sr/Ca 比值定量重建岱海古盐度的研究中,根据现代壳体Sr/Ca 比值和湖水Sr/Ca 比值计算得出分配系数为1.064[11],而日内瓦湖内的分配系数平均值为0.306[47],澳大利亚湖泊中计算的现代湖花介属Sr 的分配系数为0.35[22]。前人分别研究了日内瓦湖泊[47]和蒙古国西部湖泊[42]的意外湖花介在内的介形类壳体的地球化学与水体化学参数的关系,本文将涉及意外湖花介壳体微量元素组成与水体中M/Ca 比值的数据重新整理,得到分配系数与水体M/Ca 比值的关系如图5。由图5 可以看出,L.inopinata 壳体的Kd[M]随着宿生水体的M/Ca 变化较大,尤其是在M/Ca 较低的水体中,还存在其他的影响因素,甚至这种影响因素占到了主导地位。介形虫壳体中的Mg 和Sr 具体在壳体方解石中的哪个位置,对于壳体的形成有什么作用还不很清楚。在矿物方解石的形成过程中,Mg 十分重要,具有动力学和热力学上的控制作用[57-58]。因此,介形类壳体中的Mg 和Sr 的含量可能存在生物学和/或矿物结晶学方面的控制因素,但目前还不是很清楚。
3 存在问题及展望
经过几十年的发展和完善,利用介形类壳体的微量元素进行古环境重建取得了很大的进展,但也存在着一些问题:

图5 意外湖花介壳体Kd[M]与水体中M/Ca 比值关系(数据来源于文献[11, 42, 47])Fig.5 The relationship between Kd [M] of Limnoctyhere inopinata shell and M/Ca ratio of lake water (data from reference [11, 42, 47])
不同学者在对介形类壳体中微量元素的影响因素研究得到了一些相矛盾的结论,如一些学者认为温度对介形类壳体中的Mg/Ca 比值影响很大[25,41],而另一些学者认为温度对壳体中Mg 的含量几乎没有任何影响[59],这可能是由于不同属种的介形虫具有不同的生活习性,有些属种属于浮游类,有些属种属于底栖类,其壳体的元素组成代表了不同的水体微环境信息;有些生命周期几周至几个月,有些能够存活一两年,其壳体的元素组成代表了不同季节或者不同时间的水体信息[9]。
在定量重建过程中,多数学者在一套沉积物序列中使用一个固定的分配系数,如上文所述,介形类壳体的微量元素分配系数并不是恒定的,即使是同一属种在不同的环境下变化也很大。一些研究中定量重建研究时间跨度上千年甚至上万年,沉积环境复杂多变,利用同一个分配系数进行定量重建难免会增大误差。介形类生存水域广阔,属种繁多,是重建古环境的良好材料,但生存环境和属种的多样也给重建工作带来了复杂性,如上述两个问题。建议在以后的研究工作中加强现代过程的研究,对于不同属种的介形虫应在生活习性和壳体微量元素组成的影响因素两个方面加强现代过程研究。
经过30 余年的发展,湖相介形类壳体微量元素组成及比值已然成为了重建古环境的重要指标。随着测试技术的发展,现在已经能够对壳体的不同区域和同一位置不同厚度进行元素测试[39-40],这对以后更加精细地重建古环境提供了基础。另外,除了常用的Mg/Ca、Sr/Ca 比值,湖相介形类壳体Gd/Ca 和Mn/Ca[10]、Ba/Ca和U/Ca等[40]、Li/Ca和Mg/Li[60-61]、Na/Ca[62]也更多地在古环境研究中使用。随着技术的提高和研究的深入,湖相介形类壳体微量元素组成会有更加广泛的应用。
