不同种源羊草表型差异性
2020-04-16魏继平马红媛
李 阳,魏继平,马红媛,*
1 中国科学院东北地理与农业生态研究所,长春 130102 2 中国科学院大学,北京 100049
植物的性状与环境有着紧密的关系,它是植物在特定环境下生存策略的重要表现[1- 2]。在当今全球剧烈的气候条件变化下,植物的生存环境在时间和空间上不断变化着,对于一些广泛分布的物种来说,植物通常会通过调整表型特征来增加自身的适合度,以削弱不良环境对生长、繁殖造成的不良影响,造成不同地区植物表型的差异[3- 5]。表型分化不仅可以反映生存环境的质量,也可以反映出物种对于环境的忍受力。从物种进化角度来看,变异是生物进化的基础,有利于物种种内、种间竞争,提升物种在自然群落中的生态位。因此,探索物种表型分化对于植物物种适应环境变化来具有重要的作用[6- 9]。种群种内的多样性有以下两个原因:表型可塑性和遗传变异。前者由于环境变化某个特定基因有着不同表型,后者为自然选择压力下的遗传变异[10- 12]。物种在适应环境中表现出的这两种生长策略是不矛盾的,同一物种适应即可以有表型可塑性又可以有遗传分化,这与它们生存环境相关[13]。对物种表型分化差异的研究不仅能够了解它对环境的适应策略,而且能够对未来物种进化、发展进行预测。
羊草(Leymuschinensis)是多年生根茎型植物,具有适口性好、蛋白质含量高和耐盐碱等特点,在世界范围内具有广泛的地理分布,并在发展草原经济建设中占有重要地位[14]。近些年来关于羊草研究表明,不同种源的羊草在形态、单穗长度、小穗数、过氧化物歧化酶(POD,Peroxidase)和超氧化物歧化酶(SOD,Superoxide dismutase)酶活性等特征中存在差异,经过同地栽培后,这种差异在后代中也体现出来[15- 18]。这种传递到后代的表型差异还可以通过遗传同化的过程保持稳定[1,19],增强后代环境适应能力。Wang等的研究发现,羊草总植株密度、营养枝、生殖枝生物量随着降水量的增加而增加,随干旱指数的增加而下降。得出了营养生长和生殖生长可能是两种生态型产生的原因[20]。Yuan等对灰绿型羊草和黄绿型羊草的研究发现,灰绿色羊草有着比黄绿色羊草更强的适应性,在茎高、穗长、叶面积、种子重量等方面存在显著差异,并认为这与年均温、年降水量和土壤水分含量相关[11]。Guo等在中国东北沿大尺度干旱梯度的研究表明,羊草的叶片厚度随着干旱程度的增加而增加[21]。因为更大的叶片厚度可以为水分储存提供结构基础,并在进化角度上有利于有效水分利用。我们近期研究发现,GA3引发羊草种子存在可以遗传的表型差异,在子一代和子二代中稳定发生[22]。
以往的研究都主要集中在个别的环境因子或者控制试验上,而在同质园移栽条件下观察羊草表型差异的报道较少。本研究选取4个种源地的羊草为研究对象,开展同质园试验,分析其对环境适应特性,阐明不同种源的羊草对其生长环境是否具有较高的表型差异?这种表型差异是由遗传因素还是环境因素造成?
1 研究方法
1.1 不同种源羊草采集
本研究所在的同质园试验地位于中国科学院东北地理与农业生态研究所(43°5′—45°15′ N,124°18′—127°2′ E)。于2017年4月末羊草返青时期,分别在吉林省松嫩平原西部的镇赉(ZL),洮北区(TB),大安市(DA)和大安市姜家甸草场(JC)种源选取羊草,5月初进行田间单枝移栽。共分为4个实验小区,每个小区设置8个重复样,小区之间设有保护行,定期浇水、除草,每次浇水量保持一致。长春市年平均气温4.9 ℃,年降水量593.8 mm,属于温带大陆性半湿润季风气侯类型。
1.2 光合测定
在2017年8月,选择晴朗无云天气,使用Li- 6400便携式仪进行测定。测定时间为9:00—11:00,每个种源选取6枝长势均匀的羊草作为研究对象,选取羊草叶片顶端第二片展开叶进行测定,3次重复,取平均值。由Li- 6400便携式光合作用测定仪测出各项数据,测定指标包括:净光合速率Pn(μmol m-2s-1)和蒸腾速率Tr(mmol m-2s-1)。测定的数据经计算得到水分利用效率(WUE)。WUE(μmolCO2/mmolH2O)=Pn/Tr。
1.3 比叶面积的测定
将采集的叶片用Li- 3000型便携式叶面积仪进行叶面积的测定,将不同种源地的叶片取回到实验室,用Li- 3000型便携式叶面积仪测定叶面积(S),每个种源地取6枝羊草,对所有叶片测定叶面积;之后,将羊草叶片放在105 ℃杀青2 h,然后在85℃下烘24 h至恒重,用1/10000g电子天秤称重(W)。比叶面积(SLA)用以下公式进行测定:SLA(cm2/g)=S/W。
1.4 表型性状的测定
利用刻度尺测量羊草的株高、叶片长度;用游标卡尺测定羊草的叶宽、茎直径等指标。
1.5 数据分析
以每枝羊草为统计单位,各种源羊草株高、叶长等平均值代表该种源的总体水平。使用RStudio软件(R- 3.3.3版本)进行单因素方差分析(ANOVA)。在进行不同种源羊草性状特征比较时,采用Tukey进行多重比较。在数据分析之前,数据的正态性(shapiro-wilk)和方差齐性(LeveneTest)都进行了检验,必要时数据需进行Log转换等以满足正态性和方差齐性的条件。最后使用Origin 9.0进行绘图。
2 结果与分析
2.1 不同种源间羊草生长特性比较
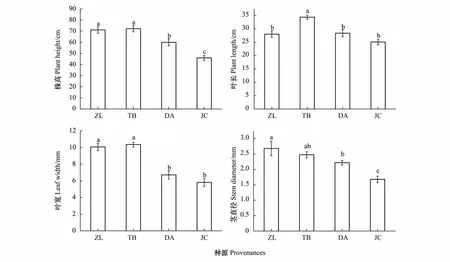
图1 不同种源羊草叶片表型比较Fig.1 The comparison of leaf traits between provenancesZL:代表镇赉种源地羊草;TB:代表洮北种源地羊草;DA:代表大安种源地羊草;JC:代表姜家甸草场种源地羊草;不同小写字母表示差异显著(P<0.05)
来自4个种源地的羊草株高、叶长、叶宽以及茎直径均存在显著差异(图1和图2)。统计分析结果表明,4个种源地羊草的株高存在显著差异(df=3,F=19.3,P<0.05),其中ZL和TB均显著高于DA和JC种源地,但ZL和TB之间无显著差异(图1)。4个种源地羊草叶长存在显著差异(df=3,F=12.48,P<0.05),其中TB种源地叶长显著高于其他三个种源,而其他三个种源羊草叶长没有显著差异(图1)。4个不同种源地羊草叶宽存在显著差异(df=3,F=29.32,P<0.05),ZL和TB种源地羊草叶宽均显著高于DA和JC种源地(图1)。不同种源茎直径之间存在显著差异(df=3,F=9.15,P<0.05)。ZL种源地茎直径最高,且显著高于DA和JC种源(图1)。

图2 不同种源羊草生长情况,每个种源羊草选取六枝 Fig.2 Image of Leymus chinensis growth. Six plants were selected for each provenance
2.2 不同种源间叶、鞘和茎生物量比较
4个种源地羊草叶的鲜重存在显著差异(df=3,F=6.144,P<0.05),其中ZL和TB种源显著高于其他两个,DA与JC种源之间无显著差异(图3)。4个种源地羊草鞘的鲜重存在显著差异(df=3,F=13.77,P<0.05),各种源鞘鲜重表现为:ZL>TB>DA>JC,其中ZL种源显著高于其他3个种源,而其他3个种源间差异并不显著(图3)。4个种源地羊草茎的鲜重存在显著差异(df=3,F=7.464,P<0.05),各种源茎鲜重表现为ZL>TB>DA>JC,其中ZL和TB种源显著高于其他两个,但DA与JC种源之间无显著差异(图3)。4个种源地羊草叶干重存在显著差异(df=3,F=3.35,P<0.05),各种源叶干重表现为:TB>ZL>DA>JC,其中TB种源显著高于JC种源(图3)。4个种源地羊草鞘干重存在显著差异(df=3,F=5.787,P<0.05),各种源鞘干重表现为:ZL>TB>DA>JC,其中ZL和TB种源显著高于JC种源(图3)。4个种源地羊草茎干重存在显著差异(df=3,F=5.419,P<0.05),各种源茎干重表现为:ZL>TB>DA>JC,其中ZL和TB种源显著高于JC种源(图3)。
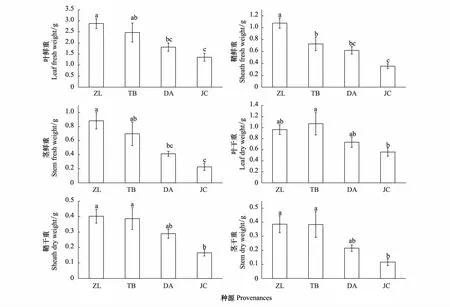
图3 不同种源间叶、鞘和茎的生物量的比较 Fig.3 The comparison of biomass of leaves, sheaths and stems between different provenances
2.3 不同种源间羊草光合特性、表型特征比较
不同种源地羊草叶片净光合速率不存在显著差异(df=3,F=22.11,P<0.05),而蒸腾速率(df=3,F=4.43,P<0.05)和水分利用效率(df=3,F=20.72,P<0.05)存在显著差异(图4)。4个种源地Pn大小表现为:JC>ZL>DA>TB(图4)。各种源间Tr大小表现为:JC>TB>ZL>DA,ZL与JC种源之间存在显著差异,DA与JC种源之间存在显著差异(图4)。各种源间WUE表现为:ZL>DA>TB>JC。不同种源地羊草生物量之间存在显著差异(df=3,F=3.87,P<0.05),ZL种源地为最高,且显著高于JC种源地,但与TB和DA不存在显著差异,各种源羊草生物量表现为:ZL>DA>TB>JC(图4)。不同种源地羊草比叶面积之间存在显著差异(df=3,F=5.75,P<0.05),羊草比叶面积在不同种源间表现为:ZL>JC>TB>DA,DA种源与其他三个种源均存在显著差异,并且DA种源呈现出了最低的比叶面积的特征(图4)。不同种源地羊草分蘖数之间不存在显著差异(df=3,F=1.06,P=0.39),各种源羊草分蘖数表现为:DA>ZL>JC>TB(图4)。
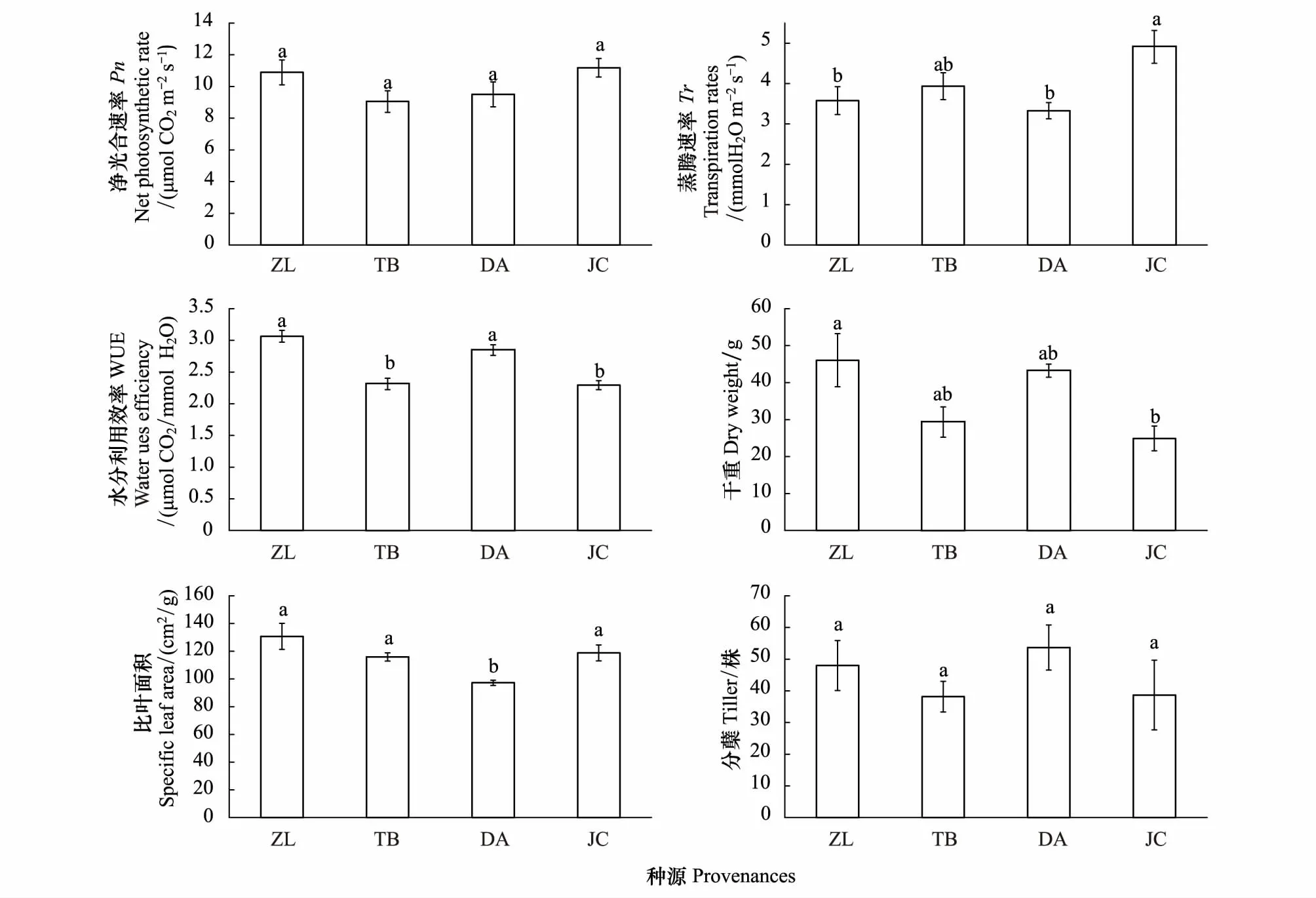
图4 不同种源间羊草光合特性、表型特征的比较 Fig.4 The comparison of photosynthetic characteristics and phenotypic characteristics between different provenancesPn表示为净光合速率,net photosynthetic rate;WUE表示为水分利用效率,water ues efficiency; Tr表示为蒸腾速率,transpiration rates
3 讨论
3.1 不同种源羊草表型特征的差异
表型可塑性被认为是物种发生进化和适应的基本条件,在物种适应生境中发挥重要作用,并逐渐成为植物适应环境变化产生表型差异的机制[23-25]。本研究中,羊草表型差异明显也证明了羊草在不同生境中有着较高的表型可塑性。在亲本经历过环境的干扰后,亲本生长的环境能够强烈的影响后代生长特性的表达[26]。在本研究中,不同种源羊草叶片表型特征存在显著差异。表明羊草的表型差异性是在长期的环境变化中由遗传因素造成的。这样的结果在其他类似的研究中也有发现,在受到长期的过度放牧或刈割处理诱导的记忆下,羊草后代叶片表型大小会出现下降的趋势[27-28]。Groot等的研究则表明,后代表型差异变化是依赖于亲本生存环境的[29]。表型差异的形成在多年生植物物种中被认为是重要的生长策略,因为相比环境条件的快速变化,自然选择产生的基因变化是很慢的[30]。各种源分蘖数高低特征与水分利用效率和生物量相似,表现为:DA>ZL>JC>TB(图4)。本研究中,ZL种源羊草长势最好,且在其他特征中都表现出最高值。虽然各种源羊草分蘖数相差较多,但各种源间并不存在显著差异,这可能是各种源羊草分蘖可塑性较低。
植物的叶片性状与植物利用资源的能力密切相关,并能反映植物为了适应多变的环境而形成相应的生长对策[31]。作为反映植物利用资源重要的指标之一,比叶面积(SLA)代表植物体单位质量的干物质所获得的捕光面积,它常常与植物的生长对策有紧密的联系,能够反映植物适应环境的特征。比叶面积也是评价群落生产力与结构是否合理的依据[32]。生长在丰富资源环境条件下的物种常常具有很高的比叶面积[32-34]。有研究认为,SLA较高的植物往往具有较高的净光合速率、生长速率,和养分利用效率。而低SLA的物种能够对资源匮乏的环境产生积极的适应力[35]。在本研究中,ZL种源羊草的SLA最高,DA种源羊草SLA显著低于其他三个种源,表明DA种源羊草生长资源环境较为差,这可能因为植物通过比叶面积和叶片组织密度共同来维持体内的水平平衡,减少比叶面积,叶干质量减小,叶组织密度增大造成的[36]。这与宋沼鹏等人对五角枫(Acermono)的研究结果相似[37]。也有研究认为,SLA的降低代表着叶片将变得更厚,蛋白都集中在叶中并参加到光合作用中以减少了水分亏缺对光合作用的负面作用[38]。羊草SLA低也可以被认为植物为了获取环境中最大的碳收入和最少的水分消耗而采取的一种良性的生存对策。本研究将SLA与叶片干物质含量一块研究发现,DA种源有最低的SLA值,但有着较高的叶片干物质含量。Rose等认为,具有较低SLA而叶片干物质含量较高的植物有着更高的水分利用效率和资源获取能力,借此适应贫瘠的环境条件[39]。在环境影响物种后代繁殖条件下,良好的条件使亲本产生大量中等质量的后代,而在贫瘠的生境中,亲本则产生数量较少但适应力很高的后代[9]。这被认为是植物适应异质生境的一种权衡对策。
生物量分配是植物生活史理论中的核心概念,也是植物生长状态和适应生境能力的重要指标[40]。植物受到生物或非生物压力下,会权衡的将获取的资源分配给不同部位[41-42],借此提高植物的适合度,增强它们的生存能力与竞争能力。植物种群的生殖分配是植物与环境相互作用、长期进化性的结果,不仅受到自身遗传学因素影响,也受到环境因素的影响,从而表现出明显的地域差异。在本研究中,我们发现在不同的种源中,叶、鞘和茎都存在显著差异,而且叶的生物量占叶鞘茎总体生物量的一半以上。Marchiori等的研究认为,高生物量是由强的光合作用引起的,在达到光饱和点之前,光合作用的增强能够积累更高的生物量[38]。这也符合本实验中不同种源羊草净光合速率的差异。相比良好条件的环境,干旱的生境使植物的后代有着更发达的根系系统,这使得经历过干旱胁迫的后代有着比生长在良好条件下的后代有更高的生物量[15]。Hamann等的最新研究认为,地上生物量是随着干旱的增加而降低,这也是通过植物将更多生物量分配到根系中来解释的[43]。在本研究中,ZL和DA种源的生物量较高也有可能是因为亲代生长在干旱的条件下,通过遗传效应使得后代具有更发达的根系系统从而产生更高的生物量。根据生物量最优分配理论,当植物生长遇到养分和水分限制时,植物会将生物量更多的分配到根部。而当植物受到光照和温度限制时,某些植物则将更多地生物量分配的地上部分[44]。但由于本实验没有对地下生物量进行测定,故无法推断羊草整体生物量分配情况,这需要后续的研究。
3.2 不同种源的生理特性差异
光合作用在植物生活史中复杂的生理过程,受到多种因素综合作用的影响,如叶绿素含量,纬度,CO2浓度等[45-46]。净光合速率(Pn)是植物生长和产量形成的关键,Pn的决定了植物光合作用的大小,并制约着植物的生长量和生长速率。Martin等认为,Pn和种源原地的气候条件相关联,这是植物在长期的环境适应中形成的结果[47]。也有研究认为,气孔因素和叶肉细胞的羧化活动也能影响净光合速率[48]。然而在本研究中,各种源羊草Pn没有显著差异。Ren等的研究发现,长期过度放牧条件通过降低叶片叶绿素含量和Rubisco酶活性来降低羊草后代的光合作用,并且调控了一系列光合速率、气孔导度和叶绿体的相关基因[28]。因此,不同种源间Pn不存在显著性差异有可能是因为在长期的环境适应中,不同种源的羊草叶片中的叶绿素含量或气孔因素等差异不大引起的。
植物叶片水分利用效率(WUE)为测定植物碳固定效率、耗水量和评估植物在胁迫条件下的适应性提供了重要指标[48]。全先奎等人认为,WUE和种源原地的年平均气温、年平均降水量呈负相关,和年平均干燥度呈正相关[49]。Warren等认为,不同种源桉树(Eucalyptussideroxylon)的叶片WUE存在差异,但与降水无明显的关系[50]。在干旱的条件下,羊草有更高的WUE,使其能在广泛的生境中生存[49]。而作为影响WUE变化的两大关键因子净光合速率(Pn)和蒸腾速率(Tr),两者的变化都会直接影响WUE。在本研究中发现,各种源间羊草Tr存在显著性差异,Pn不存在显著性差异,这说明各种源环境较为干旱,空气中的水分和土壤含水量较低,对于Tr的影响比Pn大造成的[51]。本研究中,JC种源虽然净光合速率和蒸腾速率表现出最高值,但是由于蒸腾速率过高,反而降低了WUE,积累的干物质量也因为呼吸作用被消耗掉。通过对比各种源WUE大小可发现:ZL>DA>TB>JC,而不同种源生物量(干重)也与此一致。这可能是因为较高的WUE会加速生物量的积累造成的。
4 主要结论
本研究将4个不同种源地的羊草,经过同质园实验培养后,不同种源的羊草形态、生长和生理中特征存在显著性差异。本研究不仅发现在同质园羊草移栽情况下表型差异显著,还发现羊草的表型有很广的可塑性范围。并且认为这种可传递到子代的表型差异是由遗传因素主导的。这为羊草在未来全球气候变化中的适应力提供了新的观点。除此之外,本研究在未来将选取更多环境因素的参数,为羊草表型差异提供更强有力的证据。而羊草的表型差异在消除不同环境影响后能否稳定保持,有待深入研究。
