NAD(P)H依赖型氧化还原酶不对称还原胺化制备手性胺的研究进展
2020-04-15程峰李清华李恒薛亚平
程峰,李清华,李恒,薛亚平
1 浙江工业大学 生物有机合成技术研究浙江省重点实验室,浙江 杭州 310014
2 浙江工业大学 手性生物制造国家地方联合工程研究中心,浙江 杭州 310014
手性胺是指手性中心含有氨基的一类化合物。其作为一种结构单元,在生物活性小分子中占有相当大的比例,并构成了众多医药及农药关键中间体[1-2]。研究表明FDA批准的小分子药物化合物结构中大约40%都含有1个及以上的手性胺模块[3]。鉴于光学纯手性胺在不同领域的重要应用,由简单易得的原料出发,高效、绿色合成手性胺成为当前重要的研究领域。
目前手性胺的合成方法主要包括化学催化合成法、生物催化合成法和组合催化合成法[4-5]。其中化学催化潜手性C=N键的不对称还原反应已经研究得较为透彻。但是随着绿色工业制造对三废排放等的要求不断提高,传统化学合成的方法局限性日益突出;而生物催化的不对称合成因具有反应条件温和、选择性高、绿色无污染等优点,逐渐被广泛应用于手性胺的合成中[6-8]。并且随着分子生物学及合成生物技术快速发展,对酶的改造和修饰变得更加容易[6]。生物催化的不对称合成已有的报道包括NAD(P)H依赖型氧化还原酶还原胺化生产手性胺 (图1)、水解酶不对称水解外消旋胺、黄素依赖性单胺氧化酶 (MAO)[9]去消旋化合成手性胺、ω-转氨酶以酮化合物作为底物不对称合成手性胺等。尽管ω-转氨酶合成手性胺有一些成功的实例,比如Codexis公司开发的ATA-117突变体成功为糖尿病治疗药物西他列汀(含有1个手性胺模块) 的化学-酶法合成提供了高效的催化剂,但是,转氨酶催化合成手性胺反应是一个可逆反应,受到平衡常数的影响,底物往往很难完全转化[10]。同时转氨酶的催化机制决定了它的催化范围仅允许-NH2的转移:在转氨过程中,来自供体的氨与辅因子5′-磷酸吡哆醛(PLP) 发生反应,将氨保留在辅因子5′-磷酸吡哆胺 (PMP) 中;之后将氨转移到反应的共底物中;最终,潜手性酮转化为手性伯胺 (初级胺)。而对于手性仲胺则可以使用MAO拆分外消旋胺底物,但是理论最大得率仅为50%。亚胺还原酶 (IRED)也可用于光学纯手性仲胺的制备,但是由于亚胺底物本身合成的困难,限制了该酶的应用范围[11]。还原性酮胺化酶 (RedAm) 可以以等摩尔的酮和胺为底物,催化形成碳酰胺中间体,从中除去水形成亚胺离子;然后,亚胺离子在烟酰胺辅因子NAD(P)H的作用下不对称还原,生成产物手性胺。此外,还原性酮胺化酶还需要一种机制来防止底物酮还原为醇。
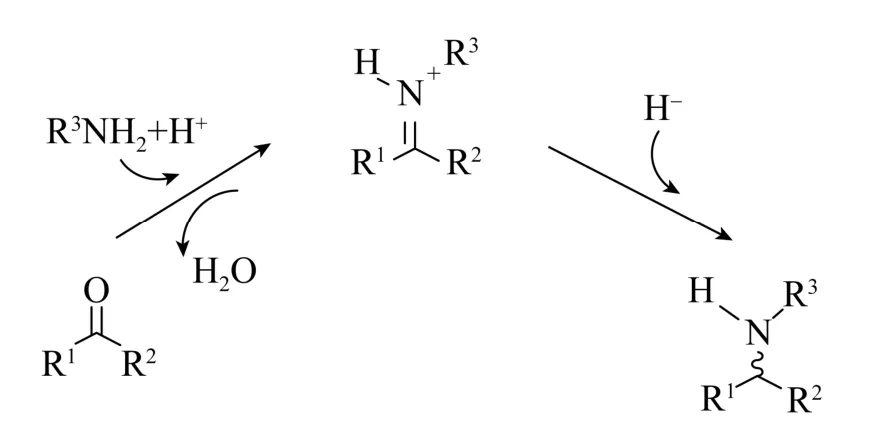
图1 还原胺化反应一般过程Fig.1 Scheme of reductive amination reaction.
以NAD(P)H依赖型氧化还原酶为催化剂、酮化合物为底物,一步还原胺化合成手性胺的反应理论最大收率 (Yield) 和产物光学纯度 (e.e.) 均可达到100%。因此,该反应是合成手性胺的最重要反应之一。本文中,我们以亚胺还原酶、氨基酸脱氢酶、冠瘿碱脱氢酶和还原性酮胺化酶为例,从酶的结构和机理、分子改造及催化应用等方面总结了NAD(P)H依赖型氧化还原酶在不对称还原胺化合成手性胺中的研究进展。
1 NAD(P)H依赖型氧化还原酶的介绍
1.1 亚胺还原酶
2011年,Mitesukura等首次报道了来源于链霉菌Streptomyces的亚胺还原酶 (Imine reductase,IRED,EC 1.5.1.48) 能够不对称还原环亚胺形成手性胺[12-16]。两个来自Streptomyces的NADPH依赖型亚胺还原酶能够催化亚胺2-甲基-1-吡咯啉不对称还原胺化 (图2),产物具有互补的立体构型,区别于酶催化酮还原胺化的一般性机制(图3)。在过去5年中,越来越多的IRED被发现并应用于一系列潜手性化合物的不对称还原制备手性胺,例如苯甲醛还原为N-苯甲胺,4-苯基丁-2-酮还原为4-苯基丁-2-胺等。

图2 通过来自Streptomyces sp.GF3546和GF5387的(S)-和(R)-IRED对2-甲基-1-吡咯啉进行同步互补还原[15-16]Fig.2 Enantio-complementary reductions of 2-methyl-1-pyrroline by (S)- and (R)-IREDs from Streptomyces sp.GF3546 and GF5387[15-16].

图3 推定的酶催化酮还原胺化的一般性机制[1-3]Fig.3 Putative mechanism for enzyme-catalyzed reductive amination of ketone[1-3].
1.2 氨基酸脱氢酶
氨基酸脱氢酶 (Amino acid dehydrogenase,AADH,EC 1.4.1.X) 是一种NADH或NADPH依赖的氧化还原酶,催化酮化合物和氨基酸之间的相互转化 (图4)[17]。在胺化方向上,通过氨基酸脱氢酶催化酮酸和氨基供体偶联形成亚胺中间体,然后利用辅助因子提供的氢化物还原亚胺中间体。关于氨基酸脱氢酶的机理及应用已经有许多报道[18],其中研究最广泛的氨基酸脱氢酶是亮氨酸脱氢酶 (LeuDH)[19]、苯丙氨酸脱氢酶(PheDH)[20]、谷氨酸脱氢酶 (GluDH) 以及二氨基庚二酸脱氢酶 (DAPDH)[21]。此外还有关于甘氨酸脱氢酶、丙氨酸脱氢酶、天冬氨酸脱氢酶、赖氨酸脱氢酶和色氨酸脱氢酶的报道。

图4 氨基酸脱氢酶催化的还原胺化反应Fig.4 Reductive amination reaction catalyzed by amino acid dehydrogenase.
1.3 冠瘿碱脱氢酶
冠瘿碱脱氢酶 (Opine dehydrogenase,OpDH,EC 1.5.1.28)能可逆地催化氨基酸和酮酸形成N-衍生氨基酸 (冠瘿碱)[22]。OpDH主要存在于细菌和高等生物体中,细菌来源的这类酶主要是NADH依赖型,例如来自节杆菌Arthrobactersp.的冠瘿碱脱氢酶可以催化氨基戊酸和丙酮酸生成2S-2-[(1-R-羧基)-氨基]戊酸盐 (图5);此外在细菌中如农杆菌Agrobacterium也发现一些黄素依赖型的OpDHs[23]。在高等生物体中 (如软体动物和海绵生物) OpDH主要是NADPH依赖型的,其主要生理作用是在缺氧条件下维持糖酵解通量[24]。OpDHs除了可以催化底物与无机铵偶联反应以外,还可以催化含有羧基的化合物与有机胺偶联的反应,通过对其进行改造还可以获得催化合成仲胺的突变体。

图5 冠瘿碱脱氢酶OpDH催化的还原胺化反应方程[25]Fig.5 Reductive amination reactions catalyzed by opine dehydrogenases OpDH[25].
1.4 还原性酮胺化酶
利用还原性的酮胺化反应,以酮与胺为底物等摩尔制备手性胺是许多生物催化专家的研究目标[26]。2017年,英国曼彻斯特大学Turner教授研究组在Nature Chemistry上发表了一个来源于米曲霉Aspergillus oryzae的还原性酮胺化酶 (Reductive aminase,RedAm,EC 1.5.1.28) 的酶学性质系统研究及结构解析[27]。在这篇文章中,作者首次将这种酶命名为“reductive aminase”,该酶能够直接催化羰基化合物和胺类物质耦合成相对应的亚胺,并实现亚胺的不对称还原。文中选取了32个酮(分成4组) 和18个胺 (分成3类) 构成的底物化合物库被用于对其底物特异性的研究。在胺供体与辅酶NADPH充足的条件下,该酶对环状酮、C5-C6线性酮及醛的活力明显高于C4 酮化合物,具有较好的对映异构选择性;而对亲核性胺化合物的筛选中发现该酶显示出对伯胺的偏好性,特别是不饱和的脂肪伯胺。进一步针对不同的酮化合物,选择合适的醇脱氢酶偶联酮胺化酶,可以实现生物催化“借氢”反应路径,并对底物还原胺化[28]。尽管这些研究报道鼓舞人心,但目前对该类酶的序列、酶学性质研究与其他手性胺制备常用酶相比还相对偏少。
2 NAD(P)H依赖型氧化还原酶的结构与机理
2.1 亚胺还原酶的结构与催化机理
IRED具有二聚体结构,其中一个亚基分子量大约30 kDa,通过结构域的共享来形成活性口袋,其活性位点位于其中一个亚基Rossman折叠的N端和临近亚基C-端的交接处 (图6A)[29-30]。每个亚基的两个结构域之间由一个长α螺旋来连接。整个二聚体在协同催化过程中是相对柔性的,与底物结合时可以观察到结构域之间明显的闭合。IREDs的晶体结构与羟基异丁酸脱氢酶(HIBDHs) 的晶体结构高度相似。但是在HIBDHs结构中 (例如2CVZ,来源于极端嗜热菌Thermus thermophilus的HIBDH) 并没有观察到类似IREDs的结构域共享机制[31]。在HIBDH结构中,活性位点Lys残基的作用是在还原方向上使新生醇质子化。当把HIBDH和IRED的结构叠加时,可以看到2CVZ结构的Lys残基与IRED结构中不同的残基重叠 (比如Lys与Asp或Tyr重叠)。有观点提出,这些残基在IRED催化的反应中起到对胺产物提供质子的作用[29]。尽管这些氨基酸残基在催化机制方面的作用还没有通过结构研究得以证实,但是将这些残基突变为丙氨酸会导致其催化活性大幅下降。最近,来自东方拟无枝酸菌Amycolatopsis orientalis的IRED结构被解析出来,这是第1个有辅因子存在的情况下观察到的IRED与胺产物 (R)-1-甲基-四氢异喹啉 (R-MTQ)结合的复合物结构 (图6B)[32]。与没有配体结合的IRED相比,复合物结构中催化口袋变得更加封闭,并且胺产物与活性位点Y179和N241残基侧链形成相互作用。这证实了活性位点氨基酸残基在配体铆定方面发挥着重要作用,同时它使胺的亲电子碳与NADPH的烟酰胺环C-4之间处于一个合适的距离,有利于氢化物的传递。IRED活性位点中的Asn与HIBDH活性位点的Lys作用是一样的,即在还原方向上给中间产物提供质子,因此在IRED催化亚胺还原过程中发挥着重要作用。

图6 亚胺还原酶的结构示意图 (A:来自Amycolatopsis orientalis的亚胺还原酶 (AoIRED) 结构图;B:AoIRED的活性位点与外消旋MTQ和NADPH的复合物结构图,改编自[32])Fig.6 The structures of imine reductase.(A) The structure of imine reductase from Amycolatopsis orientalis (AoIRED).(B) The active site of AoIRED with racemic MTQ and NADPH (adapt from [32]).
2.2 氨基酸脱氢酶的结构与催化机理
目前,针对氨基酸脱氢酶的还原胺化反应,研究较多的是亮氨酸脱氢酶 (LeuDH) 和苯丙氨酸脱氢酶 (PheDH) 等。Rice等确定了一个来自球形芽孢杆菌B.sphaericus的LeuDH晶体结构(PDB︰1LEH)[33]。该酶的晶体结构中,两个亚基缔合形成二聚体,其中一个亚基由364个氨基酸组成,存在由深裂缝分开的两个结构域。该深裂缝是辅因子结合位点及底物结合位点 (图7A)。而在溶液中,LeuDH的四级结构普遍被认为是一个八聚体,结构域运动在催化作用中发挥重要作用,在底物存在的情况下关闭结构域,从而使辅因子和底物足够接近以进行氢化物转移。
酶-底物模型表明,该酶通过115位天冬氨酸残基及68位、80位两个赖氨酸残基与底物结合,将L-亮氨酸锚定在活性中心 (图7B)。K80在AADHs序列中是高度保守的。如果将K80突变成Ala或Gln,K80Q突变体Km值从野生型的5.1 mmol/L增加至17 mmol/L,K80A突变体Km值则降低至3.7 mmol/L (以L-亮氨酸为底物)。Sekimoto等也分析了K80的作用,其最主要的作用被证明是参与了水的活化,并在脱氨方向上对亚胺离子的亲电碳进行攻击,具体催化机制如图8所示[34]。通过NAD+从亮氨酸 (Ⅰ) 中提取氢化物形成亚胺离子 (Ⅱ),K80作为质子供体,激活水分子以攻击亲电碳原子,形成碳酰胺中间体 (Ⅲ),从中间羟基到胺分子内质子转移形成氧负离子(Ⅳ),通过K80的侧链稳定氧负离子 (Ⅳ),最终脱去氨基基团,生成产物α-酮异己酸盐 (Ⅴ)。目前已解析的PheDH的晶体结构中包括能催化苯丙酮酸和L-苯丙氨酸之间相互转化的1BW9和1C1D,以及能催化苯基乳酸和β-苯丙酸酯之间相互转化的1C1X与1BXG。有研究指出,PheDH的催化活性是通过活性位点上的结构域闭合来实现的,使底物和辅因子充分接近以进行催化反应。例如来源于粟褐芽孢杆菌Bacillus badius的苯丙氨酸脱氢酶[35]和来源于红球菌Rhodococcussp.M4的PheDH[36-37]。它们具有48%的序列同源性,晶体结构也非常相似,同样由两个结构域构成,中间由一个大裂隙分开,辅因子结合在其中,结构域的移动被证明参与了还原胺化和氧化脱氨反应。

图7 亮氨酸脱氢酶的结构示意图[33] (A:含有NADH和L-亮氨酸的LeuDH复合物结构;B:含有L-亮氨酸及NADH的活性位点结构图)Fig.7 The structure of leucine dehydrogenase[33].(A) The complex structure of LeuDH with NADH and L-Leucine.(B)The active site of LeuDH with L-leucine and NADH.
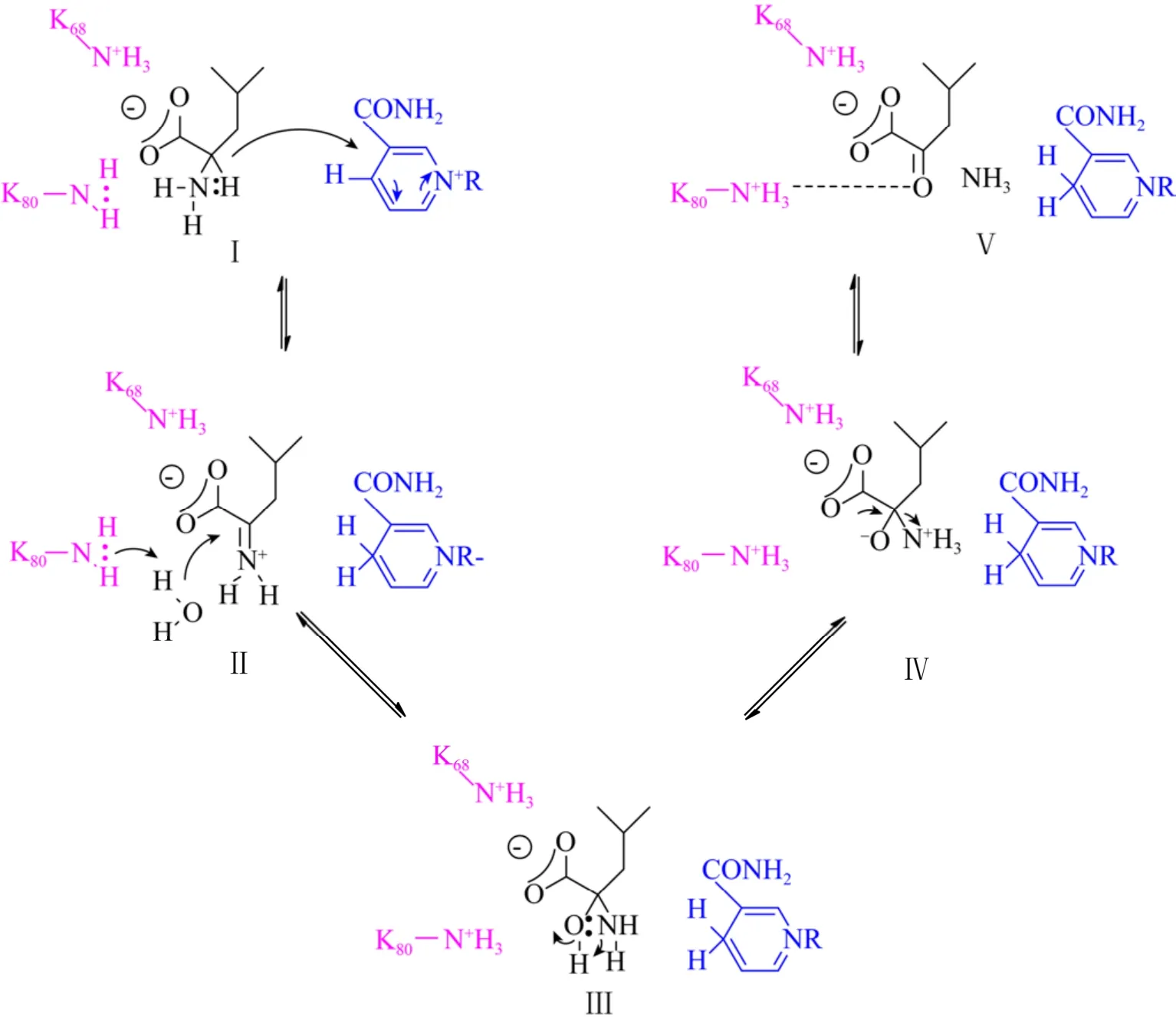
图8 LeuDH对L-亮氨酸脱氨基的催化机制[34] (Ⅰ:L-亮氨酸;Ⅱ:亚胺离子;Ⅲ:碳酰胺;Ⅳ:含氧负离子;Ⅴ:α-酮异己酸盐)Fig.8 Proposed mechanism of deamination of L-leucine by LeuDH[34].I:L-leucine;II:iminium ion;III:carbinolamine;IV:oxyanion;V:α-ketoisocaproate.
来自Rhodococcussp.的PheDH与L-Phe的复合物图如图9A所示,类似于LeuDH催化机制,底物氨基与118位点的天冬氨酸相互作用,羧基与66位点和78位点赖氨酸残基以及262位点天冬酰胺的侧链相互作用 (图9B)。当以3-苯基丙酸酯作为底物对野生型及突变体进行动力学测定时发现对于底物结合来说,D118与底物氨基的相互作用并不特别重要,而K68侧链在底物存在时,发挥了识别水分子的作用,水分子参与形成碳酰胺中间体。关于脱氨方向的催化机制被认为是首先与辅因子NAD+结合,然后通过L-Phe有序释放铵离子、苯丙酮酸和NADH。
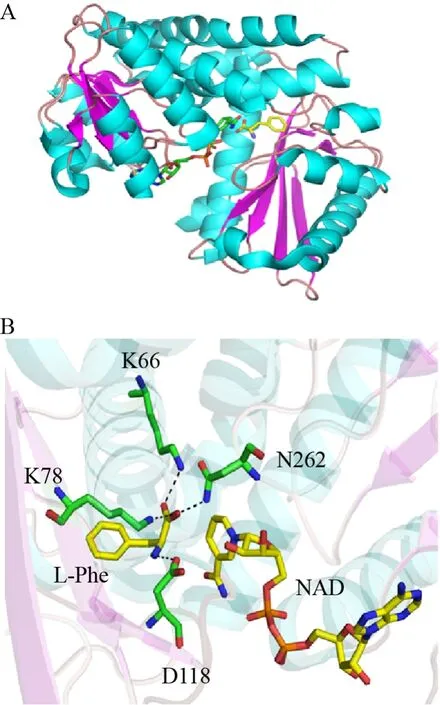
图9 苯丙氨酸脱氢酶结构示意图[37] (A:含有NAD和L-Phe的PheDH (1C1D) 单体结构图;B:PheDH活性位点处底物与辅酶及关键氨基酸残基的结构图)Fig.9 Structures of phenylalanine dehydrogenase[37].(A) Structure of monomer of PheDH (1C1D),with NAD and L-Phe.(B) The PheDH active site structure of substrate,NAD and key residues.
基于此,Brunhuber等推测出了类似于LeuDH脱氨机制的PheDH脱氨基催化机制 (图10)[37]。底物以两性离子形式存在,胺处于质子化状态。K78活化水分子催化氨基 (Ⅰ) 去质子化,氢化物转移至NAD+与其相互作用 (Ⅱ),形成亚胺离子 (Ⅲ);接着K78活化水分子以攻击亚胺碳(Ⅲ),得到碳酰胺中间体 (Ⅳ);然后K78催化碳酰胺去质子化 (Ⅴ),D118向胺基团提供质子,使得产物苯丙酮酸的羰基与K78的侧链结合 (Ⅵ),可以防止苯丙酮酸被辅因子还原成苯基乳酸。底物为苯基乳酸时,酮基与K78的侧链结合使得羰基碳与NAD(H) 的C-4的距离变为5.1 Å,对于它们之间发生氢化物递送而言距离远,因此反应很难进行。
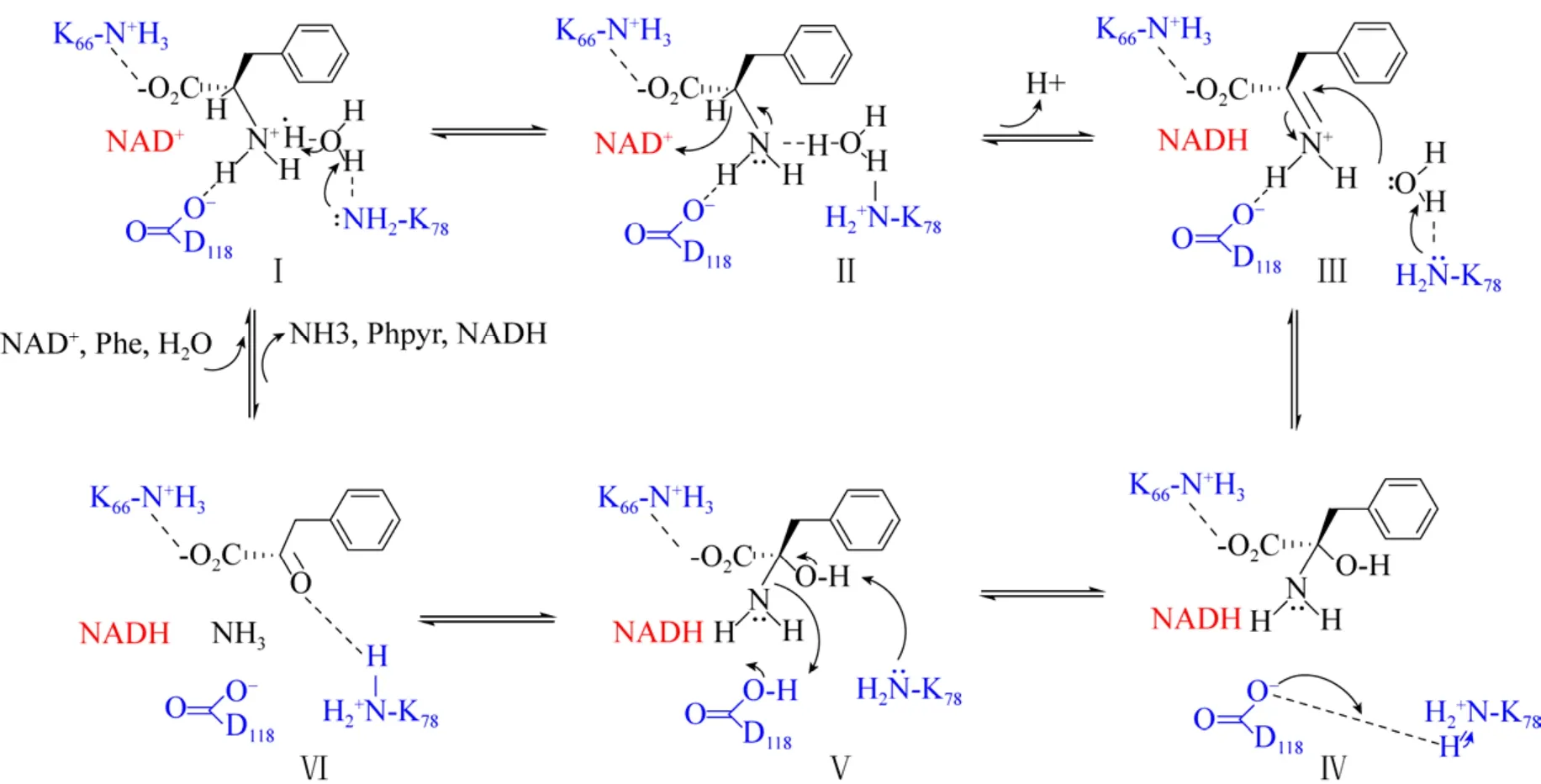
图10 推定的PheDH催化机制 (改编自[37])Fig.10 Mechanism of PheDH adapted from Brunhuber and co-workers[37].
在D-氨基酸脱氢酶 (DADH) 中研究最为广泛的是一种NADP+依赖性的氧化还原酶内消旋-2,6-二氨基庚二酸脱氢酶 (Meso-DAPDH)。meso-DAPDH可以分为两大类型:一类是表现出对内消旋-二氨基庚二酸 (Meso-DAP) 底物专一的氧化脱氨活性,属于Ⅰ型,例如来源于谷氨酸棒杆菌Corynebacterium glutamicum的meso-DAPDH (CgDAPDH);另一类不仅表现出对内消旋-二氨基庚二酸 (Meso-DAP) 底物氧化脱氨活性,更表现出优越的可逆胺化活性,并且具有更宽的底物谱,属于Ⅱ型,例如来源于嗜热菌Symbiobacterium thermophilum的meso-DAPDH(StDAPDH) (图11)[38-40]。

图11 内消旋-2,6-二氨基庚二酸脱氢酶催化的可逆反应[38-40]Fig.11 The reversible reaction catalyzed by DAPDH[38-40].
来源于Symbiobacterium thermophilum的meso-DAPDH (StDAPDH) 是在自然界中发现的第一个具有还原胺化活性的D-氨基酸脱氢酶,通过对StDAPDH的晶体结构 (PDB︰3WBF) 研究发现,StDAPDH属于六聚体结构,辅因子和底物结合在其中 (图12A)。StDAPDH通过与CgDAPDH序列比对及保守型分析,发现StDAPDH的两个氨基酸残基R35和R71在Ⅱ型中是高度保守的,通过进一步的实验研究发现,R35只是活性相关位点,并不是底物偏好性的相关氨基酸残基。而R71不仅是活性相关位点,而且是影响StDAPDH底物偏好性的重要氨基酸残基。此外StDAPDH中还存在两个丙酮酸的底物结合通道,位于M152的两侧,Asp92和Asn253与M152相互作用,从而可能影响StDAPDH结构域运动及其催化性能 (图12B)[39]。
通过进一步的研究,Gao等推导出StDAPDH催化内消旋-二氨基庚二酸脱氨基催化机制(图13)[41],首先是氢化物从内消旋-二氨基庚二酸Cα转移至辅因子烟酰胺环的C4N上,生成亚氨基酸 (Ⅰ);接着在H154的作用下,水分子进攻亚氨基酸,形成甲醇胺 (Ⅱ);随后该中间产物分解生成L-2-氨基-6-酮基庚二酸和氨 (Ⅲ)。
2.3 冠瘿碱脱氢酶的结构与催化机理
Asano等在以N-(1-(R,S)-羧基乙烷基)-S-苯丙氨酸 (图14) 为底物的富集培养基上筛选到含有OpDH的节杆菌Arthrobactersp.strain 1C,并从该菌株中成功纯化出OpDH[25]。以L-苯丙氨酸、丙酮酸盐为底物,NADH作为辅酶,OpDH-CENDH作为催化剂催化底物形成N-(1-(R,S)-羧基乙烷基)-S-苯丙氨酸。其他疏水性L-氨基酸包括L-甲硫氨酸,以及D-氨基酸包括D-亮氨酸也可以作为该酶的底物,但是对L-甲硫氨酸的催化活性很低,转化率仅有3.4%。同时,草酰乙酸以及碳链更短的乙醛酸也可以作为底物。
Asano教授课题组通过进一步的研究,成功克隆出该酶基因,编码359个氨基酸[42]。基于此,该酶的裸蛋白结构[43]及与底物精氨酸和苯丙酮酸的复合物结构[44]都得以解析。与其他AADHs相同,OpDH在两个结构域的交界处有一条裂隙,并且可以在裂隙中观察到辅酶 (图15A)。借助NMR、ITC等手段,结合复合物结构基本阐明了OpDH的催化机制[45]:在以L-精氨酸为底物的晶体复合物中,L-精氨酸的α-羧基与H212相互作用,胍基与E142的侧链基团相互作用 (图15B)[44]。由于这些发挥作用的氨基酸分别来自于不同的结构域,因此可以认为是L-精氨酸的结合触发了不同结构域的闭合,实现了酶的偶联催化。在以丙酮酸为底物的晶体复合物中,丙酮酸的α-羧基与Q118的侧链相互作用,H212与底物羰基形成氢键。由于羰基位置远离辅酶NADH,因此无法观察到从丙酮酸还原到乳酸的过程。同时将118位点进行突变,获得突变体Q118A,其还原活性大幅度下降。基于这些结果,可以得出精氨酸与丙酮酸这两个底物与酶的结合顺序。从精氨酸与丙酮酸叠合之后的晶体复合物中观察到Q118和H212的朝向几乎没有任何改变 (图15B)。
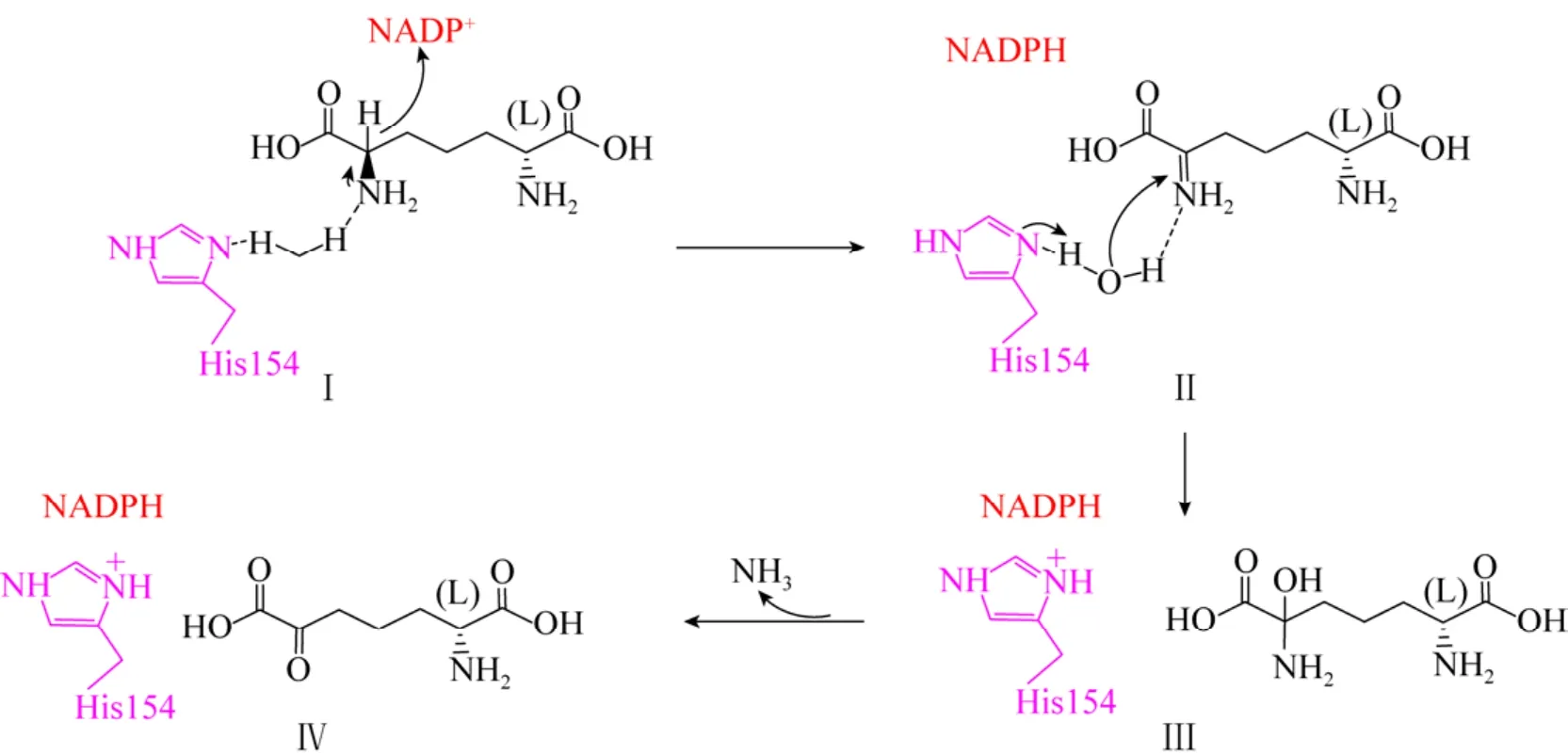
图13 StDAPDH对meso-DAP脱氨基的催化机制[41]Fig.13 Proposed mechanism of deamination of meso-DAP by StDAP[41].

图14 由来自Arthrobacter sp.strain 1C冠瘿碱脱氢酶OpDH-CENDH催化的还原胺化反应方程式[25]Fig.14 Reductive amination reactions catalyzed by opine dehydrogenases OpDH-CENDH from Arthrobacter sp.strain 1C[25].
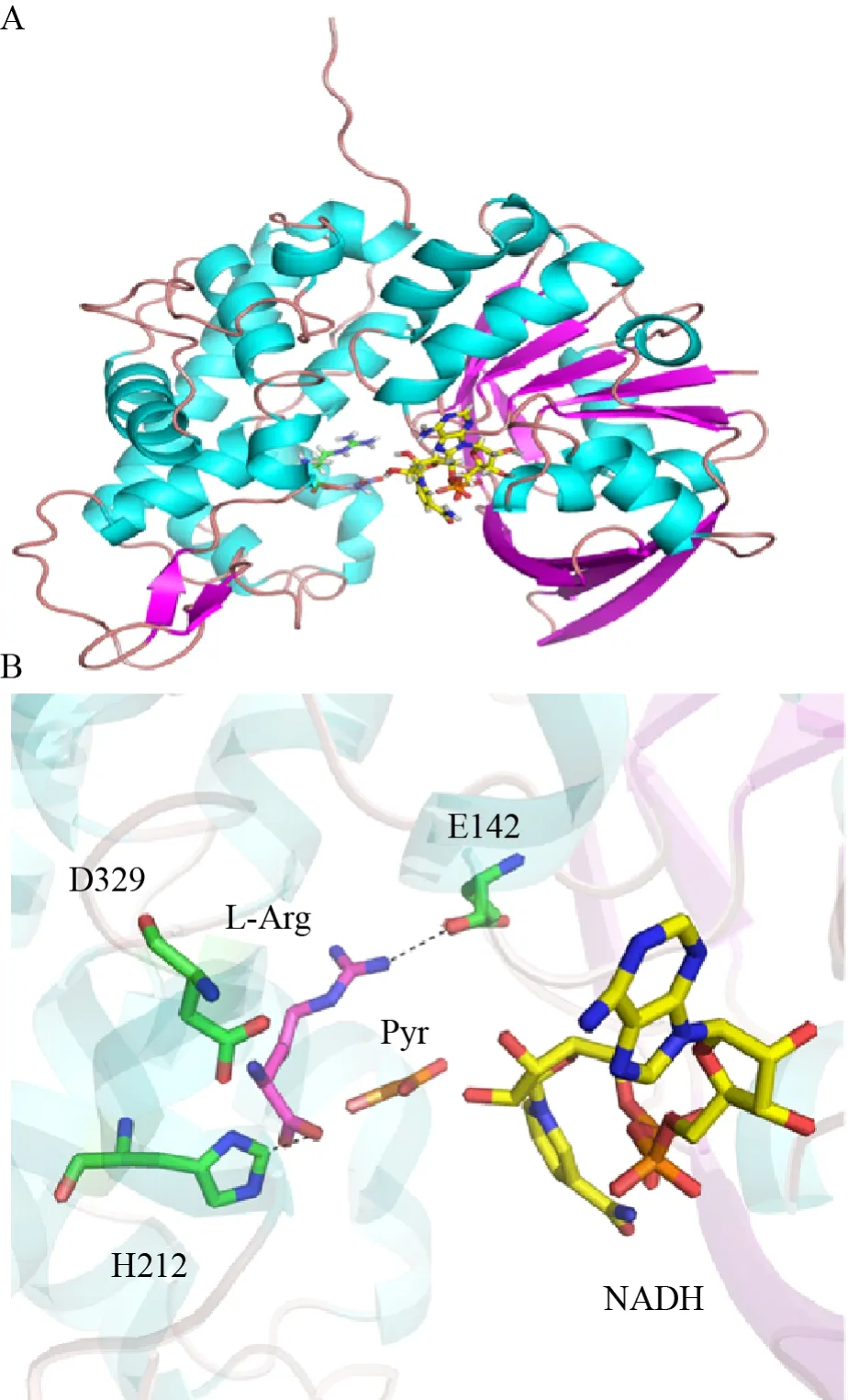
图15 OpDH的结构与机理图[43-45] (A:来源于扇贝鲆Pecten maximus的OpDH与L-精氨酸、丙酮酸和NADH的复合物结构图;B:OpDH的活性位点与底物L-精氨酸、配体丙酮酸的局部示意图)Fig.15 The structure and mechanism of OpDH[43-45].(A)Structure of OpDH from Pecten maximus with L-arginine,pyruvate and NADH.(B) The OpDH active site structure with L-arginine and ligand pyruvate.
但是,每个配体到辅酶NADPH烟酰胺环的距离都较远,这表明在活性构象中底物被烟酰胺环C4位的H还原为亚胺,因此在这种构象中包含了不同的配体结合模式。进一步,NMR的实验研究被用于探究L-精氨酸和丙酮酸与酶的结合顺序[45]。通过监测15N标记的apo蛋白,在15N-1H-TROSY光谱中选择5个峰,可以观察到NADH的结合对这些化学位移产生了扰动影响。在向OpDH-NADH复合物中添加L-精氨酸之后,观察到进一步的扰动,但是当丙酮酸添加到同样的复合物中时却没有发现扰动现象。这一点可以证明在偶联催化反应中L-精氨酸是在NADH与OpDH结合之后与酶结合的。等温量热研究也证实了这一观点:当L-精氨酸加入OpDH-NADH复合物时,可以显示出可测量的焓变,但当加入丙酮酸到同样的复合物时,却没有焓的变化。
2.4 还原性酮胺化酶的结构与催化机理
鉴于还原性酮胺化酶RedAm能够实现酮与胺的等摩尔还原胺化,科学家对其三维空间结构(apo及holo形式) 进行了解析 (PDB 5G6R:不结合底物的apo形式,PDB 5G6S:结合底物的holo形式)。结果显示该酶具有典型的亚胺还原酶同源二聚体三级结构,每个单体分别由一个N端的Rossman结构域和C端的α-螺旋束结构域构成,两个结构域中间通过一个长α-螺旋臂连接,两个结构域之间则依靠α-螺旋束之间的交叉作用力形成二聚体 (图16)。该还原性酮胺化酶的三维空间结构紧凑,底物结合区域的空间体积明显小于类似的亚胺还原酶,在酶的底物结合区域,组成了催化三连体Asn93-Asp169-Tyr177。为了探索还原性酮胺化酶底物识别分子机制与催化机制,科学家对该酶活性口袋附近的氨基酸D169、Y177进行了突变及酶学性质研究。结合动力学研究结果与晶体结构信息及结构特征,推测反应过程中亚胺中间态的形成也在酶的活性口袋,这一点明显区别于以往发现的亚胺还原酶,这也是其表现出催化活性比亚胺还原酶高的原因所在。对于还原性酮胺化酶AspRA的催化机理,认为首先是Y177侧链与酮底物的羰基相互作用,将酮底物进行固定,同时胺底物与N93和D169的侧链相互作用,D169使胺底物脱去质子,然后与酮底物偶联生成甲醇胺中间体,去除水分子形成亚胺离子,然后亚胺离子被NADPH还原,最终获得产物胺化合物 (图17)[27]。

图16 还原性酮胺化酶的二聚体结构与酶-NADPH-底物结合的放大精细图[27] (紫色区域:Rossman结构域;蓝色区域:二聚体α-螺旋束;红框区域:酶-NADPH-底物)Fig.16 The dimer structure of reductive aminase and the complex of RA-NADPH-substrate[27].Purple region:Rossman domain;blue region:dimeric α-helical beam;red box region:RA-NADPH-substrate.

图17 推测的还原性酮胺化酶反应机理[27]Fig.17 Proposed mechanism of AspRA towards ketones[27].
3 NAD(P)H依赖型氧化还原酶的改造与应用
3.1 亚胺还原酶 (IRED) 的改造与应用
首次提出IREDs具有还原胺化活性的Mueller等对来自链霉菌Streptomycessp.GF3546的S-选择性IRED不对称胺化还原4-苯基-2-丁酮生成手性胺进行了详细研究 (图18)。当甲酸铵作为氨基供体,虽然加入氨基供体的量足够多,反应过程中投入大量的酶,但是最终的转化率仍然很低,为8.8%,ee值为76%。目前尚不清楚对于这个反应IRED是否真正发生了还原胺化反应,还是因为在反应介质中加入大量氨而预先自发形成亚胺。
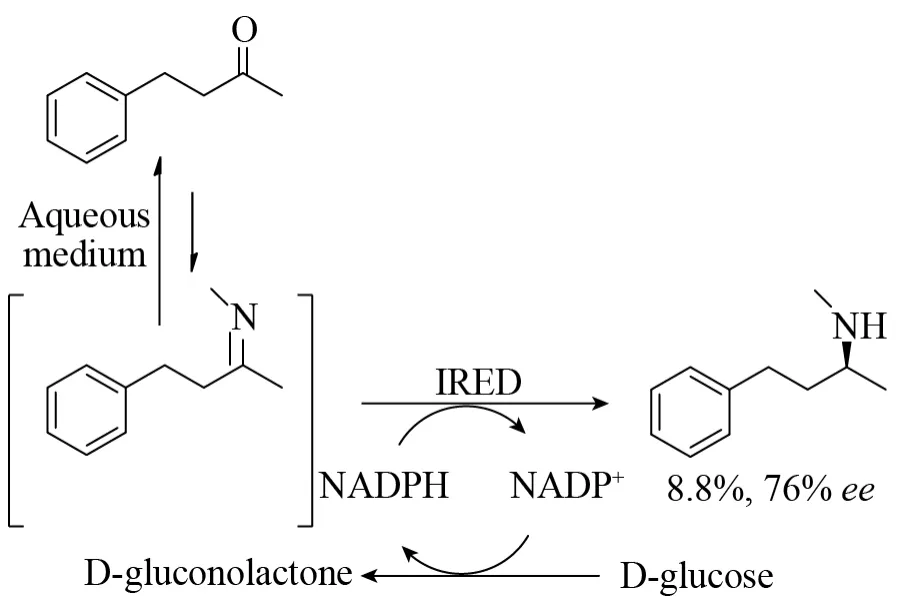
图18 由来自链霉菌属 (Streptomyces sp.GF3546)的 (S)-IRED催化还原胺化4-苯基丁-2-酮[30]Fig.18 Reductive amination of 4-phenylbutan-2-one by(S)-IRED from Streptomyces sp.GF3546[30].
为了进一步研究IREDs催化还原胺化反应的机制,Hauer等进行了双分子还原胺化反应的研究,在研究过程中将来自玫瑰链孢囊菌Streptosporangium roseum的IRED在无机铵、甲胺或者苯胺存在的情况下还原胺化苯甲醛[46]。当使用1倍的胺进行反应时,速度较慢;当胺的用量增加到10或50倍时,苯甲醛与甲胺生成N-苯甲胺,反应8 h,转化率达到73%。使用50倍的甲胺,乙酰苯和环己基酮作为底物,与甲胺反应生成相应的产物胺,转化率分别为39%和53%,ee值为87%和78%。
Roche公司的生物催化小组首次报道了具有严格立体选择性且能够不对称还原环亚胺的IREDs家族,之后该小组又继续研究了IRED催化的不对称还原胺化反应[47],向IRED酶库中添加9种酶之后,筛选对酮 (包括苯乙酮、2-己酮和环己酮) 底物和亲核胺供体 (包括氨、甲胺或丁胺) 具有还原胺化活性的IRED。实验结果表明不同底物转化率各不相同,分别在10%–90%之间,其中对苯乙酮的胺化反应产率最低。两个优势突变体“IR_11”和“IR_20”催化100 mg底物的反应中。IR_11催化酮和甲胺生成手性胺产物1,产率为71%,de值98% (图19);IR_20催化酮和无机铵生成手性胺产物2,产率为50%,de值94%。除此之外,IR_20还可以催化2-己酮和甲胺还原胺化生成手性胺产物3,产率为55%,ee值96%。上述每个反应都是在pH 9.3 (利于亚胺形成) 的条件下进行的,其中胺的摩尔量都是底物摩尔量的12.5倍。导致活性较低的原因主要归因于亚胺在水溶液中形成速率较慢,通过使用NMR也没有检测到亚胺中间体。

图19 优势突变体“IR_11”和“IR_20”催化的还原胺化反应[48]Fig.19 Reductive aminations of ketones by superior IREDs (IR_11Q and IR_20)[48].
Höhne等进一步将IRED扩展到更多的手性化合物合成上[49]。除了先前测定的活性外,再次对Roche公司的IRED酶库进行筛选。其中,使用仲胺5作为胺供体成功还原胺化环己酮4 (图20)。抗帕金森综合征药物(R)-雷沙吉兰9同样也是由Roche酶库中的IR_14催化茚酮7和炔丙胺8还原胺化生成的,产率为58%,ee值为90%。除此之外,IRED突变体“IR_Sip”用于合成(S)-雷沙吉兰,产率为81%,ee值为72%。上述的每个反应都处于pH 9.0的缓冲液中,胺供体摩尔用量为底物摩尔量的40倍,这一点证明了预形成的亚胺底物是IRED产生活性的首要条件。
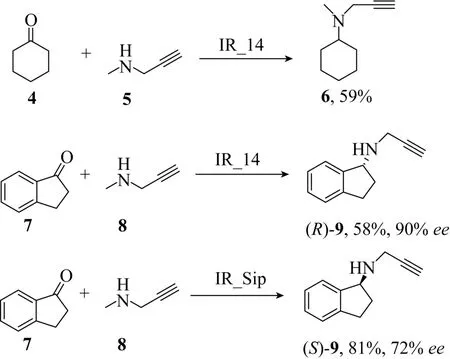
图20 利用IRED进行还原胺化反应制备(R)-和(S)-雷沙吉兰[49]Fig.20 Preparation of (R)-and (S)-rasagiline using IREDs in reductive amination reactions[49].
研究中发现,亚胺还原酶大多为NADPH依赖型,在全细胞催化的反应中NADPH的含量要都低于NADH,从而影响催化效率。Nestl等采用“Cofactor Specificity Reversal—Structural Analysis and Library Design”策略,对来自叶柄粘球菌Myxococcus stipitatus的亚胺还原酶经过多轮突变,获得的突变体V10,对NADH/NADPH的活力提高了2 900倍,从而能增加了亚胺还原酶转化2-甲基吡咯啉 (2MP) 生成2-甲基吡咯烷(2MPD)的效率[50]。
Zhu等通过对大量IREDs (包括88种酶) 进行筛选,成功筛选出亚胺还原酶优势突变体IR2和IR45 (图21)。它们能够以极高的立体选择性和转化率将间位和对位的氯/甲基/甲氧基-苄基二氢异喹啉 (DHIQ) 转化为相应的R型或S型THIQ。通过对结构的观察,发现IR45的W191位点对空间位阻影响较大,将191位的色氨酸突变为丙氨酸后,其Km值比野生型IR45降低了170倍,解除了底物抑制作用,催化效率 (kcat/Km) 比野生型IR45提高约8倍[51]。

图21 IREDs不对称还原1-苄基-DHIQ衍生物Fig.21 Asymmetric reduction of 1-benzyl-DHIQ derivatives by IREDs.
到目前为止,IRED已经被证明可以在高pH环境下对亚胺进行不对称还原,亚胺的形成一般通过如图2所示的一般催化机制来进行,但是在这个亚胺形成的过程中酶所发挥的作用尚不十分清楚。随着IRED酶序列、结构以及用于酶进化高通量方法的研究结果逐渐增加,对用于还原胺化反应的IREDs改造及机制相关研究将成为未来的研究热点。
3.2 氨基酸脱氢酶 (AADH) 的定向进化与应用
胺脱氢酶 (Amine dehydrogenase,AmDH,EC 1.4.99.3) 是一类能够催化初级胺氧化脱氨生成醛和氨的氧化还原酶。而氨基酸脱氢酶可以实现氨基酸与酮化合物之间的相互转化,因此可以通过改造氨基酸脱氢酶,获得能够催化其逆反应的胺脱氢酶,即以醛或酮为底物,无机铵为供体,通过还原胺化反应生产手性胺。例如通过定向进化改造亮氨酸脱氢酶获得的胺脱氢酶能够还原胺化α-酮异己酸酯的类似物甲基异丁基酮 (图22)[52]。Bommarius等针对来自嗜热脂肪芽孢杆菌Bacillus stearothermophilus的LeuDH中对羧酸盐识别机制的68位点残基进行了定点饱和突变,筛选到一株对甲基异丁基酮具有活性的突变体K68M,但其活性较低[52]。因此对图7B中所示的来自B.sphaericus的LeuDH晶体结构中其他活性位点进行突变,其目的是筛选到对甲基异丁基酮催化活性提高的突变株。优先考虑的突变位点有K68、D114和V291,以及参与辅因子NADH识别的N261。经过突变筛选到最优突变体为K68S/E114V/N261L/V291C,其对甲基异丁基酮的转化率显著提高至92.5%,ee值为99.8%。

图22 LeuDH定向进化获得AmDH的活性[52]Fig.22 Activity of AmDH evolved from LeuDH[52].
Bommarius等研究了LeuDH的体外的定向进化的过程,旨在进行PheDH对酮还原胺化方面的研究[35]。以LeuDH成功突变案例作为指导,将来自Bacillus badius的PheDH进行突变获得双变体K77M/N276V,其对4-甲基-2-戊酮和氟苯基丙酮的活力较野生型酶有了提高[35]。在两个位点的基础上又继续突变,获得了新的变体K77S/N276L,其kcat值较双突变体K77M/N276V又提高了15倍。新突变体K77S/N276L (F-AmDH) 增加了对苯氧基-2-丙酮、2-己酮和3-甲基-2-丁酮的还原胺化活性。F-AmDH是NADH依赖型氧化还原酶,在催化过程中使用葡萄糖和葡萄糖脱氢酶辅酶再生系统,转化氟苯基丙酮得到相应的胺产物,转化率为93.8% (产物分离产率为73.9%),ee>99.8%(图23)。
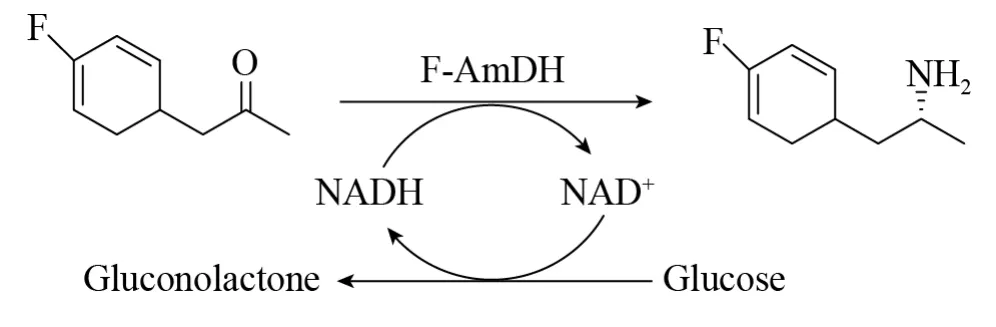
图23 F-AmDH还原胺化氟苯基丙酮,同时使用辅酶循环系统实现辅酶循环[34]Fig.23 Reductive amination of p-fluorophenylacetone by F-AmDH,with cofactor recycling[34] .
通过将F-AmDH的N-端部分 (1–149位点)与LeuDH突变体的140–166的位点组合产生的新嵌合胺脱氢酶cFL1-AmDH,其对苯乙酮和金刚烷基甲基酮具有催化活性,并且具有严格的立体选择性,只形成 (R)-胺产物[53]。在进一步的改造中,嵌合体中的两个相邻的天冬酰胺残基N270和N271突变为亮氨酸,以试图影响其胺化活性,结果表明新突变体催化氟苯基丙酮的kcat值有了提高。F-AmDH对一些底物催化活性较低,可能因为底物在水溶液中的溶解度较差的原因。因此制备了庚烷与水=1︰4的双相反应系统,在这样的催化体系中,F-AmDH催化氟苯基丙酮还原胺化的体积生产率提高为原来的2倍[54]。
随后Li等针对酮的不对称还原胺化,对来自Rhodococcussp.的PheDH进行改造[55]。将其结构中与底物羧酸盐基团相互作用的氨基酸K66、S149和N262进行三重突变,获得了一个K66Q/S149G/N262C三突变体。该突变酶能够催化苯丙酮12和4-苯基-2-丁酮14生成 (R)-苯丙胺13和(R)-1-甲基-3-苯基丙胺15,ee值均>98% (图24)。S149G位点的突变除了改变与底物羧酸盐基团的相互作用之外,还改变了与底物结合口袋入口的大小。来自Rhodococcussp.的胺脱氢酶S149G突变体与葡萄糖/葡萄糖脱氢酶NADH辅酶循环系统相互作用,15 mmol/L的14在60 h后转化为(R)-1-甲基-3-苯基丙胺15,转化率为95.2%。进一步,Li等将AmDH与GDH共同固定在磁性纳米颗粒 (MNPs) 上[57],为14的不对称还原胺化提供了一个合适的反应体系,NADH再循环的总转换数达到2 940。Mutti等验证了来自Rhodococcussp.M4的胺脱氢酶还原胺化活性,结果表明除了底物12和14之外,该酶还可以有效转化邻甲氧基苯基丙酮衍生物,脂族酮 (2-辛酮,转化率99%) 和“大体积”酮类 (如1-苯基-丁-2-酮,转化率>99%;1-苯基戊-2-酮,转化率71%;1-苯基戊-3-酮,转化率83%)[56]。此外,来自Rhodococcussp.的AmDH与甲酸/甲酸脱氢酶辅酶循环系统相互作用,可以将208 mg (对甲氧基苯基)丙酮16转化为 (R)-胺17,产物分离产率为82% (图24)。

图24 由来自Rhodococcus sp.M4的AmDH针对不同酮化合物的还原胺化反应方程式[55-56]Fig.24 Reductive amination of ketones by AmDH from Rhodococcus sp.M4[55-56].
AmDHs的发现促使人们寻找可能具有该活性的天然酶。在排除了作用于胺α-或β-羧酸酯基团的脱氢酶同源物的搜索,Vergne-Vaxelaire等使用了L-erythro-3,5-二氨基己酸脱氢酶(3,5-DAHDH) 的序列作为鉴定AmDHs此类酶的模板[58]。在169个同源物中,选取20个克隆并表达,其中有3个显示出对4-氧代戊酸和5-氧代己酸具有还原胺化活性。来自移动石袍菌Petrotoga mobilis的一种AmDH和甲酸/甲酸脱氢酶辅酶循环系统相互作用,可以将4-氧代戊酸转化为(S)-4-氨基戊酸酸,产物分离产率为88%,ee>99.5% (图25)。目前还没有找到AmDH这种酶的紧密同源物的结构,这些天然存在的AmDH为胺生产提供了新的酶目标。
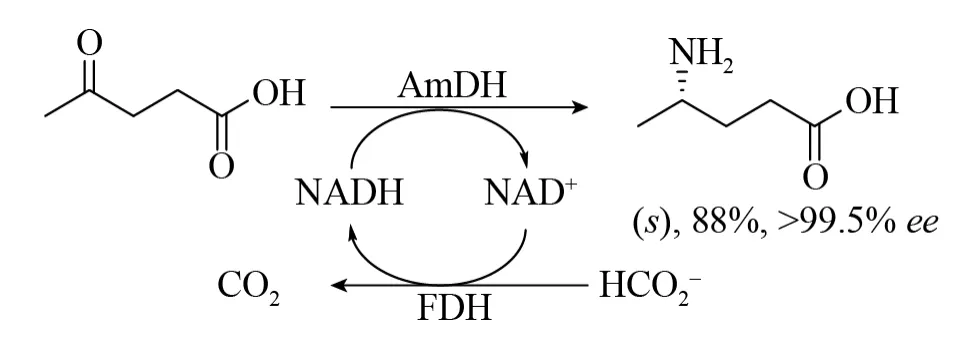
图25 通过来自Petrotoga mobilis的胺脱氢酶还原胺化4-氧代戊酸[58]Fig.25 Reductive amination of 4-oxopentanoic acid by naturally occurring amine dehydrogenase from Petrotoga mobilis[58].
华东理工大学许建和、郑高伟课题组在胺脱氢酶创制方面作出了许多出色的工作,他们以自主研发的氨基酸脱氢酶为模板,开发出了3个新的胺脱氢酶并针对其底物谱较窄的问题,采用计算机辅助的蛋白质工程技术,构建了突变体LfAmDH(A113G/T134G),实现了对活性口袋的“容积拓展”,从而将该酶所催化的底物范围由最长6个碳链的脂肪酮拓展至长达10–12个碳链的脂肪酮,显著地拓宽了该酶催化的底物范围[59]。最近,许建和课题组与曼彻斯特大学Turner教授课题组合作,对来源于纺锤形赖氨酸芽孢杆菌Lysinibacillus fusiformis的氨基酸脱氢酶进行了分子改造,开发出了一系列能够转化-羟基酮不对称还原胺化的胺脱氢酶 (图26),其中的最优突变体可以不对称还原胺化不同链长-羟基酮,合成相应的(S)-邻位氨基醇 (99%的转化率且ee>99%)。该工作将胺脱氢酶的底物谱由酮类化合物拓展到羟基酮类化合物,为手性药物中间体邻位氨基醇的合成提供了新型的生物催化剂[60]。

图26 胺脱氢酶催化的α-羟基酮不对称还原胺化合成手性邻位氨基醇[60]Fig.26 Asymmetric reductive amination of α-hydroxy ketones catalyzed by amine dehydrogenase for the synthesis of chiral vicinal amino alcohols[60].
针对D-氨基酸脱氢酶底物谱狭窄的缺点,许多科学家在拓宽其底物谱上作出了贡献。中国科学院天津工业生物技术研究所朱敦明、吴洽庆课题组对来自Symbiobacterium thermophilum的meso-DAPDH催化口袋进行改造,提高催化口袋的大小容纳更大体积的2-酮酸。选择Phe146、Thr171、Arg181和His227四个点进行突变,获得的StDAPDH突变体H227V催化苯丙酮酸活力达到2.39 U/mg,较原始菌株提高了35.1倍[61]。在此基础上,进行半理性设计及饱和突变,获得一株突变体W121L/H227I,不仅对大位阻2-酮酸底物展现出优良的活性,成功合成了对应的D-氨基酸 (D-苯甘氨酸、D-色氨酸、D-叔亮氨酸),而且提高了对其他结构2-酮酸的比活力。通过将底物与野生型和突变体进行分子对接,发现这些有益突变重塑了底物结合口袋,不仅可以容纳这些大位阻底物,同时拉近了底物与辅酶之间的催化距离,使其获得对大位阻底物的催化活性[62]。另一个有意思的例子是Vedha等对来源于谷氨酸棒杆菌Corynebacterium glutamicum的meso-DAPDH进行三轮定向进化,获得的突变体BC621打破了该酶只能氧化脱氨的限制。该突变体可以催化多种酮酸生成相应的D-氨基酸,其中催化生成D-2-氨基己酸酯、D-2-氨基庚酸、D-2-氨基辛酸酯、D-环己基丙氨酸,酶活力较原始菌株分别提高257、538、975、625倍,ee均大于95%[63]。
日本科学家Akita等成功通过在活性位点附近引入5个突变获得一株热稳定性的突变体。该突变体在65 ℃下活力几乎没有任何损失,具有巨大的工业应用潜力[64]。Hayashi和其合作者通过定点突变技术将来源于嗜热球形脲芽胞杆菌Ureibacillus thermosphaericus的meso-DAPDHs的底物谱进行了拓宽[65]。通过将94位的天冬氨酸替换为丙氨酸 (D94A),该酶的底物谱得到了极大的提高,其对D-亮氨酸、D-正亮氨酸、D-甲硫氨酸、D-异亮氨酸和D-色氨酸的活力提高了6–25倍,其中对苯丙酮酸的活力最高 (16.1 µmol/(min·mg))。通过晶体结构解析,发现D94A减少了底物苯基与侧链天冬氨酸的空间碰撞,通过结构比对发现底物口袋的扩大和口袋疏水性的增强是导致突变株对大位阻疏水性D-氨基酸活性增强的主要原因。一年后,Akita等将来源于U.thermosphaericus的meso-MADPHs进行了组合突变 (Q154L、D158G、T173I、R199M和H249N),创造出了高热稳定性的meso-MADPHs[66]。进一步,通过引入单突变D94A,该高热稳定性的D-氨基酸脱氢酶对大位阻侧链D-氨基酸的活力明显提高。
针对非天然氨基酸前体酮作为底物的氨基酸脱氢酶定向进化,浙江大学杨立荣团队挖掘到来源于恶臭假单胞菌Pseudomonas putida的氨基酸脱氢酶,将其应用到L-草铵膦的合成,采用蛋白质工程技术,获得突变体I170M,其活力较野生型有2.1倍的提高[67]。通过后期的继续工作研究,构建突变体A167G和V378A,活力较之前有了很大提升[68]。
同时,笔者所在课题组采用计算机辅助的蛋白质工程技术,获得一系列谷氨酸脱氢酶超级突变体“草铵膦脱氢酶”,其能高效地转化2-羰基-4-(羟基甲基氧膦基)-丁酸 (PPO) 生成L-草铵膦。与葡萄糖脱氢酶辅酶循环系统相互作用,500 mmol/L的PPO可以在30 min内转化完全 (图27)[69],并通过将“草铵膦脱氢酶”与D-氨基酸氧化酶偶联采用一锅法制备L-草铵膦,以外消旋D,L-草铵膦为底物,D-草铵膦经过D-氨基酸氧化酶催化生成PPO,PPO经过“草铵膦脱氢酶”催化生成L-草铵膦,实现L-草铵膦的高效合成,为工业化生产奠定了基础[70]。
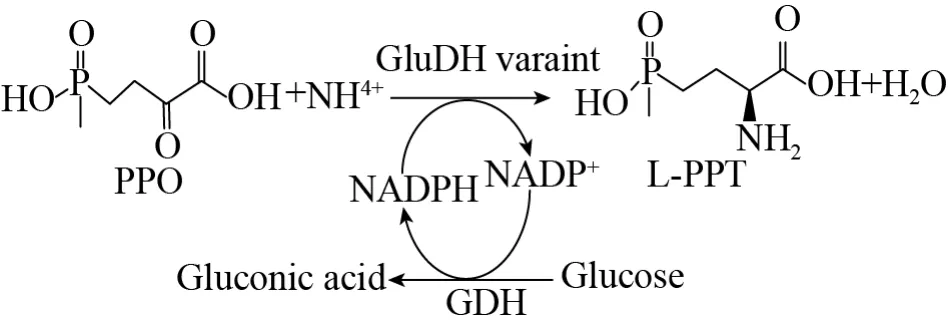
图27 谷氨酸脱氢酶 (GluDH) 不对称还原胺化PPO生成L-PPT,葡萄糖作为第二底物供给葡萄糖脱氢酶辅酶循环系统生成NADPHFig.27 Asymmetric reductive amination of PPO catalyzed by glutamate dehydrogenase mutant for the synthesis of L-PPT and combined with glucose dehydrogenase,glucose as a secondary substrate for the regeneration of NADPH.
3.3 冠瘿碱脱氢酶 (OpDH) 的定向进化与应用
OpDH催化的酮酸不对称胺化反应中,以有机胺作为氨基供体的转化效率大于无机铵,因此Codexis公司的研究人员希望能应用这些酶催化无羧基的酮和有机胺直接的反应[71]。通过以Arthrobactersp.strain 1C菌株来源的CENDH作为出发序列,通过定向进化,他们获得了一株不仅能够催化丙酮酸和L-缬氨酸生成相应的(2S)-2-(1-(羧乙基)氨基)-戊酸,同样能够催化1-丁胺生成相应的2-(丁氨)-丙酸的突变体 (图28)。后续的定向进化又获得了更多的突变体,它们能够利用多种胺来催化环己酮、2-戊酮、2-四氢萘酮的衍生物生成相应的仲胺产物,ee值均大于99.5% (图28)。其中一个七重组合突变体首次表现出了对催化环己酮和丁胺生成相应的仲胺的活性。通过解析CENDH的晶体结构 (1BG6和3C7D),发现这些突变位点都位于两个结构域之间的裂隙内 (图29)。

图28 由定向进化产生的CENDH突变体催化的选择性还原胺化反应[71]Fig.28 Selected reductive aminations catalyzed by CENDH variants created using directed evolution[71].
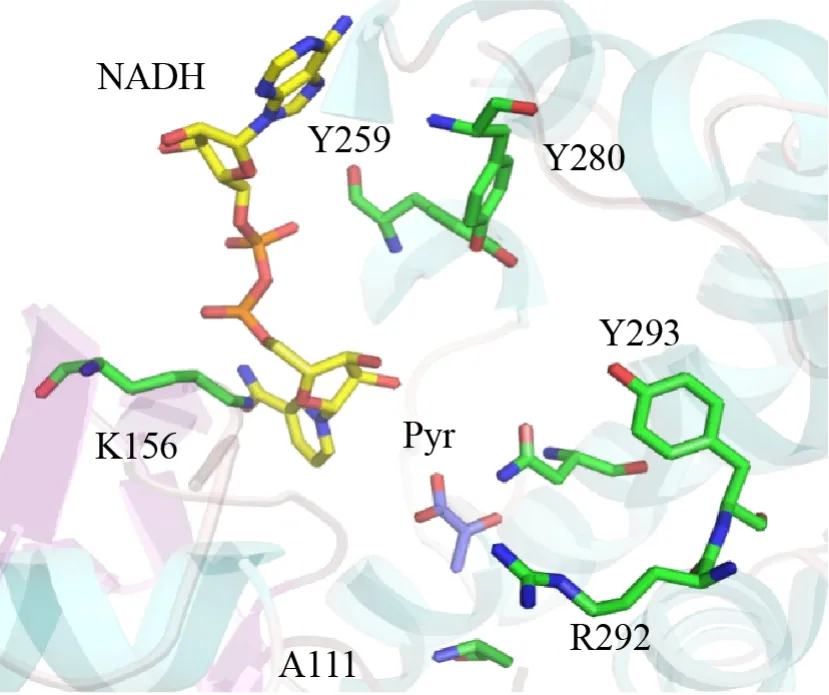
图29 CENDH的活性中心示意图 (含辅酶NADH和丙酮酸)[71]Fig.29 The active site structure of CENDH with NADH and pyruvate[71].
根据以上的这些研究可以清楚地发现在以有机胺为胺供体的情况下,OpDH具有催化酮还原胺化为手性胺的潜力,但是有关OpDH催化机制以及活性位点残基与底物之间的相互作用等方面还不清楚,这些信息将有助于进一步的蛋白质工程研究。
3.4 还原性酮胺化酶 (RedAm) 的定向进化与应用
还原性酮胺化酶可以催化等摩尔的酮与伯胺的不对称还原胺化得到仲胺产物,在合成手性胺方面具有巨大潜力。英国曼彻斯特大学Turner课题组针对来源于曲霉菌A.terreus和A.dermatitidis的还原性酮胺化酶AtRedAm和AdRedAm进行定向进化,获得突变体AtRedAm-Y222A,以烯丙胺作为胺供体,对18、20、21 (图30) 三种不同酮类进行生物还原胺化,其转化率相对野生型酶分别提高了11%、49%和5%。优势突变体I123A使用环己酮19作为酮底物,以A、B、D、E、F、G为胺供体,发现其比酶活较野生型酶提高了2.7–6.2倍;以C、H为胺供体,由野生型无活性变为有活性,比酶活达到8.5 mU/mg和17.2 mU/mg。AdRedAm突变体H215A使用烯丙胺作为胺供体,对18、19、21进行还原胺化,其转化率分别提高了20%、2%和16%[27]。

图30 还原性酮胺化酶涉及到的酮和胺底物[27]Fig.30 Ketone and amine substrates catalyzed by RedAm[27].
4 总结
利用NAD(P)H依赖型氧化还原酶不对称还原胺化已成为制备光学纯手性胺的重要方法,相关的研究引起了国内外学者的极大兴趣,也取得了显著的进展。从现有氨基酸脱氢酶的蛋白质工程改造到天然胺脱氢酶的发掘,再到亚胺还原酶及还原性胺化酶的补充,已经有许多手段和方法进行挖掘、鉴定、改造获得具有还原胺化活性的NAD(P)H依赖型氧化还原酶。通过对这些酶胺化反应机理的解析,可以成功改造这些酶 (氨基酸脱氢酶、胺脱氢酶、冠瘿碱脱氢酶、亚胺还原酶和还原性酮胺化酶) 的部分酶学性质 (表1)。但总体来说这类酶在工业应用上成功的案例还相对较少。未来,针对NAD(P)H依赖型氧化还原酶实际应用的工业环境,突破它们底物谱较窄、活性较低、高浓度底物转化率偏低等瓶颈限制,是实现其在工业上广泛应用的关键所在。
