基于Biolog-Eco法对鄱阳湖不同湿地类型下土壤微生物功能多样性
2020-04-10程家劲陈圆圆刘亚军
吴 兰,程家劲,贺 勇,陈圆圆,刘 颖,魏 鑫,刘亚军,
(南昌大学a.生命科学学院;b.鄱阳湖环境与资源利用教育部重点实验室,江西 南昌 330031)
湿地是介于水陆生态系统之间的过渡带,因其独特的水文特征,使其成为全球碳密度最高的区域[1]。水文通过影响湿地水淹深度、持续时间、干湿交替频次,控制着湿地生物地球化学循环过程[2]。受水情长期驱动的影响,湖泊湿地形成了复杂多样的湿地景观类型[3]。近年来,由于极端天气(干旱、洪水等)的频繁发生,加速了内陆湖泊湿地的退化,主要表现为湿地水位下降、景观格局变化剧烈等生态问题[4-5]。湿地景观类型的变化对湿地生态系统的能量流动、物质循环以及物种迁移等都会产生很大影响[6]。目前围绕不同湿地类型的研究大多集中在湿地植物、野生动物和土壤性质上[7-8]。土壤微生物作为湿地生态系统重要的组成部分,参与了湿地土壤中的物质循环和能量流动,深入研究湿地土壤微生物群落功能多样性,对评价湿地土壤质量和生态系统功能具有重要意义[9]。而当前有关不同湿地类型下土壤微生物功能活性的研究却鲜见报道。
当前,对于湿地土壤微生物的研究大多集中在物种组成和群落结构上,较少直接关注微生物的功能活性,更多的是通过基因序列对微生物群落的功能代谢进行推测[10-11]。而考虑到微生物群落表现出的功能可塑性和功能冗余现象,使得微生物群落结构和组成变化很难直接反映在功能上[12]。因此研究不同湿地类型下土壤微生物的代谢特征,是揭示景观格局剧烈变化下湿地生态功能转变的重要基础。Biolog-Eco微平板法是目前研究微生物代谢多样性的一种有效方法,通过分析微生物对31种碳源的利用程度来反映其功能多样性,能够表征微生物群落的整体活动,而不需要分析单个微生物,这种简单的高分辨率检测方法已广泛应用于微生物生态学和环境微生物检测中[13]。
鄱阳湖作为一个典型的季节性通江浅水湖,湿地类型多样、景观变化剧烈[14]。基于此,在鄱阳湖全湖范围内选择长期水情驱动形成的4种湿地类型为研究对象,采集表层土壤,分析土壤理化性质,同时采用Biolog-Eco微平板法对微生物碳代谢活性特征进行分析,以揭示不同湿地景观下土壤的生态功能特征变化,以期为湿地生态保护提供理论依据。
1 材料与方法
1.1 研究地概况及样地介绍
鄱阳湖是我国第一大淡水湖泊,地处江西省北部、长江中下游南岸,承接五河之水,汇通长江,是典型的洪泛湖泊。所在区域属亚热带湿润季风型气候,年均气温17.6 ℃,年降雨量1 450~1 550 mm。鄱阳湖水位呈明显的周期性变化(平均年内水位变幅为11.01 m),每年10月进入枯水期,随着水位的涨落,鄱阳湖湿地呈现出草洲湿地、河流湿地、泥滩湿地、湖泊湿地等多种湿地类型,形成了以湖泊湿地为核心的多类型复合的湿地生态系统类型[15]。本实验选取草洲、泥滩、河口、湖泊作为典型的研究样地,其土壤质地及淹水状况见表1。

表1 不同湿地类型土壤质地及淹水情况
1.2 样品采集
本研究于鄱阳湖枯水期进行样品采集(具体采样时间及样地坐标信息如表2所示),此时湖泊湿地土壤表现出不同的覆盖类型。选择草洲(为更具有代表性选择处于不同高程上的3种植被群落:狗牙根、南狄和苔草,采样时植被覆盖)、河口(采样时为入湖河水覆盖)、泥滩(采样时地表裸露无植被)和湖泊(采样时为湖泊水体覆盖)4种湿地类型土壤进行研究,每种类型各设置3个大样地(图1),在每个样地内部设置3个平行样方(1 m×1 m),相邻样方相距200 m。根据五点混合采样法用土钻采集表层土壤样品(0~20 cm),共采集36个土壤样品。样品采集后,剔除残留的草根和石砾,取部分土壤样品自然风干后用于土壤理化性质的测定,其余新鲜土壤装于自封袋中保存在4 ℃冰箱,用于土壤微生物Biolog生态板培养分析。

表2 取样地坐标、时间及淹水状态
1.3 样品分析及方法
参照相关文献[16-17]测定土壤理化性质,其中土壤含水量(SM)采用烘干恒重法(105 ℃,8 h);pH采用水浸提点位法(2.5:1的水土比);土壤有机碳(SOC)采用重铬酸钾外加热法;土壤总氮(TN)和总磷分别采用半微量凯式法和钼锑抗比色法;土壤可溶性有机碳(SDOC)采用TOC仪法;微生物生物量碳(MBC)采用氯仿熏蒸—K2SO4(0.5 mol·L-1)浸提法。
土壤微生物碳源代谢特征的测定采用Biolog-Eco微平板法[18]。具体步骤:取10 g新鲜土壤放入250 mL形瓶中,加入90 mL 0.85% NaCl溶液,振荡30 min,制备成土壤微生物悬浮液,静置片刻吸取上清液并稀释1000倍得到菌悬液。吸取150 μL的菌悬液接种在Biolog-Eco微平板中,置于25 ℃恒温培养箱连续培养10 d。在培养过程中,每隔24 h通过Biolog自动读数装置读取590 nm处的吸光值(OD)[19]。
1.4 数据分析
选用120 h OD值分析土壤微生物对不同碳源的利用以及代谢功能多样性的差异。首先将每孔OD值减去培养0 h的初始OD值,用以排除本底干扰[20],然后计算平均每孔颜色变化率(Average well color development,AWCD)。同时参照文献[21]选用Shannon指数、Simpson指数和Evenness指数反应微生物群落的代谢功能多样性,计算方法如下:
AWCD=Σ(Ci-R)/31;
Shannon指数H′=1-∑(PilnPi);
Evenness指数J=H′/lnS;
Simpson指数D=1-∑Pi2
式中:Ci表示每种碳源孔的OD值;R表示对照孔的OD值;Pi表示第i个非对照孔中的吸光值与所有非对照孔吸光值总和的比值;S为被利用的碳源总数。
此外,利用SPSS 20.0对数据进行差异性分析和Pearson相关性分析。主成分分析(PCA)通过Canoco 5.0软件完成。
2 结果及分析
2.1 不同景观类型下的土壤理化性质
不同湿地土壤类型的理化性质和微生物生物量表现出不同的差异特征。由表3可见,土壤含水量(SM)由湖泊、河道、泥滩、草洲依次递减,差异显著(P<0.05)。研究发现,所有的湿地土壤均呈现出弱酸性,范围在5.03~6.86之间,其中草洲土壤酸性最强,pH值为5.03,显著低于其它3种湿地土壤(P<0.05)。对于土壤营养元素,草洲与湖泊湿地表现出更高的含量的SOC和TN,显著高于泥滩和河口湿地土壤(P<0.05),其中草洲与湖泊湿地SOC含量分别达到了12.993和14.93 g·kg-1,泥滩和河口湿地SOC分别为6.21和6.7 g·kg-1。而对于土壤TP,各土壤类型间并未表现出显著差异。值得注意的是,对于土壤可溶性有机碳(SDOC)和微生物生物量碳(MBC),草洲湿地土壤显著高于其他3种湿地土壤(P<0.05),分别高达106和462.19 mg·kg-1。

表3 土壤理化参数
2.2 土壤微生物利用碳源的动力学特征及功能多样性分析
AWCD值体现了微生物群落对碳源利用的总体状况。由图2可知,不同湿地类型下土壤微生物AWCD值,培养时间的延长逐渐增大。在培养24 h内变化不明显,均小于0.1,培养24~120 h AWCD值迅速增大,微生物进入指数生长期,底物碳源被大量利用,120 h以后进入稳定期,至240 h时达到平衡。据此,本研究有关微生物碳源利用的分析,均采用120 h时的AWCD值计算获得。对比不同湿地类型土壤的AWCD值发现(表4),草洲土壤微生物群落对底物碳源的总体利用能力最强,其次是河口,最后是湖泊和泥滩土壤(P<0.05)。
土壤微生物群落功能多样性能够反映群落整体的动态变化,代表了微生物群落数量分布和物种组成信息。多样性指数分析表明(表4),4种湿地土壤微生物群落功能多样性存在显著差异,其中草洲湿地土壤微生物3类多样性指数显著高于其他湿地土壤(P<0.05)。同时研究还发现,对于Shannon和Simpson指数而言,泥滩表现出更低功能多样性。

表4 土壤微生物群落功能多样性指数
2.3 土壤微生物对碳源的利用分析
Biolog-Eco微平板上31种碳源,分属于6大类碳源,分别为聚合物类、碳水化合物类、酚酸类、羧酸类、氨基酸类和胺类。结果如图(3)所示,鄱阳湖不同湿地土壤微生物对不同碳源相对利用率存在显著差异。总体而言,4种湿地类型土壤均表现出对羧酸类、氨基酸类和胺类碳源利用的偏好性,占总碳原利用率的14.73%~26.69%;而对聚合物类和碳水化合物类表现较低的利用能力,占总碳原利用率的9.33%~14.64%。具体来看,与其他3种湿地土壤相比,草洲湿地土壤对聚合物类、碳水化合物类及酚酸类碳源具有更好的利用偏好性(P<0.05),分别占总碳原利用率的14.65%,12.91%和20.08%;泥滩土壤微生物更倾向于利用氨基酸类碳源,占其总碳源利用率的26.69%;河口湿地土壤微生物倾向于利用酚酸类和胺类碳源,分别占其总碳原利用率的18.86%和23.56%;此外,湖泊湿地土壤微生物对羧酸类碳源有更好的利用倾向,占其总碳原利用率的22.14%。
2.4 不同湿地类型土壤微生物碳代谢图谱主成分分析
对31种碳源的AWCD值进行主成分分析(图4)。由图可知,不同湿地土壤微生物的碳源利用能力有着显著的分异情况。4种湿地土壤聚为3簇,其中草洲聚于PC1正半轴,泥滩聚于PC2正半轴,河口与湖泊聚为一簇,位于PC1和PC2的负半轴。初始载荷值反映了主成分分析图上碳源利用的相关系数,载荷值越高表示对主成分贡献越大。对筛选31种碳源中与每一主成分得分系数较高(载荷绝对值>1)的碳源进行分析(表5)。表5可以看出,与第1主成分PC1相关性较高的碳源有7种碳源,分属于6类碳源,其中多聚物类1种、碳水化合物类2种、酚酸类1种、羧酸类2种、氨基酸类1种、胺类1类,表明草洲湿地土壤微生物大量的碳源代谢功能不同于其他湿地土壤。而与PC2相关性较高的碳源仅有4种,分属于3类碳源,其中羧酸类2种、氨基酸类1种、胺类1种。表明泥滩、河口和湖泊湿地土壤碳源利用特征差异较小,主要体现在羧酸类、氨基酸类和胺类上。
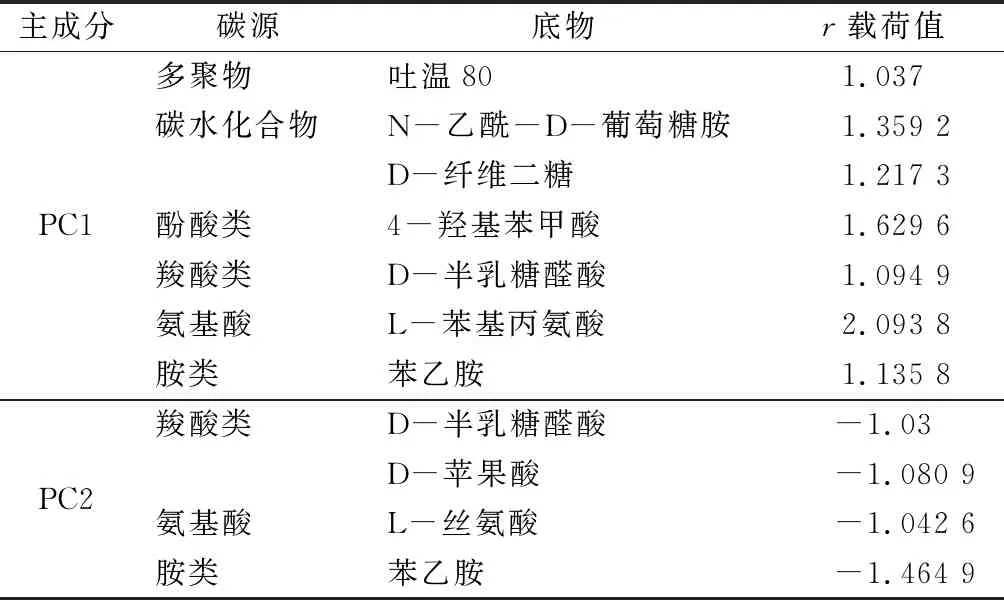
表5 31种碳源的主成分载荷因子
2.5 环境因子对于微生物群落代谢功能影响分析
土壤物化环境决定了微生物的生长状态和组成特征。经相关分析发现(表6),SM、pH与6类碳源的利用能力及3种微生物群落多样性均呈显著负相关(P<0.05)。值得注意的是,SOC、TN、TP作为土壤的主要营养元素,除SOC与碳水化合物呈显著正相关外,与其他5种碳源利用及功能多样性指数均无相关性(P>0.05)。然而对于土壤中最为活跃的碳组分SDOC和MBC而言,除MBC与胺类碳源、辛普森指数无相关性外,与其他碳源利用能力均呈显著正相关(P<0.05)。表明,湿地土壤微生物对碳源的利用很少受土壤营养盐总量的影响,而是受土壤中活性碳组分的影响。

表6 土壤理化性质与微生物群落多样性指数及利用不同碳源的相关性
3 讨论
受水情驱动的影响,鄱阳湖湿地形成了以湖泊湿地为核心的多类型复合的微生物土壤生境,在不同的湿地类型中,土壤物化环境差异巨大[22]。本研究发现,受土壤淹水条件的影响,常年积水的湖泊、河口湿地SM显著高于周期性淹水的泥滩、草洲土壤(表3)。同时,草洲土壤的酸度更强,pH低至5.03,这可能是因为植被根系分泌物的释放和残体降解会生成一系列酸性代谢产物,从而降低了土壤pH[23]。在本研究中,湖泊和草洲湿地土壤中均表现出较高的SOC和TN含量,同时,相较于其他3类湿地土壤,草洲湿地土壤中还表现出更高的SDOC和MBC含量,分别达到了106和462.19 mg·kg-1(表3)。一般研究认为,地上植被持续的向土壤中输入有机质是其维持高含量营养盐的重要原因[24]。而对于湖泊湿地土壤,较高的碳氮含量得益于其独特的土壤环境,湖泊湿地土壤一般处于淹水状态,温度低且变幅小,同时通气性较差,抑制了好氧微生物的生长,致使土壤有机质分解十分缓慢,逐渐形成了富含有机质的湿地土壤,使其成为重要的碳汇[25]。对于同样处于过湿状态的河口和泥滩湿地土壤分别为粉砂质粘土和砂质壤土(表1),较低的碳氮含量很可能是不同的土壤质地导致的,有研究发现土壤中含有较高的砂粒不利于碳的储存[26]。
本研究中,4种湿地类型土壤均表现出对羧酸类、氨基酸类和胺类碳源具有较强的利用能力(占总碳原利用率的14.73%以上),而不善于利用聚合物类和碳水化合物类碳源(占总碳原利用率的14.64%以下)。这可能是由于湿地土壤一般处于湿润或过湿状态,形成的这种特殊的还原环境导致的,这在以往的研究中得到了证实[27]。微生物在不同的湿地土壤生境下,其功能特征出现了显著的分异变化(图4)。本研究中草洲湿地土壤微生物表现出了更高的碳源代谢潜力(AWCD)和功能多样性(均匀度指数、香农指数和辛普森指数)。此外,相对其他湿地土壤微生物而言,草洲对聚合物类、碳水化合物类及酚酸类碳源具有更高的利用偏好性,分别占总碳原利用率的14.65%,12.91%和20.08%。以往的研究发现,地上植被产生的凋落物和根系分泌物在增加土壤有机碳和总氮的含量的同时,会加速土壤的物化循环,使得土壤微生物碳源代谢能力和利用碳源的多样性较高[28]。此外,相对于其他湿地土壤,草洲湿地土壤远离湖心,地势较高,淹水时长最短,因而其土壤环境更加适宜微生物的生长,生物活性更高,更加多样[29]。同样,对于相对干燥的草洲土壤(含水量显著低于其他3种湿地土壤),受水分条件的影响,土壤微生物更倾向于利用碳水化合物和聚合物类碳源,这在先前受控实验的研究中已得到证实[17]。需要注意的是,在本研究中,选择了3种植被覆盖类型代表周滩湿地,虽然这在一定程度上能够更全面的反映草洲湿地土壤微生物的功能代谢特征,但不同湿地植被下土壤物化性质和微生物功能活性仍存在较强的异质性[16],因而,未来对湖泊湿地土壤微生物的研究还应关注不同植被覆盖类型带来的影响。而对泥滩湿地碳代谢特征的研究中发现,土壤微生物更倾向于利用与氮相关的氨基酸类碳源(图3),占总碳源利用率的26.69%,这主要是由于泥滩土壤频繁干湿交替的水环境决定的[30]。有研究发现,对于同样处于干湿交替的河岸带土壤,具有更高的N去除潜力[31]。而对于常年淹水的河口和湖泊湿地,其土壤微生物功能代谢更加类似(图3~4),但仍表现出一定的差异特征,其中河口湿地土壤微生物对酚酸类和胺类碳源利用能力相对较强,分别占其总碳原利用率的18.86%和23.56%,而湖泊湿地土壤微生物对羧酸类碳源具有偏好性,占其总碳原利用率的22.14%。这主要是由于土壤中有机质的来源不同造成的,酚类物质和胺类物质主要来源于人类的生产生活,在河流中有大量富集,这可能是造成河口湿地土壤较高利用能力的原因[32-33]。而对于湖泊湿地土壤,长期淹水形成的还原环境,厌氧微生物占据主导地位,其分解代谢途径以发酵、乙酰化占优势,从而表现出对羧酸类碳源的偏好性[34]。本研究发现,不同的湿地土壤微生物存在着显著的碳代谢功能特征差异,这对维持整个湖泊湿地生态系统生态功能的发挥极为重要。
土壤的物化环境是影响微生物生长代谢的关键因素。与以往研究类似[27,35],土壤含水量和pH均是影响湿地土壤微生物碳源代谢功能的关键性因素(表6)。其中土壤含水量与土壤碳代谢活性及功能多样性呈负相关关系(P<0.05),表明随着湿地土壤含水量的降低,土壤活性逐渐增强,这不利于湖泊湿地土壤对碳的汇集。大量的研究发现,湖泊湿地土壤有机质的积累得益于其过湿的水环境和还原条件,而干旱条件会增加土壤碳的释放,与本研究结论一致[36]。一般认为,微生物的生长代谢活性主要受土壤营养状况的影响[37]。在本研究中,不同湿地土壤碳源代谢特征并不是受SOC、TN和TP的直接影响(P>0.05),而是与土壤活性碳组分(SDOC和MBC)有很好的耦合关系。这可能是因为湿地土壤微生物更多的受土壤质地和水环境的影响[38]。例如,湖泊湿地虽然拥有较高的营养(SOC和TN)物质,但受长期淹水环境的影响,微生物代谢活性受到抑制。而SDOC和MBC作为土壤碳储存库中最活跃的组分,其含量大小反映土壤同化和矿化能力,是土壤微生物活性大小的标志[39],有研究基于微生物群落代谢预测发现,湿地土壤微生物的功能代谢特征主要受DOC的影响,与本研究结果类似[]。综上所述,本研究发现,湿地土壤微生物群落代谢活性受土壤含水量和pH的负向影响,而与SDOC和MBC有很好地相关性。
4 结论
鄱阳湖不同湿地类型下土壤微生物的总体碳代谢能力不同,草洲湿地土壤微生物具有最高的碳代谢能力,其次是河口,最后是湖泊和泥滩湿地。此外,相较于其他3种湿地类型,草洲湿地土壤微生物具有更高的功能多样性。
4种湿地类型土壤均表现出对羧酸类、氨基酸类和胺类碳源具有较强的利用能力(占总碳源利用率的14.73%~26.69%)。此外,不同湿地类型下土壤微生物对不同碳源具有一定的利用偏好差异,相对其他湿地土壤而言,草洲表现出对聚合物类、碳水化合物类及酚酸类碳源具有偏好性,分别占总碳原利用率的14.65%,12.91%和20.08%;泥滩更倾向于利用氨基酸类碳源,占其总碳源利用率的26.69%;河口倾向于利用酚酸类和胺类碳源,分别占其总碳源利用率的18.86%和23.56%;湖泊倾向于利用羧酸类碳源,占总碳原利用率的22.14%。
相关性分析发现,湿地土壤微生物的碳源利用能力及功能多样性受土壤含水量和pH的负向影响,受SDOC和MBC的正向影响。
